
益生菌延长秀丽隐杆线虫寿命分子机制研究进展
2023-03-23 06:22张俊桃赵艳梅孙天松
中国食品学报 2023年2期
张俊桃,赵艳梅,孙天松*
(1 内蒙古农业大学 乳品生物技术与工程教育部重点实验室 农业农村部奶制品加工重点实验室 内蒙古乳品生物技术与工程重点实验室 呼和浩特 010018 2 中国科学院生物物理研究所 中国科学院生物大分子研究中心 RNA 生物学重点实验室 北京 100101)
衰老是一种具有时间依赖性的生物复杂过程。衰老的标志是组织、细胞以及器官的功能衰退和死亡风险的增加,是机体随年龄增加发生的退行性变化,主要受饮食、环境和遗传因素等影响[1]。已有研究发现,神经退行性疾病、糖尿病、心血管疾病乃至癌症等疾病的发生率和死亡率均同患者的年龄呈正相关[2]。
益生菌(Probiotics)最早源自希腊语“for life”,意为“对生命有益”[3]。Metchnikoff[4]提出“益生菌”一词,并推测保加利亚人的长寿可能与其摄入乳杆菌发酵乳制品密切相关。随着时间的推移,益生菌有了较明确的定义:活的微生物,当摄入足够数量时,对宿主健康产生有益作用[5]。目前发现的益生菌促进健康作用,包括改善肠道微生态平衡[6-7],抵抗有害菌[8-9],免疫调节[10]和延缓宿主衰老[11-12]等。
秀丽隐杆线虫(简称线虫)是一种体长约为1 mm 的无脊椎蠕虫,实验室培养多以大肠杆菌(Escherichia coliOP50)为食。由于其存活周期短(2~3 周),遗传筛选可获得大量纯合突变体和易于培养且不存在伦理问题等特点,成为生物学研究的理想模型[13]。
1 益生菌延缓线虫衰老的研究现状
不同细菌摄食可能诱导特定的宿主反应,进而在宿主寿命调控中发挥重要作用[11]。一些致病菌会缩短线虫的存活时间[14],而益生菌会延长线虫寿命并延缓其衰老[15]。这些效应与线虫的宿主防御能力和抵抗外界环境压力密切相关[16]。表1中总结了以线虫为模式生物进行的益生菌延缓宿主衰老的信号通路研究,如营养感知信号通路、p38 MAPK 信号通路、线粒体信号通路和应激反应,为探讨益生菌延缓宿主衰老的作用机理提供理论参考。
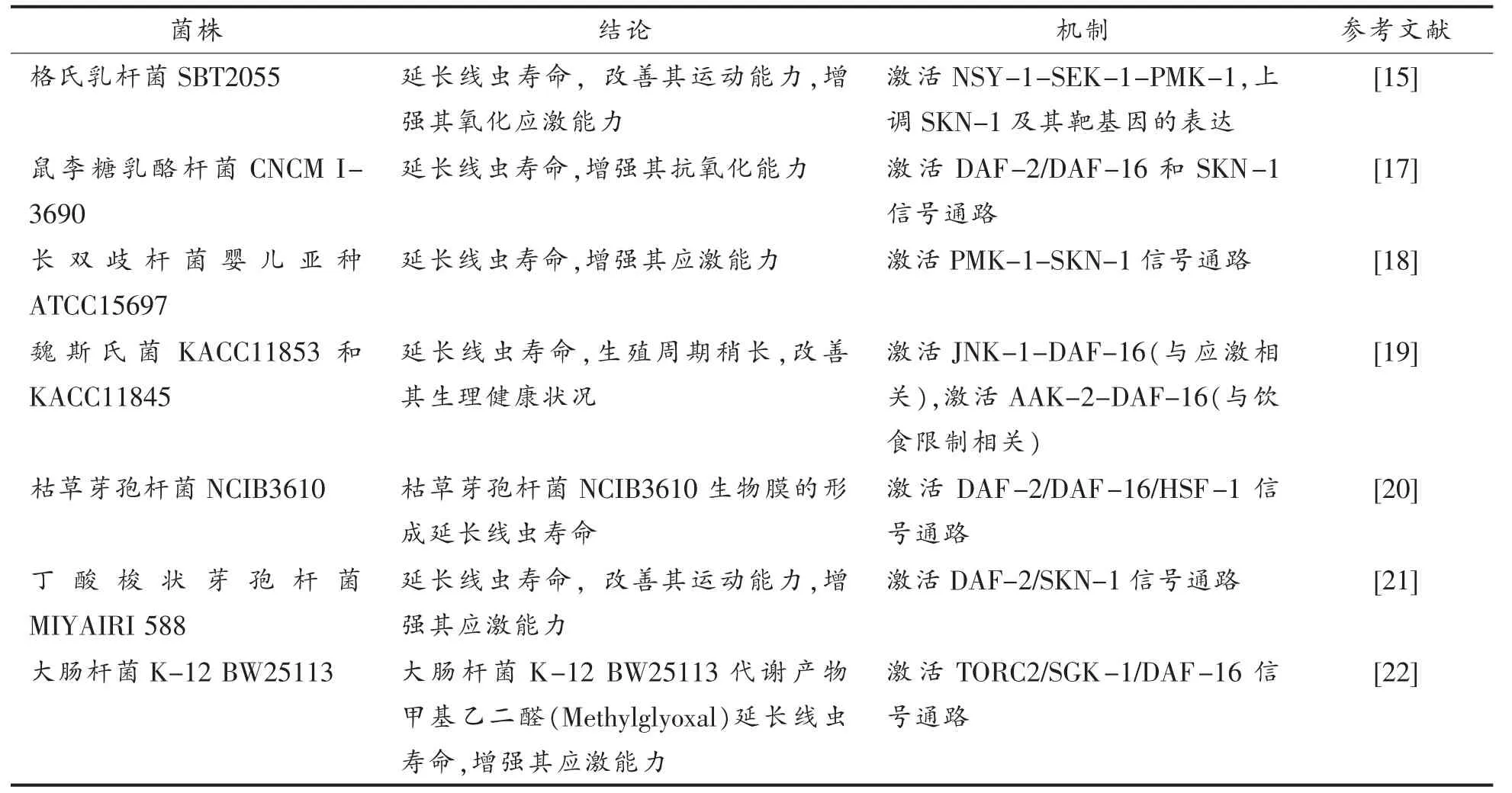
表1 益生菌延长线虫寿命的分子机制Table 1 Molecular mechanisms of probiotics on the longevity of C.elegans
2 益生菌延长线虫寿命的分子机制
2.1 益生菌与营养感知信号通路的调控研究
线虫作为生物个体,其衰老受饮食、环境和遗传因素等影响,并且也会受生理状况的影响,包括生长和发育。补充益生菌对宿主有多种有益作用,可以保护宿主免受有害菌或病毒的侵害,增强免疫力,进而促进宿主健康。如图1所示,益生菌作为膳食补充剂,通过一系列信号分子,最终激活或调节特定靶基因的转录,进而调控线虫寿命。
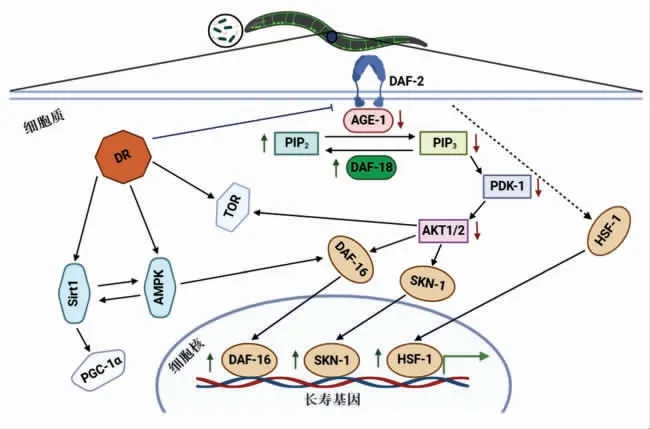
图1 益生菌影响营养感知信号通路示意图Fig.1 Schematic diagram of probiotic influencing nutrient sensing signaling pathways
2.1.1 益生菌作用于胰岛素信号通路 胰岛素(IIS)信号通路包含多种进化保守的衰老调节途径,是目前研究最为完整的应激反应和营养感知通路之一。
IIS 根据周围环境条件传递信号[23]。营养充足时,DAF-2 激活AGE-1/PI3K,同时促进磷脂酰肌醇4,5-二磷酸肌醇【PI(4,5)P2】生成磷脂酰肌醇3,4,5-三磷酸肌醇【PI(3,4,5)P3】,激活PDK-1和AKT-1/2[24-25],且激活的磷酸水解酶DAF-18/PTEN 通过水解PI(3,4,5)P3 生成PI(4,5)P2,进而影响下游信号通路[26]。此外,激活的AKT-1/2 磷酸化转录因子DAF-16,使得DAF-16 与14-3-3蛋白结合并停留在细胞质中,阻止其进入细胞核发挥转录因子功能[27-28]。反之,饥饿时,类胰岛素不足,DAF-2 受体激活受到抑制,AGE-1 激酶活性下调,使得PI(3,4,5)P3/PI(4,5)P2 比例随之下降,同时抑制PDK-1 和AKT-1/2 活性,此时DAF-16 进入细胞核发挥其转录因子功能,进而延缓宿主衰老。Grompone 等[17]研究发现,鼠李糖乳酪杆菌CNCM I-3690 通过激活DAF-2/DAF-16/SKN-1 信号通路延长线虫寿命。
DAF-16 是IIS 下游的关键转录因子[29],并且DAF-16 可由Jun-N 末端激酶[30]和腺苷酸活化蛋白激酶(AMPK/AAK-2)[31]磷酸化激活。Schifano 等[32]研究发现,发酵粘液乳杆菌MBC2 通过激活DAF-16 信号通路延长线虫寿命。此外,益生菌代谢产物也具有延缓衰老的作用,例如大肠杆菌K-12 BW25113 代谢产物甲基乙二醛(Methylglyoxal)通过调节DAF-16 信号通路延长线虫寿命[22]。
SKN-1 是一种氧化应激胁迫NRF 转录因子,与DAF-16 类似,IIS 蛋白激酶磷酸化SKN-1 使其停留在细胞质中,参与应激胁迫和解毒相关基因的表达[33]。此外,SKN-1 还可以通过产生蛋白酶体从而促进蛋白质稳态,进而延长宿主寿命[34]。Kato 等[21]研究表明,丁酸梭状芽孢杆菌MIYAIRI 588 通过调控IIS 信号通路激活SKN-1/Nrf2 转录因子进而延长线虫寿命。Komura 等[18]研究发现,长双歧杆菌婴儿亚种ATCC15697 通过调控IIS 和p38 MAPK 信号通路进而激活SKN-1/Nrf2 转录因子延长线虫寿命。
HSF-1 是另一种作用于IIS 下游的转录因子,其降低蛋白毒性和促进长寿的作用与保守的IIS 通路密切相关[35]。此外,HSF-1 与包含热休克因子(HSEs)的DNA 特定区域结合[36],诱导编码分子伴侣基因hsp-70 和hsp-16,使其过表达进而延长宿主寿命[37-38]。Donato 等[20]研究发现,枯草芽孢杆菌NCIB3610 形成的生物膜通过激活DAF-2/DAF-16/HSF-1 信号通路延长线虫寿命。
2.1.2 益生菌作用于雷帕霉素靶蛋白信号通路 雷帕霉素靶蛋白(TOR)首次发现于酿酒酵母中,随后在其它物种中检测到[39],是一种跨物种的保守的丝/苏氨酸蛋白激酶,包含TOR 复合物1(TORC1)和TOR 复合物2(TORC2),调节合成多个代谢途径[40-41]。
当雷帕霉素存在时,TORC1 可以抑制TOR 激酶活性,进而调控蛋白质稳态、自噬和能量代谢等生理过程[42]。此外,TORC1 调控寿命的关键因子为RSKS-1(核糖体蛋白S6 激酶S6K)和RAGA-1(Ras-related GTPase)[43-44]。Han 等[12]研究发现,大肠杆菌K-12 BW25113 ΔpsuK通过调节TORC1/RAGA-1/RSKS-1 信号通路延长线虫寿命。
TORC2 亦称作PDK2,可调控血清和糖皮质激素诱导的蛋白激酶(SGK)的活性[22],同时有助于激活Akt[45]。Shin 等[22]研究发现,大肠杆菌K-12 BW25113 代谢产物甲基乙二醛通过调节TORC2/SGK-1 信号通路延长线虫寿命。
2.1.3 益生菌作用于饮食限制信号通路 饮食限制(DR)是指保证营养充足的同时适当减少食物摄入,广泛应用于酵母、线虫和果蝇等物种的长寿研究[46]。
对线虫进行DR 的方式有很多种,不同的DR条件涉及的调控机制也不一样,因此表型也有差异[47]。目前,研究较多的DR 主要是进食缓慢的“eat”突变体,eat-2 编码维持线虫咽部肌肉功能的配体门控离子通道,其突变后使得咽部蠕动缓慢,进而减少进食[48]。Zhao 等[49]研究发现,唾液联合乳杆菌FDB89 通过DR(eat-2)依赖的方式延长线虫寿命。
腺苷酸活化蛋白激酶(AMPK)信号通路也是高度保守的营养感知信号通路,在调节宿主衰老方面具有重要作用,通常在饥饿时激活[50],通过增强抗应激能力延长线虫寿命[51]。其中,aak-2 是哺乳动物AMPK 的α-催化亚基同源物,且在哺乳动物和线虫中都具有保守性[52]。Lee 等[19]研究发现,魏斯氏菌KACC11853 和KACC11845 延长线虫寿命依赖于饮食限制AMPK(aak-2)信号通路。
Sirtuins 是烟酰胺腺嘌呤二核苷酸(NAD)依赖蛋白去乙酰化酶,通常作为潜在抗衰老因子应用于衰老研究[53]。Sirtuins 起源于对酵母、果蝇和线虫的一系列研究,通过DR 诱导细胞内NAD+水平增加,激活Sirtuins,进而延长宿主寿命[54]。Sugawara 等[55]研究发现,热灭活的长双歧杆菌长亚种BR-108 通过激活SIR-2.1 信号通路延长线虫寿命。
2.2 益生菌通过p38 丝裂原活化蛋白激酶信号通路调控线虫寿命
p38 丝裂原活化蛋白激酶(MAPK)是线虫免疫系统中保守的MAPKs 亚家族信号通路,是通过遗传筛选鉴定的对铜绿假单胞菌致死较敏感的突变体[56]。在人类中,p38 级联反应通常由病原体暴露和炎症性细胞因子激活[57-58]。在线虫中,p38 级联反应由NSY-1 MAPK 激酶、SEK-1 MAPK 激酶和PMK-1 激活[56]。Nakagawa 等[15]研究表明,格氏乳杆菌SBT2055 通过激活p38 MAPK 信号通路和增强氧化应激SKN-1 能力进而调控先天免疫应答延长线虫寿命。此外,Park 等[10]研究发现,发酵粘液乳杆菌JDFM216 通过激活PMK-1 信号通路和核激素受体(NHR)家族延长线虫寿命。
PMK-1 信号通路上游Toll-白细胞介素-1 受体(TIR)结构域效应蛋白TIR-1,其人源同源蛋白SARM 是线虫中唯一的TIR 结构域效应蛋白[59],该蛋白在p38 级联反应上游发挥特异功能[60],且是激活下游PMK-1 信号通路所必需的[61]。Zhao 等[62]研究表明,长双歧杆菌长亚种BB68 通过激活TIR-1-JNK-1-DAF-16 信号通路延长线虫寿命。Kim 等[63]研究发现,嗜酸乳杆菌NCFM 通过调控p38 MAPK(TIR-1 和PMK-1)和β-catenin 信号通路(BAR-1),进而激活线虫防御革兰氏阳性细菌的关键免疫信号通路。
碱性亮氨酸拉链(bZIP)转录因子家族保守的cAMP 反应元件结合蛋白(CREB)/激活转录因子(ATF)ATF-7,位于PMK-1 信号通路下游,是哺乳动物ATF2/ATF7/CREB5 的同源基因,参与p38 调控[64]。Wu 等[65]研究发现,通过饮食限制(DR)激活p38-ATF-7 信号通路,进而调控先天免疫应答延长线虫寿命。
2.3 益生菌作用于线粒体信号通路
随着细胞和机体的衰老,线粒体呼吸链功能随之减弱,释放的电子增加,合成的ATP 减少[66],进而引起线粒体的功能和结构发生改变[67]。抑制线粒体呼吸链功能减弱有助于延缓宿主衰老[68-69],如电子传递链(ETC)基因突变可延长线虫和果蝇寿命[69-70]。
此外,线粒体内蛋白的表达情况也与线粒体功能密切相关。若线粒体的转录或翻译过程受损,则会引起一些未折叠或错误折叠的蛋白聚集,发生线粒体未折叠蛋白反应(UPRmt)。UPRmt是线粒体启动核基因编码的线粒体热休克蛋白和蛋白酶等基因的转录激活过程的应激反应,有助于维持线粒体稳态,减少线粒体内错误折叠的蛋白大量聚集[71]。调控UPRmt的转录因子ATFS-1 具有线粒体核定位和靶向序列[72],当ATFS-1 从线粒体转位到细胞核时,激活UPRmt[73]。此外,具有延长寿命效应的ETC 突变也可以激活UPRmt[74]。Han 等[12]研究发现,大肠杆菌K-12 BW25113 代谢产物荚膜异多糖酸(CA)通过调节宿主线粒体动态变化和UPRmt,进而延长线虫寿命,且纯化的CA 聚合物通过ATFS-1 促进线虫寿命延长。随后,Shin 等[22]研究发现,大肠杆菌K-12 BW25113 Δhns通过激活UPRmt进而延长线虫寿命。
2.4 益生菌作用于压力应激反应
应激反应是指机体或细胞在其所处环境发生改变时抵抗压力的能力,包括热应激、氧化应激、蛋白毒性应激和渗透胁迫应激等[47]。随着机体的衰老其在压力作用下生存能力逐渐下降[75],线虫对热应激、百草枯、渗透胁迫和致病菌的抵抗能力随之减弱[75-77]。然而,具有长寿效应的age-1和daf-2突变体其抗氧化应激和热应激能力明显增强[16,78]。因此,增强机体抗应激能力可能有助于延缓宿主衰老[79]。Martorell 等[80]研究表明,动物双歧杆菌乳亚种CECT8145 通过激活IIS 信号通路增强线虫氧化应激能力进而延缓衰老。此外,Schifano 等[32]研究发现,发酵粘液乳杆菌MBC2 不仅可以减缓老年线虫肌肉功能衰退,并且通过降低线虫体内活性氧(ROS)积累水平及增加GST-4 解毒酶的表达延缓线虫衰老。Donato 等[20]研究发现,枯草芽孢杆菌NCIB3610 通过增强线虫耐热性、渗透压应激、金属胁迫和H2O2氧化应激能力延长线虫寿命。
3 总结与展望
近年来,越来越多的研究发现益生菌具有促进健康的益处。膳食中补充益生菌广泛应用于探究延缓宿主衰老,增强机体健康以及减少年龄相关疾病的发生。目前应用最多的益生菌是乳酸菌和双歧杆菌,因其普遍存在于人和动物的胃肠道中,常作为益生菌研究的首选菌株。利用线虫作为模式生物来探究益生菌延缓宿主衰老相比无菌小鼠、无特定病原菌小鼠和果蝇,具有生命周期短、易获得大量纯合突变体,且可以同时研究多种益生菌间的相互作用等特点。
益生菌的益生特性大多通过益生菌具有维持肠道菌群平衡、提高免疫力、减少炎症、降低胆固醇等方面来表征,因此需要进一步明确益生菌延缓衰老的相关作用机制,进而深入探究益生菌延缓宿主衰老可能作用的信号通路,为益生菌有助于延缓宿主衰老提供理论依据。此外,明确益生菌中延缓衰老的主要成分尤为重要,针对不同益生菌菌株所具有的不同益生特性,有望开发复合益生菌产品以增强其促进健康作用。一些研究发现,益生菌作为膳食补充剂有助于减缓老年人的肠道微生物组成变化,以及免疫功能的下降,从而降低年龄相关疾病的发生。然而,益生菌对哺乳动物长寿效应的影响还需要进一步的研究。
猜你喜欢
今日农业(2022年14期)2022-09-15
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
猪业科学(2018年5期)2018-07-17
中成药(2018年2期)2018-05-09
幸福(2017年18期)2018-01-03
饲料与畜牧(规模养猪)(2017年9期)2017-10-16
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
中国乳品工业(2017年1期)2017-02-20
天然产物研究与开发(2016年6期)2016-06-05
