
广东石门台国家级自然保护区老虎山次生林群落特征
2023-03-09 01:52饶兴权曾阳金李远球曹洪麟刘占锋蔡锡安
热带亚热带植物学报 2023年1期
饶兴权, 曾阳金, 李远球, 曹洪麟, 刘占锋, 蔡锡安*
广东石门台国家级自然保护区老虎山次生林群落特征
饶兴权1,2, 曾阳金3, 李远球3, 曹洪麟1,2, 刘占锋1,2, 蔡锡安1,2*
(1. 中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广州 510650;2. 华南国家植物园, 广州 510650; 3. 广东石门台国家级自然保护区管理局,广东 英德 513000)
为了解在广东石门台国家级自然保护区的老虎山南坡建设的大型林冠模拟氮沉降野外控制试验平台的植被组成特征,开展了18个样方的每木调查和分析研究。结果表明,在面积0.72 hm2中有维管束植物53科86属127种;胸径(DBH)≥1.0 cm的立木有2 465株,平均密度为0.34 ind./m2,最高达25 m。群落以锥()、罗伞树()、木荷()等为主要优势种类,且以高位芽、中小型叶、革质和非全缘叶植物为主要种类组成的常绿阔叶次生林。热带-亚热带区系成分在群落中占明显优势,具有热带向亚热带过渡的特征。18个样方乔木层的Simpson指数(0.851~0.936)和Shannon-Wiener指数(3.504~4.494)相近,Pielou均匀度指数较高(0.780~1.000),但Gleason和Margalef丰富度指数差异较大(分别为3.209~7.738和16.760~40.806),样方间有10~26种共有种(Sorensen相似性系数为0.377~0.722),而灌木层则相反,物种多样性指数均变化较大,样方间只有0~10种共有种(Sorensen相似性系数为0~0.714)。因此,灌木层的密度和种类差异对群落结构的影响应充分考虑。
常绿阔叶林;区系成分;物种组成;生物多样性;石门台自然保护区;广东
广东石门台国家级自然保护区地处北回归线北缘,属南亚热带与中亚热带过渡地区。该保护区的主要保护对象是南亚热带季风常绿阔叶林向中亚热带典型常绿阔叶林过渡的森林生态系统[1]。保护区生物多样性丰富,植被类型多种多样,早在20世纪30年代, 就有学者陆续对该地区进行标本采集和植被调查研究[1–4],并开展典型的森林群落结构、种类组成、植物多样性等研究[1,5–10]。2012年中国科学院华南植物园在保护区的老虎山南坡建设大型林冠模拟氮沉降野外控制试验平台(简称氮沉降平台),拟通过林冠及林下人工施氮处理,研究模拟大气氮沉降增加对常绿阔叶森林生态系统结构和功能的影响[11]。
常绿阔叶林蕴藏着丰富的生物资源,对保护环境和维持全球碳平衡都具有极重要的作用。20世纪30年代始,我国就开展了常绿阔叶林的研究,特别是新中国成立后多次成规模的植被调查和研究,这为了解我国常绿阔叶林植物区系的性质与特点、地理分布、物种多样性分布规律等提供了重要的科学依据[12–13]。后期科研人员在群落动态监测,局部地区的植物科属成分、区系性质与起源,不同植物类群的起源、扩散与分布,生物多样性的形成与维持机制,生态功能与服务等方面做了大量的研究[14–16]。随着人们对全球变化等问题的关注,以及遥感、人工智能、无人机和大数据等新技术的应用,常绿阔叶林的研究迎来了新的发展机遇[16]。在老虎山次生林设立氮沉降试验平台,研究模拟大气氮沉降增加对常绿阔叶林的影响,有助于认识热带亚热带地区常绿阔叶林的生物多样性维护机制,以及全球变化对其碳汇功能的影响。
群落的物种组成与结构特征是生态系统功能和过程的基础,对群落物种组成与结构的分析可以进一步揭示物种多样性的形成和维持机制[17]。植物物种多样性影响着生态系统生产力、稳定性、可入侵性以及养分动态[18]。利用物种多样性指数可以描述群落的结构特征,比较群落的复杂性,物种的丰富度和均匀度。植物的叶片、胸径、株高等这些属性,能够反映植物对生长环境的响应和适应,能将环境、植物个体和生态系统结构、过程与功能联系起来,常常被作为衡量植物特征的方式[19–22]。
研究表明,氮沉降增加已悄然改变了区域甚至全球氮循环格局[23–24],氮沉降增加不可避免地影响着常绿阔叶林的物种组成、外貌、结构和生态功能等属性[25]。因此,开展氮沉降增加对常绿阔叶林群落的物种组成、结构与功能的影响,以及植物性状如叶型特征,径级结构,株高等属性的影响,可有效地揭示常绿阔叶林对氮素输入量增加的响应与适应。
对于控制试验来说,样方间植物的丰富度、分布均匀度、植株密度、立木结构等群落特征的差异会影响试验结果,了解样方的群落基本特征是研究的基础。然而,该氮沉降试验平台的植物区系的性质、物种组成、植物属性和样方间的物种相似性等特征还未见报道,其常绿阔叶林群落是否具有南亚热带与中亚热带过渡的性质也需要进一步调查分析。因此,本文在样方每木调查基础上,进行群落的种类组成、生活型谱、叶型特征、立木径级结构、区系成分等的分析,并开展样方间的生物多样性指数,包括Simpson指数和Shannon-Wiener指数、Gleason和Margalef物种丰富度指数、Pielou均匀度指数和Sorensen物种组成相似性系数等指标的分析,以检验样方间植物群落基本特征的差异,为后续研究提供参考。
1 研究地概况
广东石门台国家级自然保护区位于广东省中北部(113°01′11″~113°46′22″ E, 24°17′49″~24°31′02″N),南岭山脉的最南端,面积约822.6 km2,为广东省最大的森林生态系统自然保护区,是珠江三角洲北部的绿色屏障[1]。该保护区地处南亚热带与中亚热带过渡地带,属于亚热带季风气候,年均温20.9 ℃,年均降水量为1 882.8 mm,降水主要集中在4—8月间[1]。
本调查样地为保护区外延老虎山南坡,距锦潭保护站约7 km,海拔为200~400 m,面积约2 hm2,现存植被为常绿阔叶次生林,林分曾遭受过人为选择性砍伐,少见大径材个体,样地内的锥()、木荷()等多为砍伐后萌生个体。土壤类型为酸性山地黄壤,成土母质是花岗岩。
该群落外貌终年常绿,一些高大树木突出林冠之上,不连续,呈伞状冠型。由春到秋,群落冠层呈嫩绿到深绿,在冬季老叶脱落前,常可见部分红叶映衬在绿色冠层中。冬季可见落叶树种枫香树()等叶片变黄、红色(图1)。

图1 群落冠层外貌
2 方法
2.1 样地设置和调查方法
参考BCI大样地建设的技术规范[26],在样地内设18个20 m×20 m样方,在每个样方内细分为4个10 m×10 m和16个5 m×5 m的小样方。以5 m× 5 m小样方为基本测树单元,挂牌标记每个胸径(diameter at breast height, DBH)≥1 cm,高度(height, H)>1.5 m的个体,记录树种名、树高、胸径、冠幅、坐标和生长状况等信息[27]。每个样方内选择1个5 m× 5 m小样方调查林下DBH<1 cm、H为50~150 cm的灌木植物,记录种类、树高、地径、冠幅和坐标。在5 m×5 m小样方内,选择3个1 m×1 m小样方调查H<50 cm的草本层植物(包括小苗),记录其类、株数(丛数)、株高、盖度和坐标。在调查乔木的同时调查层间植物,记录种类、株数、高度和盖度。另外,在5 m×5 m小样方内还调查幼苗更新状况[28]。
2.2 数据处理
参考曲仲湘等[29]的方法,将径级结构分为5级: H<33 cm为Ⅰ级、H≥33 cm且DBH<2.5 cm为Ⅱ级、DBH在2.5~7.5 cm为Ⅲ级、DBH在7.5~22.5 cm为Ⅳ级、DBH≥22.5 cm为Ⅴ级。
重要值(IV)=(相对多度+相对频度+相对显著度)/3×100,Simpson指数=1-Σ(/)2;Shannon- Wiener指数=Σln,式中i=i/,i为种的个体数,为群落中全部物种的个体数。为种的个体数占全部个体的比例。
Gleason指数=/ln;Margalef指数=(-1)/ ln,Pielou均匀度指数max,Sorensen相似性系数s=2/(+),式中,为单位面积,为群落中的物种数目,为观察到的个体总数,为实际观察的物种多样性指数,max为最大的物种多样性指数,max=ln,、分别为2样方的物种数,为2样方共有的物种数。
3 结果和分析
3.1 种类组成
18个样方7 200 m2共有维管束植物53科86属127种(表1),其中蕨类植物6科7属7种,裸子植物1科1属1种,被子植物46科78属119种。双子叶植物40科71属111种,单子叶植物6科7属8种。含3种以上的有11科,占总科数的20.8%,这些科所含种数占总种数的59.1%。本群落有32科只含1种,占总科数的61.4%。还有国家2级保护植物金毛狗()和黑桫椤(), 及国家珍稀植物粘木()。

表1 维管束植物统计
从表2可见,乔木层重要值(IV)>2.0的有19种,占乔木层物种的79.00%,IV排前10位的种类为锥(11.69)、罗伞树(, 9.60)、木荷(6.60)、柏拉木(, 4.78)、多花山矾(, 4.15)、鼎湖血桐(, 4.12)、鸭脚木(, 4.12)、粘木(3.86)、黄杞(, 3.83)和华润楠(, 3.54)。
从表3可见,灌木层(DBH<1.0 cm,H>50 cm)有51种植物,IV>2.0的有12种,占灌木层物种数的76.13%,其余39种为23.87% (表3)。IV排前3位的分别为罗伞树(37.67)、九节(, 6.98)和柏拉木(4.94)。灌木层罗伞树占较大优势, 其次是九节、柏拉木、粗叶木()和香楠()。
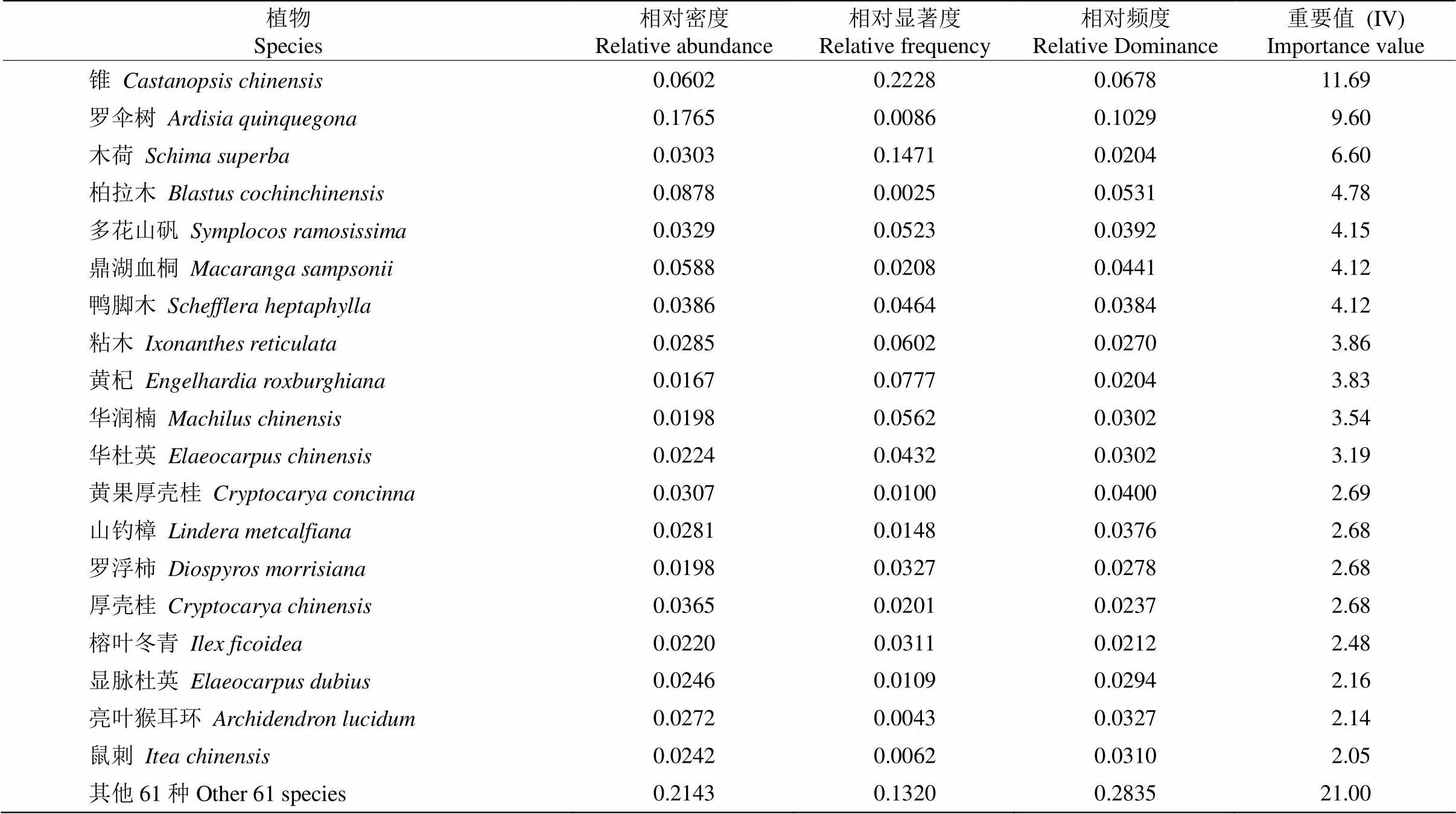
表2 乔木层种类重要值(IV)
3.2 区系特征
按吴征镒[32]的中国种子植物区系类型的划分, 群落内77属种子植物可分为12个分布区类型(表4)。世界分布型2属,热带分布型65属(86.67%),其中以泛热带分布(37.33%)、旧世界热带分布(18.67%)和热带亚洲分布(14.67%)为主。除栗属为东亚和北美间断分布外,重要值排前10的物种有9属都为热带分布,如紫金牛属、木荷属和柏拉木属等。东亚和北美洲间断分布有5属(6.67%), 如栗属、枫香树属、木犀属等。北温带分布只有1属为荚蒾属(1.33%)。林中还有少量棕榈科植物杖藤(),以及茎花、板根等热带雨林特征,但发育不显著。

表3 灌木层植物的重要值(IV)

表4 种子植物属的区系地理成分
不包括世界分布属。
Cosmopolitan genera are not include.
3.3 物种的生活型谱
按C. Raukiaer生活型分类系统[33],本群落以高位芽植物为主,约有102种, 占总种数的80.31%,藤本高位芽植物16种, 占12.60%,地面芽有5种,一年生植物3种,地下芽植物仅1种(表5)。未见有大高位芽、附生高位芽和地上芽植物。中高位芽71种(55.90%),小高位芽28种,矮高位芽仅3种。林下草本层以蕨类植物为主,大部分为地面芽植物,同时还有少量一年生草本植物。藤本植物以木质藤本为主, 多呈灌丛状,少见大型木质藤本。
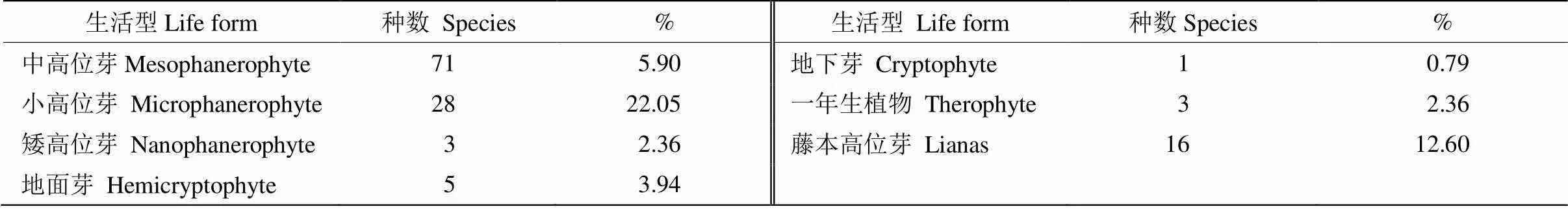
表5 群落植物的生活型
3.4 物种的叶型特征
群落以中、小型叶种类为主,未见大型叶和微型叶种类。单叶植物有107种,占总数的84.25%, 复叶有20种,占15.75%。乔木复叶种类主要有橄榄()、黄杞、猴耳环()等。蕨类和藤本复叶种类有金毛狗、黑桫椤、异叶鳞始蕨()、崖豆藤()、木通()和小叶红叶藤()等。革质叶植物72种(56.69%), 纸质叶51种(40.16%),膜质叶4种(3.15%); 全缘叶47种(37.00%),非全缘叶80种(63.00%),以锯齿叶种类占多数。
3.5 优势种立木径级结构
对重要值排前10种的立木径级结构分析表明,木荷和粘木的径级分布呈倒金字塔型,它们的Ⅰ、Ⅱ级树苗较少,个体主要分布在Ⅳ和Ⅴ的大径级上(表6)。锥、多花山矾、鸭脚木、黄杞和华润楠在各个径级上都有分布,在群落中表现为稳定增长型。罗伞树、柏拉木和鼎湖血桐在Ⅰ、Ⅱ和Ⅲ级中有较大数量的个体,但在Ⅳ和Ⅴ大径级上很少或没有分布,表现为金字塔型。

表6 乔木层重要值前10位物种径级分布
N: 数量; Ⅰ: H<50 cm; Ⅱ: DBH<2.5 cm, H>50 cm; Ⅲ: DBH 2.5~7.5 cm; Ⅳ: DBH 7.5~22.5 cm; Ⅴ: DBH>22.5 cm。
N: Number; Ⅰ: H<50 cm; Ⅱ: DBH<2.5 cm, H>50 cm; Ⅲ: DBH 2.5~7.5 cm; Ⅳ: DBH 7.5~22.5 cm; Ⅴ: DBH>22.5 cm.
3.6 垂直结构
在7 200 m2的样方中,乔木81种,高于1.5 m的立木2 465株,平均密度为0.34 ind./m2,最高达25 m。
本群落可划分为乔木层、灌木层和草本层等3个层次,地被层少见,此外还有一些层间植物。乔木层可进一步分为2个亚层,第一亚层18~25 m, 胸径20~50 cm,覆盖度15%~30%,树冠呈不连续分布,以锥、木荷占多数,但个体都不大,多数是遭砍伐后的萌生个体,丛生明显。最高的个体是木荷,高达25 m,胸径达52.6 cm,另外黄杞、华润楠、橄榄和枫香树等的胸径也较大。第二亚层高度为2~16 m,胸径为1.0~25 cm,覆盖度65%~80%,除林窗外,树冠较连续,密度较大,个体较小,主要种类为多花山矾、鼎湖血桐、鸭脚木、粘木、华润楠、罗伞树等。
灌木层明显,盖度为30%~40%,高50~150 cm,但冠层不连续,多呈团块状分布。灌木层植物种类较多,多数为乔木树种的幼树,如黄果厚壳桂、厚壳桂、罗伞树等。常见的灌木种类有九节、溪边九节()、红背山麻杆()、算盘子()、柏拉木和野牡丹()等。
草本层也不发达,盖度为5%~10%,高20~ 50 cm,主要由乔、灌木层的幼苗、蕨类和少量的草本植物组成,如罗伞树、金毛狗、乌毛蕨和扇叶铁线蕨等。另外,华山姜()和黑莎草()也较常见。
层间植物的种类和数量均较少,主要有崖豆藤、大叶白纸扇()、杖藤、薯莨()、海金沙()等18种。除大叶白纸扇、木通能达到乔木冠层外,其余一般呈灌状丛生或在低矮的灌木间盘绕,群落中未见有附生植物。
3.7 生物多样性和相似性
从表7可见,18个样方乔木层的Simpson指数为0.851~0.936,最小的是14号样方,最大的是16号样方; Shannon-Wiener指数为3.504~4.494,最小的是14号样方,最大的是10号样方。Gleason丰富度指数为3.209~7.738,Margalef丰富度指数为16.760~40.806,均是6号样方最小,10号样方最大。Pielou均匀度指数为0.780~1.000,最小的是14号样方,最大的是10号样方。
18个样方灌木层的多样性指数变化较大, Simpson指数为0.537~0.918,Shannon-Wiener指数为1.654~ 3.958;最小的均是17号样方,分别为0.537和1.654,最大的是16号样方,分别为0.918和3.958。Gleason和Margalef丰富度指数分别为1.864~7.145和5.719~22.772,以17号样方最小,2号样方最大。Pielou均匀度指数为0.418~1.000,以17号样方的最小, 16号样方的最高。
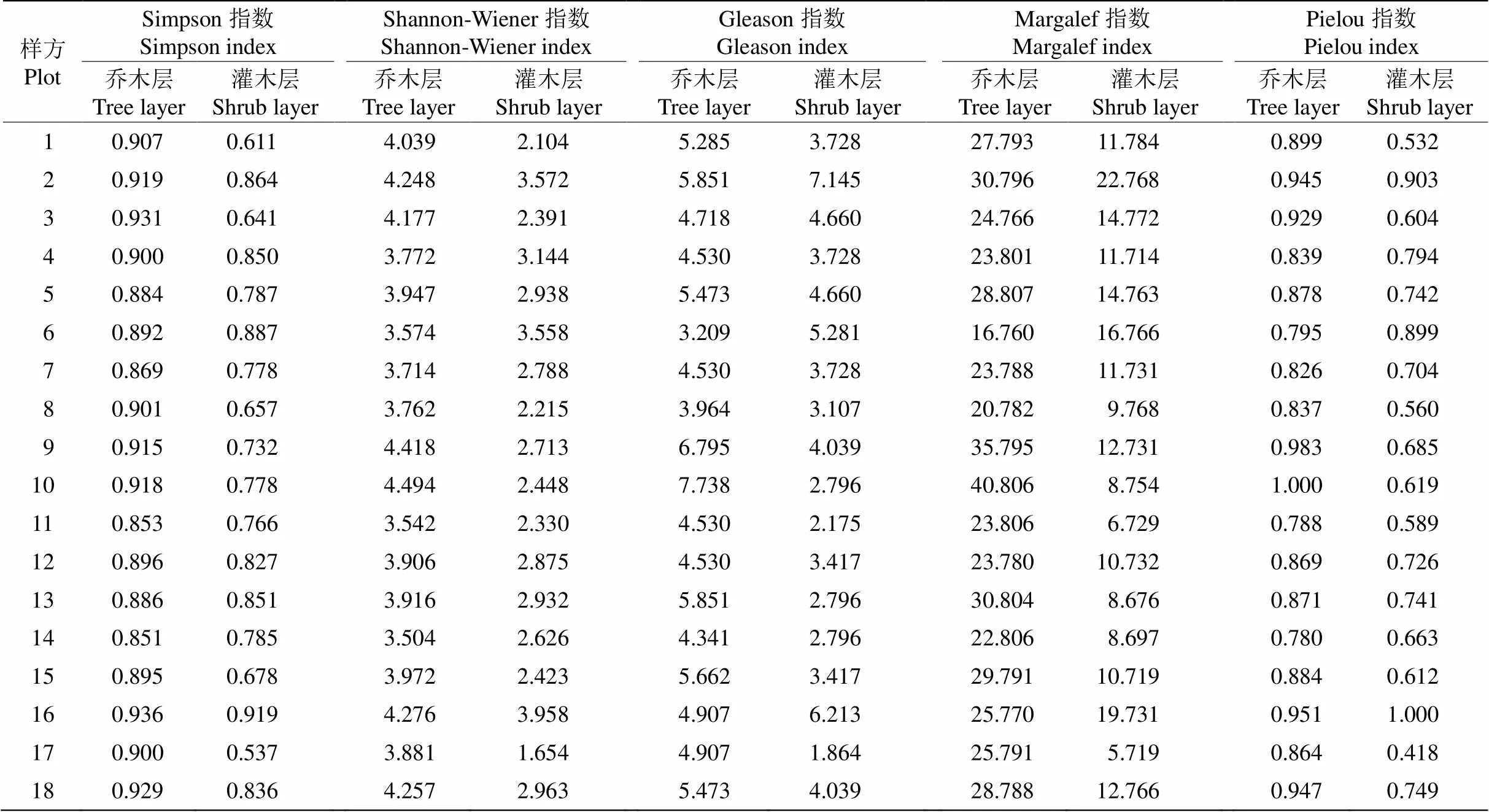
表7 乔灌木层的植物多样性指数
从表8可见,18个样方乔木层的相似性系数为0.377~0.722,最高的是10与13号样方,有26种个共同种;最低的是6与9号样方,有10种共同种。灌木层相似性系数为0~0.714,最大的是5与18号样方(0.714),有10种共同种,最低的是4与17号样方,没有共同种。
4 结论和讨论
广东石门台国家级自然保护区老虎山南坡植物群落的0.72 hm2样地中,有维管植物53科86属127种,乔木层主要优势植物为锥、罗伞树、木荷等,灌木层主要为罗伞树、九节、柏拉木、溪边九节、红背山麻杆等。本群落没有绝对优势的植物种类,较高大乔木主要是锥、木荷和黄杞等种。虽然罗伞树、鼎湖血桐和鸭脚木等种类有一定数量的个体, 但多数仅居于乔木的亚层,个体较矮小,无法形成顶层优势。结合历史走访调查和本群落缺乏高大乔木个体的现实,本群落为人为干扰后发展成的常绿阔叶次生林,可命名为锥-罗伞树常绿阔叶次生林。

表8 18个样方乔木层(右上)和灌木层(左下)相似性系数
以往研究表明,石门台自然保护区的森林植被以热带、亚热带的科属为代表,优势科有壳斗科、山茶科、樟科、杜英科、木兰科等[5–6]。本群落优势科植物组成与保护区内的其他样地的研究结果相似[6,9],都以壳斗科、紫金牛科、山茶科等为优势科,但本群落中木兰科的优势地位并不明显。另外,本群落的优势科属与典型的南亚热带和中亚热带最具优势代表的科属相比,有相似也存在差异[27,34–35]。如杜鹃花科、木兰科、金缕梅科和清风藤科等在本群落中很少见,保护区内的其他样地中常见的柿属、鼠刺属以及其所在的柿科和鼠刺科也不是本群落的优势科属[9],这可能与样地的地理位置、人为干扰程度等有关。本群落的主要科属组成(表1)都是本地区的主要分布科属,其主要组成种类也都是南亚热带常绿阔叶林的常见种类成分[27]。其比邻近地区广东南雄青嶂山常绿阔叶林1 hm2固定样地的物种相对丰富一些(起测胸径1.0 cm,有38科58属74种)[34],但比车八岭和鼎湖山常绿阔叶林20 hm2样地的植物种类少,这可能与地理位置和调查面积有关[27,35]。
从区系特征来看,虽然本群落物种以热带成分占较大优势,但其茎花、板根等热带雨林特征发育不显著,同时也含有少量的温带成分,如忍冬科的荚蒾属,以及金缕梅科的枫香树等。从生活型谱看, 本群落高位芽植物占多数,草本植物少,藤本植物也多为灌丛状,未见有大高位芽、附生高位芽和地上芽植物。群落叶型特征也以中、小型革质锯齿叶为主,未见有大型叶和微型叶种类。这些区系特征、生活型谱和叶型特征都与石门台大西山的粘木-甜锥群落相似[6],也与相邻的八宝山和南昆山的常绿阔叶林相似(本群落高位芽植物占92.91%,大西山粘木-甜锥群落高位芽植物占88.5%,八宝山常绿阔叶林群落占83.3%,南昆山常绿阔叶林群落占89.5%)[6,36–37]。因此,群落的区系成分、物种组成、生活型谱以及叶型特征等均表明本群落植物区系性质具有南亚热带向中亚热带过渡的特点。
重要值排前10种的立木径级结构分析表明, 木荷和粘木径级分布呈倒金字塔型,随着时间的推移,这2种可能会退出群落优势种群,而锥、多花山矾、鸭脚木、黄杞和华润楠为稳定增长型,这些种将会成为群落的主要建群种。虽然罗伞树、柏拉木和鼎湖血桐的径级分布呈正金字塔型,但由于其生物和生理特性等的原因,它们没法成长为大径级个体,难于成为群落的顶层优势种。因此,除木荷和粘木外,群落主要种类都属于增长型种群,群落处于进展演替阶段。
综上,广东石门台国家级自然保护区老虎山南坡植物群落以锥、罗伞树、木荷等主要优势种类的常绿阔叶次生林为主,其具有南亚热带向中亚热带过渡的植物区系性质特征,群落处于进展演替阶段。18个样方中乔木层的物种数和个体数量分布较均匀,相对密度较接近,样方间物种组成较接近。而灌木层则相反,样方间的物种数和个体数量分布较不均匀,相对密度差异较大,样方间物种组成差异大。以后开展实验时,应充分考虑灌木层种类和密度差异的影响。
致谢 平台建设和野外调查工作得到广东石门台国家级自然保护区管理局及其下属单位锦潭管理站工作人员的大力支持。林冠外貌照片由孙中宇拍摄。参加野外植被调查人员还有陈炳辉、吴林芳、曾小平、朱师丹、贺鹏程、赵秀华、时雷雷、王敏、李荣华、陈小花、张振振等,特此致谢。
[1] ZHANG J Q. Comprehensive Investigation Report of Shimentai National Nature Reserve in Guangdong Province [M]. Wuhan: Huazhong University of Science and Technology Press, 2017: 1–101. [张金泉. 广东石门台国家级自然保护区综合科学考察报告 [M]. 武汉: 华中科技大学出版社, 2017: 1–101.]
[2] XU X H, ZHONG Z C, WANG L Z, et al. The plant community of Yingde Huashui Mountain, Guangdong Province [J]. Chin J Plant Ecol, 1958, 2(1): 1–59. [徐祥浩, 钟章成, 王灵昭, 等. 广东英德滑水山的植物群落 [J]. 植物生态学报, 1958, 2(1): 1–59.]
[3] Zhang Hongda Anthology Editotial Group. Anthology of Zhang Hong Da [M]. Guangzhou: Sun Yat-Sen University Press, 1995: 676–714. [《张宏达文集》编辑组. 张宏达文集 [M]. 广州: 中山大学出版社, 1995: 676–714.]
[4] XU Y Q. The important signicance of establishing nature reserve in Yingde municipality [J]. Ecol Sci, 1998, 17(2): 107–110. [徐燕千. 建立英德市石门台自然保护区的重大意义 [J]. 生态科学, 1998, 17(2): 107–110.]
[5] SU Z Y, CHEN B G, WU D R. Vegetation types and community structure of Shimentai Nature Reserve, Yingde, Guangdong [J]. J S China Agric Univ, 2002, 23(1): 58–62. [苏志尧, 陈北光, 吴大荣. 广东英德石门台自然保护区的植被类型和群落结构 [J]. 华南农业大学学报, 2002, 23(1): 58–62. doi: 10.3969/j.issn.1001-411X.2002.01. 017.]
[6] CHEN H F, YAN Y H, XING F W, et al. A study on characteristics of-community in Shimentai Nature Reserve, Guangdong [J]. Guihaia, 2003, 23(6): 488–494. [陈红锋, 严岳鸿, 邢福武, 等. 广东石门台自然保护区粘木-甜锥群落特征研究 [J]. 广西植物, 2003, 23(6): 488–494. doi: 10.3969/j.issn.1000-3142. 2003.06.002.]
[7] CHEN H F, YAN Y H, XING F W, et al. Survey on original vegetation in Shimentai Nature Reserve, Guangdong [J]. J S China Agric Univ (Nat Sci), 2003, 24(2): 22–26. [陈红锋, 严岳鸿, 邢福武, 等. 广东石门台自然保护区原生植被的调查研究 [J]. 华南农业大学学报(自然科学版), 2003, 24(2): 22–26. doi: 10.3969/j.issn.1001-411X.2003.02. 007.]
[8] MIAO S Y, WANG W T, ZENG Y J, et al. Basic characteristics ofcommunity in Shimentai Nature Reserve, Guangdong [J]. Guihaia, 2004, 24(5): 390–395. [缪绅裕, 王伟彤, 曾阳金, 等. 广东石门台自然保护区广东松群落的基本特征 [J]. 广西植物, 2004, 24(5): 390–395. doi: 10.3969/j.issn.1000-3142.2004.05. 002.]
[9] LI Y Q, HUANG L S, MIAO S Y, et al. Species diversity of the forest vegetation in Shimentai Natural Reserve, Guangdong [J]. J Fujian For Sci Technol, 2018, 45(3): 13–18. [李远球, 黄林生, 缪绅裕, 等. 广东石门台自然保护区森林植被物种多样性 [J]. 福建林业科技, 2018, 45(3): 13–18. doi: 10.13428/j.cnki.fjlk.2018.03.003.]
[10] HUANG L S, LI Y Q, DAI K Y, et al. Dominant populations and their age structures of forest vegetation in Shimentai, Guangdong [J]. Subtrop Plant Sci, 2018, 47(2): 144–148. [黄林生, 李远球, 戴克元, 等. 广东石门台森林植被的优势种群及其年龄结构 [J]. 亚热带植物科学, 2018, 47(2): 144–148. doi: 10.3969/j.issn.1009-7791.2018.02.010.]
[11] ZHANG W, SHEN W J, ZHU S D, et al. CAN canopy addition of nitrogen better illustrate the effect of atmospheric nitrogen deposition on forest ecosystem? [J]. Sci Rep, 2015, 5(1): 11245. doi: 10.1038/ srep11245.
[12] Editorial Board of the Vegetation of China. Vegetation of China [M]. Beijing: Science Press, 1980: 3–1144. [中国植被编辑委员会. 中国植被 [M]. 北京: 科学出版社, 1980: 3–1144.]
[13] SONG Y C, CHEN X Y, WANG X H. Studies on evergreen broad-leaved forests of China: A retrospect and prospect [J]. J E China Norm Univ (Nat Sci), 2005(1): 1–8. [宋永昌, 陈小勇, 王希华. 中国常绿阔叶林研究的回顾与展望 [J]. 华东师范大学学报(自然科学版), 2005(1): 1–8. doi: 10.3969/j.issn.1000-5641.2005.01.001.]
[14] WANG X H. Phytogeography and species diversity of typical evergreen broad-leaved forest in China [D]. Shanghai: East China Normal University, 2006. [王希华. 中国典型常绿阔叶林植物地理与物种多样性研究 [D]. 上海: 华东师范大学, 2006.]
[15] MI X C, FENG G, ZHANG J, et al. Review on biodiversity science in China [J]. Bull Chin Acad Sci, 2021, 36(4): 384–398. [米湘成, 冯刚, 张健, 等. 中国生物多样性科学研究进展评述 [J]. 中国科学院院刊, 2021, 36(4): 384–398. doi: 10.16418/j.issn.1000-3045.20210307 002.]
[16] MA K P, GUO Q H. Progress and recent trends of vegetation ecology in China [J]. Sci Sin Vitae, 2021, 51(3): 215–218. [马克平, 郭庆华. 中国植被生态学研究的进展和趋势 [J]. 中国科学: 生命科学, 2021, 51(3): 215–218. doi: 10.1360/SSV-2021-0010.]
[17] LOREAU M, NAEEM S, INCHAUSTI P, et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges [J]. Science, 2001, 294(5543): 804–808. doi: 10.1126/science.1064088.
[18] CADOTTE M W, CARSCADDEN K, MIROTCHNICK N. Beyond species: Functional diversity and the maintenance of ecological pro- cesses and services [J]. J Appl Ecol, 2011, 48(5): 1079–1087. doi: 10. 1111/j.1365-2664.2011.02048.x.
[19] MCINTYRE S, LAVOREL S, LANDSBERG J, et al. Disturbance response in vegetation: Towards a global perspective on functional traits [J]. J Veg Sci, 1999, 10(5): 621–630. doi: 10.2307/3237077.
[20] CORNELISSEN J H C, LAVOREL S, GARNIER E, et al. A handbook of protocols for standardized and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 2003, 51(4): 335–380. doi: 10.1071/ BT02124.
[21] MENG T T, NI J, WANG G H. Plant functional traits, environments and ecosystem functioning [J]. J Plant Ecol, 2007, 31(1): 150–165. [孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能 [J]. 植物生态学报, 2007, 31(1): 150–165. doi: 10.17521/cjpe.2007.0019.]
[22] LIU X J, MA K P. Plant functional traits: Concepts, applications and future directions [J]. Sci Sin Vitae, 2015, 45(4): 325–339. [刘晓娟, 马克平. 植物功能性状研究进展 [J]. 中国科学: 生命科学, 2015, 45 (4): 325–339. doi: 10.1360/N052014-00244.]
[23] GRUBER N, GALLOWAY J N. An earth-system perspective of the global nitrogen cycle [J]. Nature, 2008, 451(7176): 293–296. doi: 10. 1038/nature06592.
[24] SULLIVAN B W, SMITH W K, TOWNSEND A R, et al. Spatially robust estimates of biological nitrogen (N) fixation imply substantial human alteration of the tropical N cycle [J]. Proc Natl Acad Sci USA, 2014, 111(22): 8101–8106. doi: 10.1073/pnas.1320646111.
[25] CLARK C M, TILMAN D. Loss of plant species after chronic low- level nitrogen deposition to prairie grasslands [J]. Nature, 2008, 451 (7179): 712–715. doi: 10.1038/nature06503.
[26] CONDIT R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island Panama and a Comparison with Other Plots [M]. New York: Springer, 1998: 1–181.
[27] YE W H, CAO H L, HUANG Z L, et al. Community structure of a 20 hm2lower subtropical evergreen broadleaved forest plot in Dinghu- shan, China [J]. J Plant Ecol, 2008, 32(2): 274–286. [叶万辉, 曹洪麟, 黄忠良, 等. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究 [J]. 植物生态学报, 2008, 32(2): 274–286. doi: 10.3773/j.issn. 1005-264x.2008.02.005.]
[28] DONG M. Survey, Observation and Analysis of Terrestrial Biocom- munities [M]. Beijing: Standards Press of China, 1996: 1–23. [董鸣. 陆地生物群落调查观测与分析 [M]. 北京: 中国标准出版社, 1996: 1–23.]
[29] QU Z X, WEN Z W, ZHU K G. An analytical study of the forest of the spirit valley, Nanking [J]. Acta Bot Sin, 1952, 1(1): 18–49. [曲仲湘, 文振旺, 朱克贵. 南京灵谷寺森林现况的分析 [J]. 植物学报, 1952, 1(1): 18–49.]
[30] WU D X, ZHANG L, SONG C Y, et al. Protocols for Standard Biological Observation and Measurement in Terrestrial Ecosystems [M]. Beijing: China Environmental Science Press, 2019: 34–508. [吴冬秀, 张琳, 宋创业, 等. 陆地生态系统生物观测指标与规范 [M]. 北京: 中国环境出版集团, 2019: 34–508.]
[31] MA K P, LIU C R, LIU Y M. The methods of measuring community biodiversity: II. Measurement ofdiversity [J]. Chin Biodiv, 1995, 3(1): 38–43. [马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法: II.多样性的测度方法[J]. 生物多样性, 1995, 3(1): 38–43.]
[32] WU Z Y. The area types of Chinese genera of seed plants [J]. Acta Bot Yunnan, 1991, 13(S4): 1–139. [吴征镒. 中国种子植物属的分布区类型 [J]. 云南植物研究, 1991, 13(S4): 1–139.]
[33] Raunkiaer C. The Life Forms of Plants and Statistical Plant [M]. Oxford: Geography Clarendon Press, 1934:1–632.
[34] DAI W T, CHEN W L, MIAO S Y, et al. Analysis on the composition and structure of dominant species within an evergreen broadleaved forest in Qingzhangshan, Nanxiong [J]. For Environ Sci, 2017, 33(2): 8–13. [戴文坛, 陈伟霖, 缪绅裕, 等. 南雄青嶂山常绿阔叶林优势物种组成与结构分析 [J]. 林业与环境科学, 2017, 33(2): 8–13. doi: 10.3969/j.issn.1006-4427.2017.02.002.]
[35] WU Z H, WANG Z, LUAN F C, et al. Community composition and floral characteristics of the Chebaling 20 hm2forest dynamic plot in a mid-subtropical evergreen broad-leaved forest [J]. For Environ Sci, 2021, 37(3): 86–91. [吴智宏, 王梓, 栾福臣, 等. 车八岭20 hm2中亚热带常绿阔叶林监测样地群落物种组成和区系特征 [J]. 林业与环境科学, 2021, 37(3): 86–91.]
[36] CHEN Z H, LI M G, LÜ X H, et al. A study on the forest communities in Natural Reserve of Nankunshan, Guangdong [J]. Ecol Sci, 1983, 2(1): 18–29. [陈章和, 李鸣光, 吕小红, 等. 广东南昆山自然保护区森林群落 [J]. 生态科学, 1983, 2(1): 18–29.]
[37] ZHANG J Q. Vegetation characters of the Ruyang Babao Mountain Nature Reserve, Guangdong Province [J]. Ecol Sci, 1993, 12(1): 39– 124. [张金泉. 广东乳阳八宝山自然保护区的植被特点 [J]. 生态科学, 1993, 12(1): 39–124.]
Community Characteristics of Tiger Mountain Secondary Forest in Shimentai National Nature Reserve, Guangdong
RAO Xingquan1,2, ZENG Yangjin3, LI Yuanqiu3, CAO Honglin1,2, LIU Zhanfeng1,2, CAI Xi’an1,2*
(1.South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510160,China; 2. South China National Botanical Garden, Guangzhou 510650, China; 3.Administrative Bureau of Shimentai Nature Reserves,Yingde 513000, Guangdong, China)
A large canopy simulation and nitrogen deposition field control experimental platform was established on southern slope of Tiger Mountain in Shimentai National Nature Reserve, Guangdong Province, China. To further understand the characteristics of vegetation composition at the platform, 18 plots covering a total area of 0.72 hm2were set up. The results showed that there were 127 vascular species, belonging to 86 genera and 53 families. Furthermore, there were 2 465 individuals with diameter at breast height (DBH)≥1.0 cm. The tree density was 0.34 ind./m2, and the tallest up to 25 m. The dominant species in the community was,, and, which was a secondary evergreen broad-leaved forest consisting of phanerophytes, medium- and small-leaf, leathery, and unentire leaf as main species. The tropical-subtropical floristic elements were dominant in the community, with the characteristics of transition from the tropical to subtropical. Among 18 plots at tree layer, the Simpson index (0.851-0.936) and Shannon-Wiener index (3.504-4.494) were similar; the Pielou’s evenness index was high (0.780-1.000); and the Gleason and Margalef richness indexes were rather variable (3.209-7.738 and 16.760-40.806, respectively). The plots shared 10-26 specieswith Sorensen similarity coefficient of 0.377-0.722. On the contrary, in shrub layers, the above indexes all varied greatly, and only 0–10 common species among plots with Sorensen similarity coefficient of 0- 0.714. Therefore, the effect of shrub layer density and species difference on community structure should be considered.
Evergreen broad-leaved forest; Flora element; Species composition; Biodiversity; Shimentai Nature Reserve; Guangdong
10.11926/jtsb.4560
2021-11-05
2022-05-09
国家自然科学基金项目(42177289)资助
This work was supported by the National Natural Science Foundation of China (Grant No.42177289).
饶兴权(1978年生),男,工程师,主要从事植物生态学研究。E-mail: rxq99@scbg.ac.cn
E-mail: xncai@scbg.ac.cn
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
林业勘查设计(2022年1期)2022-02-15
广西林业科学(2021年6期)2022-01-21
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
绿色中国(2019年18期)2020-01-04
农家科技下旬刊(2017年8期)2017-11-13
西北植物学报(2015年9期)2015-07-05
