
国内外萱草新品种选育研究进展
2023-03-09 09:48段连峰刘晓娜于守超
核农学报 2023年4期
段连峰 李 颖 刘晓娜 董 燕 于守超 李 俊
(1北京市园林绿化科学研究院,绿化植物育种北京市重点实验室,北京 100102;2聊城大学农学与农业工程学院,山东 聊城 252059;3山东英才学院,山东 济南 250000;4中国农业科学院农产品加工研究所,北京 100193;5中国花卉协会,北京 100102)
萱草(HemerocallisL.)又名忘忧草、宜男草,在中国传统文化中有孝亲、忘忧、宜男多子的寓意,是阿福花科(Asphodelaceae)多年生草本植物。萱草作为世界三大宿根花卉之一,在园林中应用广泛,尤其在中国古典园林中常用于点缀山石。16世纪传入欧洲,至19世纪末,萱草已成为欧美庭院中广泛应用的园林植物[1]。
萱草属植物还是一类食药兼顾的植物。萱草属植物主要的食用品种是黄花菜,其花蕾经过蒸、晒加工成菜品,含有丰富的蛋白质、脂类、糖类、维生素等营养物质,有“素山珍”的美称[2]。萱草属植物的根、茎、叶、花均可入药,最早在《神农本草经》中便有记载。截至2020年12月31日的统计数据显示,在萱草属植物中,共发现266种次生代谢产物[3-4],包括110种挥发性成分、41 种黄酮类、39 种生物碱、29 种萜类、14 种蒽醌类、15 种苯丙素和18 种其他化合物[5-6],具有抗抑郁、抗菌、抗肿瘤、抗氧化、杀虫、镇静、催眠等多种药理活性[7-8]。
我国虽然是萱草属植物自然分布最广的国家,相关研究却落后于国外。目前国内所使用的园艺品种主要依靠国外引进,具备我国自主知识产权的园艺品种相对较少。本文阐述了国内萱草种质资源概况、国内外的萱草新品种选育研究的历史概况和成果,重点突出了国内新品种的选育成果,同时介绍了萱草新品种选育技术,旨在为萱草新品种选育提供一定的理论参考。
1 萱草种质资源与新品种选育历史概述
1.1 种质资源
Stout[8]于1941年最早开展了萱草属植物的收集与研究工作。萱草属植物资源相关研究在日本、韩国开始于1960年,到1990年取得阶段性进展[1],而我国对萱草属植物的收集最早可见于1974年[9]。
萱草属植物原属于百合科(Liliaceae),但2016年APG Ⅳ分类系统将萱草属植物划分到阿福花科(Asphodelaceae),下设萱草亚科(Hemerocallidoideae)[1]。萱草属植物约有15~19 种[4],《Flora of China》(http://www.iplant.cn/foc)记载为15 种,主要集中在东亚,部分品种延伸到中欧,其中中国分布最广,有11种,分别是黄花菜(H.citrina)、北黄花菜(H.lilioasphodelus)、小黄花菜(H.minor)、萱草(H.fulva)、北萱草(H.esculenta)、大苞萱草(H.midden-dorffii)、多花萱草(H.multiflora)、西南萱草(H.forrestii)、折叶萱草(H.plicata)、小萱草(H.dumortieri)、矮萱草(H.nana),其中多花萱草、西南萱草、折叶萱草和矮萱草为我国的特有种,主要分布于华北、华中、华东地区[10]。此外,刘长命等[11-12]、段九菊等[13]分别对秦岭南麓和山西省野生萱草种质资源展开研究,发现野生萱草具有丰富的遗传多样性和表型多样性。
中国野生种质资源的多样性为新品种选育提供了基础资源。例如,大苞萱草具有早开花特性;重瓣萱草具有重瓣特性;黄花菜具有花果期较长的特性;小黄花菜较具有耐阴、耐旱的特性[14]。上述种均携带优良的园艺性状,且均可用于萱草属植物新品种选育,具有很高的研究价值。
1.2 新品种选育历史概述
相比于国内,国外早在18 世纪中叶就开始了萱草新品种的选育工作,直到19世纪90年代第一个栽培品种Aprioot 的问世,现代意义上新品种选育工作才正式开始[1]。由于该时期杂交育种工作刚刚兴起,进展非常缓慢。直到20 世纪20年代,Stout[15]以我国南京的萱草资源,培育出第一个红色晚花品种,随后育种者选育出了多种色系的萱草,极大丰富了萱草的花色,萱草正式成为一类重要的观赏植物。20 世纪50年代,Griesbach 等[16]通过染色体加倍的方法培育出四倍体萱草品系,多倍体萱草的出现使萱草的花型、花色更加丰富。20世纪70年代新品种金娃娃的问世,使具有连续开花性状的萱草新品种逐渐受到育种者追捧。
我国最早在春秋时期的《诗经·卫风·伯兮》中就有关于萱草的记载,并于唐代正式将其定名为“萱草”。随后萱草逐渐引申有“母爱”、“忘忧”、“宜男”等寓意。明代《本草纲目》中记载了其习性与形态。《群芳谱》根据其花期和形状进行了分类[17]。现代意义上的萱草育种大致可以分为两个阶段,第一阶段主要集中于食用萱草;第二个阶段主要集中于观赏萱草。20 世纪70年代,中国科学院植物研究所选育出60 余个萱草多倍体新品种[9]。此后,萱草育种陷入停滞阶段,直到20世纪90年代,何立珍等[18]用染色体加倍的方式培育出了第一个黄花菜同源四倍体新品种HAC-大花长嘴子花,我国萱草育种进入快速发展时期,但仍处于起步阶段。近年来,上海应用科技大学萱草育种团队在极早花及大花、常绿新品种的选育工作中获得突破,在美国萱草协会登录新品种21个[19]。
2 萱草新品种选育研究进展
2010年以来全球与萱草育种相关的专利约50 项(图1),其中以我国和美国的专利数量最多。近年来,我国专利申请数量稳步上升,已有超越美国的趋势,但我国申请的专利多集中于萱草新品种选育相关的技术与设备[20-21],涉及杂交亲和性[22]、生殖隔离的克服[23]、野生种质资源的保护和利用[24]等,与美国申请的专利相比,选育出的萱草新品种较少[25-26]。可见我国对于萱草属植物的育种工作较为基础,虽取得了一系列的成果,但距离国外还有一定的差距。
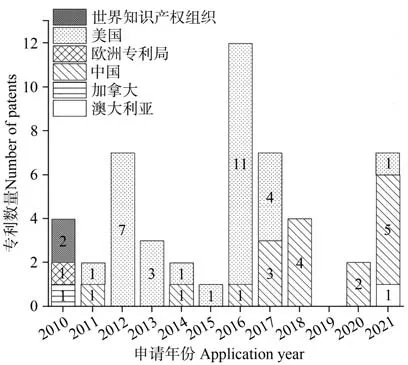
图1 2010年以来萱草新专利申请量分布图Fig.1 Distribution of Hemerocallis patents by country and region in each year since 2010
目前,在美国萱草协会登录的品种已超9 万个[4]。2010年以来,我国在萱草育种工作在花径、花色、花型、花眼、花瓣等方面均取得了不菲的成果,但在花期方面,与园林中广泛应用的金娃娃长达半年的花期相比仍然有一定的差距,在抗性方面,仍未选育出耐极端环境的品种,可见目前萱草的抗病、花香育种仍进展缓慢(表1)。
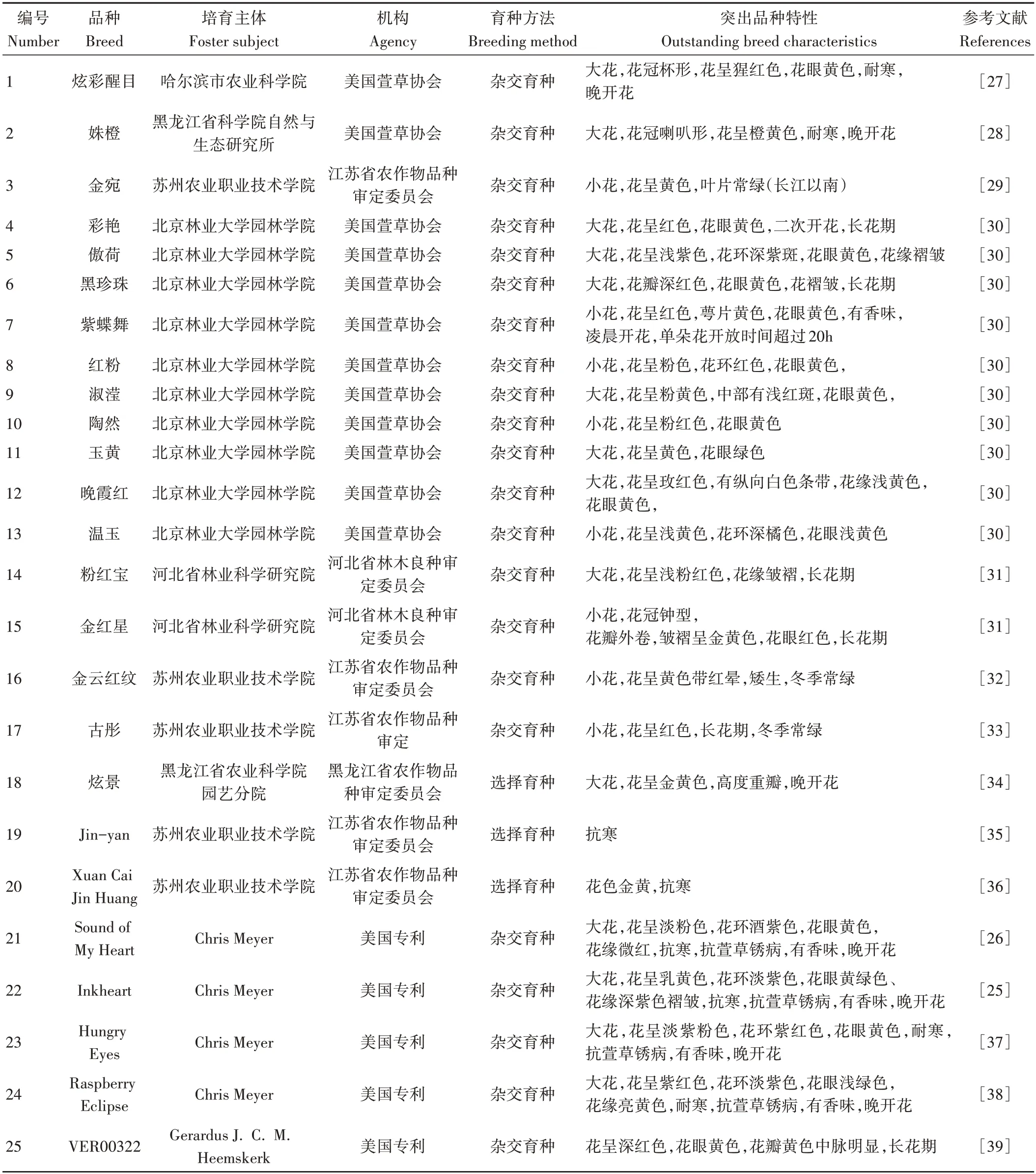
表1 2010年以来国内外萱草新品种选育情况Table 1 Breeding of new varieties of Hemerocallis at home and abroad since 2010
2.1 以花型为选育目标
近年来,花型选育的目标主要是重瓣、异型、蜘蛛型。黑龙江农科院由重瓣野生萱草培育出高度重瓣品种炫景[34],重瓣最高可达7 层。此外,Jamie Gossard 在美国萱草协会(American Hemerocallis Society,AHS)注册了多个花型优良品种,其中White Eyes Pink Dragon获得了2014年AHS的异型萱草品种奖[42]。
2.2 以花香为选育目标
花香是一种优秀的园艺性状,但由于长期被育种者忽视,萱草花香在长期的杂交过程中逐渐丧失。近年来,国内仅有北京林业大学选育出一个具有花香的萱草品种紫蝶舞[30]。国外Chris Meyer 近年来培育的新品种Inkheart[25]、Sound of My Heart[26]、HungryEyes[37]、Raspberry Eclipse[38]均具有花香特征。Keene等[4]研究了147 种萱草的香味特征,发现萱草花香主要来源于挥发性的萜类化合物。
2.3 以花色为选育目标
萱草原生种的花色多以红、橙黄、黄色为主,并不丰富。近年来国内选育的新品种在花色上都较为突出,如红色系品种彩艳、紫色系品种傲荷、粉色系品种红粉、黄色系品种玉黄,其大多数为混合色,带不同颜色花眼、花环、花缘,纯色系仅有黄色的金宛较为突出,红色、紫色、粉色均未见有纯色系品种出现。而国外除了纯白、纯蓝色系品种外,其他色系品种均较为常见。Bahrim 等[43]培育了7 个花色优良新品种,其中米白色的Columna 最为突出;Philip[44-45]培育出鲜红品种R10和深红品种R5。此外,国外已有很多品种花瓣接近纯白色,花眼接近蓝色,例如花瓣为奶白色的Pandora’s Box 和Raspberry Candy[46],花眼为蓝色的Blue Sky Baby、Blast of Blue和Blue Note。
2.4 以花期为选育目标
萱草正常花期在6月中旬到7月下旬,花期短是制约萱草应用的一个重要因素。近年来,萱草在花期育种方面较为突出,主要体现在单花花期、群体花期、早开花和晚开花等方面。紫蝶舞的单朵花期超过20 h[30]。古彤的花期超过120 d,Gerardus[39]培育的VER 品系花期都比较突出,其中VER00322、VER00213、VER00204、VER00198、VER00112、VER00323、VER00157在荷兰栽种的花期均可达到120 d。彩艳具有二次开花特性[30]。Philip[45]培育的WF 品系中WF712、WF73 具有早开花的特性。炫彩醒目[27]、姝橙[28]、炫景[34]晚开花近30 d。
进一步研究单花花期的遗传规律发现,开花性状受一个主基因和多个次要基因调控,白天开花为显性性状,夜间开花为隐性且占优势;花朵闭合的情况更为复杂,可能受多基因调控,推测存在另一个主基因且与控制开花性状的主基因不同,均是核基因控制单花花期[47-48]。不同开花时间的萱草与黄花菜杂交可延长杂交苗的单花花期[49]。Matsuoka 等[50]以不同开花时间的萱草与黄花菜进行了杂交,得到了单花花期达到16 h的杂种苗;Ren 等[51]以同样的方法得到的子代杂种苗单花花期最长达到了23 h。而群体花期与叶数、植株的分支数量有关,叶数增多能为开花提供更多的营养物质,分支数量则为花朵数量的增多提供了条件[52-54]。多位学者研究萱草开花相关基因发现,6 个DSA 家族基因、转化酶抑制蛋白基因、AP1基因、SVP基因、TFL1基因及NAC转录因子均被证明与花瓣衰老相关,为长花期萱草的选育提供了理论依据(表2)。其中HkTFL1可能为连续开花萱草选育的关键基因,但仍需要进一步研究确定其功能。生物节律基因(LHY基因)能控制成花转变,其相似基因在萱草中被发现,可能是晚花和早花品种产生的关键基因,但其功能仍需更进一步研究。
2.5 以抗寒为选育目标
高纬度地区耐寒、低纬度地区常绿新品种的选育是近年来萱草育种的热点。炫彩醒目[27]、姝橙[28]在东北大部份地区可露地越冬,金云红纹[32]、古彤[33]在长江三角洲地区冬季常绿。Chris Meyer培育的几个新品种在美国多个地区具有较好的抗寒性[21-23]。多位学者在研究抗寒相关基因时发现(表2),HfSWEET2a、HfCIN3与HfVIN1的基因功能与低温胁迫相关,miRNA家族、Ca2+信号通路基因、MAPKs信号通路基因、转录因子、热激蛋白基因和热休克蛋白基因在低温胁迫条件下均有候选基因被鉴定,但仍未获得相应的转基因萱草。尤其是其中的HSP90可作为抗低温胁迫候选基因,其功能尚需进一步确定。

表2 萱草属植物部分已克隆的基因Table 2 Parts of cloned genes of Hemerocallis
2.6 以抗病为选育目标
自21 世纪初以来,萱草锈病成为一个世界性的问题,而抗锈病萱草新品种选育成为一个研究热点。Chris Meyer 近年来培育的Sound of My Heart[21]、Inkheart[22]、Hungry Eyes[23]、Raspberry Eclipse[24]均可抗萱草锈病。Philip[45]培育的WF 品系中WF712、WF73、WF7-155 都具有较强的抗锈病能力。有研究表明,Prairie Blue Eyes、Bertie Ferris 为高抗品种[70]。Sandoval 等[71]对比了5 种萱草基因型的抗病性,发现北黄花菜为低抗品种,大花萱草品种Radiant Greetings和Stella de Oro具有中等抗性;萱草抗锈病能力最强。
3 萱草新品种选育方法研究进展
3.1 杂交育种
萱草属植物倾向于异花授粉机制[72],易于杂交,部分自交亲和。常规条件下,萱草属植物的自交亲和性差[73],相同倍性萱草杂交亲和性存在较大差异,结实率相对较高,二倍体间杂交的亲和性整体优于四倍体间杂交[74-75];不同倍性萱草杂交亲和性同样存在较大差异[74],结实率普遍较低,且正反交差异明显[75]。此外,萱草属植物不同品种间的杂交亲和性差异大,且无明显规律[74],因此在杂交试验中筛选出杂交亲和性较强的亲本组合尤为重要。目前已有研究表明,金娃娃是具有优良育性的亲本选择[76]。
花药与柱头的相对位置过大是萱草属植物自交不亲和的主要原因,同时也是其保持进化的重要机制[77]。对于亲和性较高的品种而言,杂交结实率受受精前障碍和受精后障碍共同影响;而对于亲和性较差的品种而言,杂交结实率低由受精前障碍导致[78]。受精前障碍主要包括:1)单朵花期短所导致的花期不育;2)花粉母细胞染色体分裂异常;3)柱头可授性不确定;4)花粉管生长异常。受精后障碍主要是杂种胚与胚乳不亲和导致的双受精比例低及胚发育异常。针对花期不育,通过花粉超低温贮藏来保存花粉活力;粉母细胞染色体分裂异常由遗传因素决定,因此亲本选择尤为重要;而柱头可授性最佳时间同样需要根据亲本来确定,一般在晴天上午9 点进行。蕾期授粉、重复授粉、生长调节剂和食盐涂抹柱头等均可增强花粉和雌蕊的亲和性,进而提高杂交结实率[78]。在胚发育到一定阶段,可通过幼胚离体培养技术克服杂交败育[79-80]。有研究表明授粉后7 天是最佳胚培养时间[81],同时倍性相近的植株成功率更高[82]。
杂交育种是目前萱草属植物育种的主要手段,由于其简单便捷,从最初较为原始的群众性常规杂交到后来多种花粉混合授粉杂交,选育出了几万个观赏新品种。但通过杂交的方式选育新品种时间长、效率低,且对萱草属植物杂交的认识大多停留在表型规律,其深入的机理机制相关报道还较少。
3.2 多倍体育种
萱草属植物常见的基因型有二倍体、三倍体和四倍体。多倍体萱草通常表现出优良的性状[83],例如抗寒能力强[84]、花色丰富[85]。组织培养技术未成熟之前多直接使用秋水仙素处理植株器官的方法获得多倍体萱草[86],现在多采用浸泡法或共培法处理植株愈伤组织获得多倍体萱草[87-88],转化率最高可达50%,秋水仙素的最佳浓度为20 mg·L-1[89]。秋水仙素诱导二倍体萱草产生的新植株大多为同源四倍体,再与其他四倍体植株杂交时亲和性明显提高,甚至可以产生杂交种子。这有效解决了萱草属植物二倍体与四倍体植株杂交亲和性低的问题。
由于秋水仙素有微毒,同时诱导多倍体过程中易形成嵌合体导致加倍失败,恢复为二倍体植株[90],可用安全高效的安定灵、氟乐灵和甲基氨基丙酸磷等有丝分裂抑制剂代替秋水仙素[91-92]。此外,分离嵌合体可使用倍性鉴定技术分离[93]。倍性鉴定方法主要有以细胞学、形态学为主的间接鉴定法和分子水平的直接鉴定法,最常用的是流式细胞测定法和根尖染色体制片法[94-95]。
多倍体常出现新的表型,使物种在极短的时间里产生质的进化。目前多倍体育种大多使用秋水仙素抑制愈伤组织有丝分裂过程,从而获得多倍体植株,但试验预期与结果有时差距较大,其结果具有一定的盲目性,同时同源多倍体的出现也增加了萱草属植物品种间的相似性,后续研究可深入剖析染色体加倍的微观变化,尝试通过胚拯救技术培育出更多异源多倍体植株。
3.3 空间诱变育种
空间辐射诱变技术主要是利用太空特定的环境,引起植物染色体畸变。研究表明,空间辐射诱变的主要诱因是强辐射和微重力。强辐射能够抑制茎尖的生长,打破植株的顶端优势,促进侧芽的生长[96],对细胞分裂产生诱变作用,同时微重力的环境为其提供了充足的时间进行DNA 修复,但其结果并不可控[97-98]。2020年,我国开展了黄花菜的空间辐射育种的研究,选育出了马莲黄花菜新品种金蕾二号,其产量和抗病性明显增强,但是其园艺性状并不突出[99],进一步开展组织培养研究发现花丝更加适宜做外植体[100]。
我国对空间诱变育种的研究总体处于比较初级的阶段,由于其试验周期长,且条件苛刻[101],在萱草属植物中的相关报道不多,且多以使用品种黄花菜为试验材料,观赏品种还鲜见相关报道。
3.4 分子育种
分子育种通过对遗传基因的定向改造,克服了传统育种方法时间长、效率低的缺点。目前,萱草属植物的分子育种进步迅速,尤其是近5年来,分子标记辅助选择技术、转基因技术、基因编辑技术取得了一定的进展。
侯非凡[102]使用表达序列标签微卫星(expressed sequence tag-simple sequence repeats,EST-SSR)标记首次构建了1张黄花菜种内遗传图谱、1张黄花菜与萱草种间遗传图谱。高阳等[103]进一步针对秋水仙碱和昼夜节律这两个性状使用基因组微卫星(simple sequence repeats,SSR)标记对黄花菜与萱草种间遗传图谱进行了加密。张玲玲等[104]使用单核苷酸多态性(single nucleotide polymorphism,SNP)标记定位了5 个花部性状紧密相关的15 个标记,对黄花菜与萱草种间遗传图谱进行了加密。但是,分子标记辅助选择技术在实际新品种选育中应用仍较少。目前,多位学者从萱草中已经克隆了近30 个基因,主要集中在花期、抗逆等方面(表2),但仅有部分基因成功转化到萱草中。倪迪安等[65]通过花粉管道法获得转基因萱草,但是其转化效率仅为0.39%。钟荣[56]同样通过花粉管道法构建了萱草遗传转化体系,但是并未得到转基因植株。刘颖竹[57]构建了基于农杆菌介导的基因编辑遗传转化体系,得到的转基因植株前期出现白化现象,整体矮化、生长慢。
导致分子育种技术应用较少、转化率低的主要原因是萱草属植物的基因组研究相对较少,尤其是国内对于萱草属植物基因组的研究起步较晚。Qing 等[105]报道了黄花菜的全基因组约为3.8 Gb,同时在黄花菜基因组中并未发现秋水仙碱合成途径的相关基因。Lee 等[106]报道了萱草(H.fulve)叶绿体的全基因组,同时发现其与芦荟的分类关系较为接近。Zheng等[107]同样发表了黄花菜(H.citrina)叶绿体的全基因组。Hirota等[108]推测叶绿体捕获导致了叶绿体基因组和核基因组系统发育之间的不一致,绝大多数萱草属植物分类种群之间存在显著的种间基因流动,但是花部性状所对应的核基因组种间基因流动有限,加剧了萱草属植物基因组研究和系统发育研究的复杂性。
4 问题及展望
近年来,萱草属植物育种研究主要集中在以下五个方面:种质资源的收集及遗传多样性分析;杂交亲和性差及其克服措施;育种目标主要集中在花色、花型、花期、抗逆、花香;花期和抗逆基因的克隆及分子标记的开发;遗传转化体系的建立。但是在育种过程出现了很多问题:野生资源收集与保护的力度远远不够;杂交规律缺乏分子水平的阐释;品种间的相似性越来越高;新技术使用少且不成熟;基因组研究相对较少。
未来,萱草属植物研究可以围绕以下几个方面深入开展:1)在野生种质资源利用方面,花色、花期、花香、常绿一直是萱草新品种选育的重要方向,需要特别重视尚未开发的野生种质资源,配制组合,充分利用其杂种优势,选育出适合我国应用的、具有优良性状的萱草新品种;2)在新品种选育方法方面,现有萱草育种方法主要依赖于常规的选择育种、杂交育种,育种周期长,效率低,结合分子育种技术,构建遗传图谱,定位出控制花色、花期、花香等的关键基因,缩短育种周期,提高育种效率,将成为未来萱草育种的必然趋势。
猜你喜欢
今日农业(2022年14期)2022-09-15
园林科技(2021年1期)2022-01-19
红领巾·萌芽(2021年9期)2021-10-09
紫禁城(2020年5期)2021-01-07
人大建设(2019年4期)2019-11-17
世界热带农业信息(2019年9期)2019-01-05
山东农业科学(2017年5期)2017-06-05
上海农业学报(2017年3期)2017-04-10
海峡姐妹(2016年7期)2016-02-27
人间(2015年19期)2016-01-04
