
二穗短柄草BdGI基因表达及其蛋白互作分析
2023-03-09 09:47童伟杨路雪萍罗文举钱佳璇吴佳海王小利
核农学报 2023年4期
童伟杨 路雪萍 罗文举 钱佳璇 吴佳海,3,* 王小利,4,*
(1贵州大学生命科学学院/农业生物工程研究院,山地植物资源保护与种质创新教育部重点实验室/贵州省农业生物工程重点实验室,贵州 贵阳 550025;2贵州大学动物科学学院草业科学系,贵州 贵阳 550025;3贵州省农业科学院畜牧兽医研究所,贵州 贵阳 550005;4贵州省农业科学院草业研究所,贵州 贵阳 550006)
植物的成花转变受到外界环境信号和植物体内分子网络的共同调节,具有多种调控途径,其中光周期途径作为一条重要途径,通过感知外界日照长度调控植物成花[1]。关于拟南芥的研究发现,CO(CONSTANS)是调控成花的关键基因,该基因编码一种锌指转录因子,可以促进FT(FLOWERING LOCUS T)的表达,进而促使植物成花[2]。生物钟是植物感知日照长度变化并依此调节自身节律的内在生理机制,其调控网络由输入途径、输出途径、维持自身节律的中央振荡器三部分组成[3]。生物钟可以依赖外界光信号的输入来实现与外界光环境的同步变化,其中GI(GIGANTEA)是调节昼夜节律的重要基因[4-5]。GI基因位于CO和FT基因的上游,能通过调控CO和FT的表达来调控植物成花,在控制生理节律和调控植物开花的过程中具有重要意义[6-9]。GI突变体在1962年首次被描述为晚花表型[10],Suárez-López 等[11]最先在拟南芥中发现了在长日照下促进植物成花的GI-CO-FT调控路径,GI的表达受到植物生物钟的调控。CCA1(CIRCADIAN CLOCK ASSOCIATED 1)作为生物钟的关键基因,其转录翻译的CCA1 蛋白可以结合到GI的启动子上,从而降低GI的表达[12-13],当CCA1的表达受到抑制时,GI的mRNA在白昼中期积累。ELF3(early flowering 3)最早确定为拟南芥开花的负调控因子,负责将环境的光信号传递入生物钟的中央振荡器中,从而调节下游的各种生理过程[14]。当处于连续光照条件下时,elf3突变体中GI基因转录水平节律消失,表明ELF3蛋白质可能具有调控GI基因的转录节律性的功能[7]。
植物GI基因有调节生物钟与调控开花时间两大功能。但是目前有关GI同源基因的作用机制研究主要集中在常规的模式植物拟南芥[Arabidopsis thaliana(L.)Heynh]上[15],GI在禾本科植物的农学性状如分蘖、穗分化等的作用机制有待进一步研究。二穗短柄草[Brachypodium distachyon(L.) Beauv.]作为一种新型模式植物,其功能基因的挖掘有待完善。为明确GI基因在二穗短柄草昼夜节律调控与光周期开花应答中的分子作用机制,本研究以二穗短柄草为试验材料,利用实时荧光定量PCR(quantitative real-time PCR,qRTPCR)技术检测BdGI在不同日照长度和不同生长发育状态下的表达节律,应用酵母双杂交、双分子荧光互补和免疫共沉淀技术验证BdGI和ZTL 蛋白互作的真实性,以期为进一步探讨BdGI的功能作用提供理论基础。
1 材料与方法
1.1 试验材料
二穗短柄草(Brachypodiam distachyon)Bd21,由四川农业大学馈赠。
主要试剂:Axyprep RNA 提取试剂盒,美国Axygen;GoScriptTM反转录试剂盒,美国Promega。
主要仪器:RXZ型人工智能气候箱,宁波江南仪器厂;实时荧光定量PCR仪,德国Eppeddorf。
1.2 试验方法
1.2.1 二穗短柄草的种植和光照处理 提前将营养土与蛭石按1∶1比例混合装盆并加水浸湿。挑选饱满的Bd21种子均匀撒在花盆中,表面覆少许蛭石,放置于人工智能光照培养箱中,给予不同的光照处理并定期浇水管理。长日照(long day,LD)处理条件:温度22 ℃、相对湿度60%、光照16 h/黑暗8 h、光照强度5 400 lx;短日照(short day,SD)处理条件:温度22 ℃、相对湿度60%、光照8 h/黑暗16 h、光照强度5 400 lx,随后转入全光照/全黑暗处理。
1.2.2 二穗短柄草BdGI基因的表达分析 取不同光照处理下的二穗短柄草叶片,每次取样间隔4 h,连续取样24 h,每次取靠近植株顶端的3 片叶,混合备用。在长日照条件下,对处于不同生长时期的二穗短柄草取样,取样方式同上。取样时间安排如表1。取样后将叶片置于液氮速冻后置于-80 ℃冰箱备用。
表1 取样时间列表Table 1 Sampling time list
使用Axyprep RNA 提取试剂盒提取总RNA,用GoScriptTM反转录试剂盒合成cDNA。cDNA 合成反应结束后稀释4 倍用于qRT-PCR 检测,以泛素结合酶基因(ubiquitin-conjugating enzyme gene)为内参基因(BdUBC18),反应体系共20 μL:GoTaq®qPCR Master Mix 10 μL,10 μmol·L-1引物上下游各0.3 μL,模板cDNA 2 μL,ddH2O 7.4 μL,引物信息见表2。反应程序:96 ℃预变性6 min,96 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,共45个循环。每个样品3次技术重复,3次生物学重复,用2-ΔΔCT方法分析数据。
表2 引物列表Table 2 Primer list
1.2.3 GI 与ZTL 酵母双杂交试验 首先构建GIpGADT7 和ZTL-pGBKT7 载体,随后准备好酵母感受态细胞(LiAC Method),将构建好的载体与酵母感受态细胞共转化,得到酵母菌株Y2H gold 后,以0.9% NaCl悬浮酵母细胞,将菌液涂布于SD/-Leu/-Trp培养基上,待其长出克隆后,将其挑选到SD/-Ade/-His/-Leu/-Trp培养基上,30 ℃培养3~5 d 后,将长出的克隆挑到含有X-α-Gal的四缺培养基上,观察酵母生长情况。
1.2.4 免疫共沉淀检测 采用免疫共沉淀法验证蛋白之间存在相互作用。构建pUC-SPYNE-GI与pSPYCE-35S-ZTL 目的质粒,将目的基因构建至载体上,测序验证,抽提质粒备用。将重组质粒转化到农杆菌感受态细胞GV3101中,并将其制备成浸提液,注射入烟草叶片中。
光照处理48 h 后收集叶片,使用0.5 mL 预冷的RIPA 裂解液(RIPA Lysis Buffer)使细胞裂解提取总蛋白,用磷酸缓冲盐溶液(phosphate buffered saline,PBS)将总蛋白稀释到约1 μg·μL-1,使用Pierce Magnetic HA-Tag IP/Co-IP Kit(货号88838)和Pierce Magnetic MYC-Tag IP/Co-IP Kit(货号88844)试剂盒,按照说明书进行免疫共沉淀试验。
上述步骤洗脱后得到的样品进行非变性聚丙烯酰胺凝胶(Native-PAGE)电泳,使用DYCZ-40D 型迷你转印电泳仪进行转膜(北京六一生物科技有限公司),转膜电流为200 mA,转膜时间为2 h。
上述样品用5%的脱脂奶粉进行封闭。加入1∶1 000稀释的IP专用荧光标记一抗(DyLight 488标记的HA一抗(货号ABIN1113144);Alexa Fluor 647标记的MYC一抗(货号ABIN6915479),于室温下封闭1 h,TBST(TBS+Tween)洗涤3 次,每次5 min。最后用ChemiDoc™XRS+凝胶成像系统(Bio-rad,美国)成像。
1.2.5 双分子荧光互补验证 构建pUC-SPYNE-GI与pSPYCE-35S-ZTL 目的质粒,将其转化到农杆菌感受态GV3101中,在LB培养基中加入100 μg·mL-1利福平(农杆菌株GV3101 携带抗性)、200 μg·mL-1氨苄霉素(载体携带抗性)筛选培养,将携带表达载体的菌株制成侵染液,注入烟草细胞,继续培养72 h 后,激光共聚焦显微镜下观察。
2 结果与分析
2.1 长日照和短日照下BdGI基因的表达变化
由图1-A可知,在短日照(SD)处理下,从ZT0时间点(8:00)开始直至ZT8时间点(16:00),BdGI基因的表达量逐渐增大并在ZT8 达到最大值,在进入黑暗处理后BdGI表达量不断下降,并在ZT24 时间点(次日8:00)的表达量与ZT0近乎一致。由图1-B可知,长日照(LD)处理下,在ZT0~ZT8,BdGI的表达模式与短日照处理下的表达模式相似,并在ZT8 时间点到达峰值,随后BdGI表达量呈现下降趋势,在ZT24 表达量恢复至与ZT0一致。由图1可知,光照会影响到BdGI的表达,不同的光照处理其表达模式不同,但都体现出了BdGI的表达具有一定的昼夜节律性。
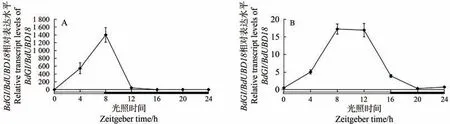
图1 短日照(A)和长日照(B)下BdGI基因的相对表达Fig.1 The relative expression of BdGI gene under short-day (A) and long-day (B)
2.2 长日照和短日照分别转全光照、全黑暗下BdGI的表达变化
24 h 内长日照转全光照[(LD-light-light(LL)]BdGI的表达量变化趋势与长日照(LD)基本相似,BdGI的表达峰值均在ZT8时间点(16:00)时,随后逐渐降低(图2-A);长日照转全黑暗处理[LD-dark-dark(DD)],初始时,BdGI的表达量最高,随着黑暗处理,其表达量开始下降,直至ZT16 时间点(24:00)时表达量几乎检测不到,随后表达量逐渐升高(图2-C);短日照转全光照处理(SD-LL),BdGI的表达量逐渐下降至ZT4时间点(12:00)后逐渐升高,至ZT8 达到最高,随后开始逐渐下降(图2-B);短日照转全黑暗处理(SD-DD),ZT0~ZT12(8:00~20:00)这一阶段BdGI的表达量逐渐升高,即ZT12时,BdGI的表达量达到最高峰,随后从ZT12~ZT20(20:00~次日4:00)这一阶段开始逐渐下降,到ZT20~ZT24(次日4:00~次日8:00)其表达量几乎检测不到(图2-D)。
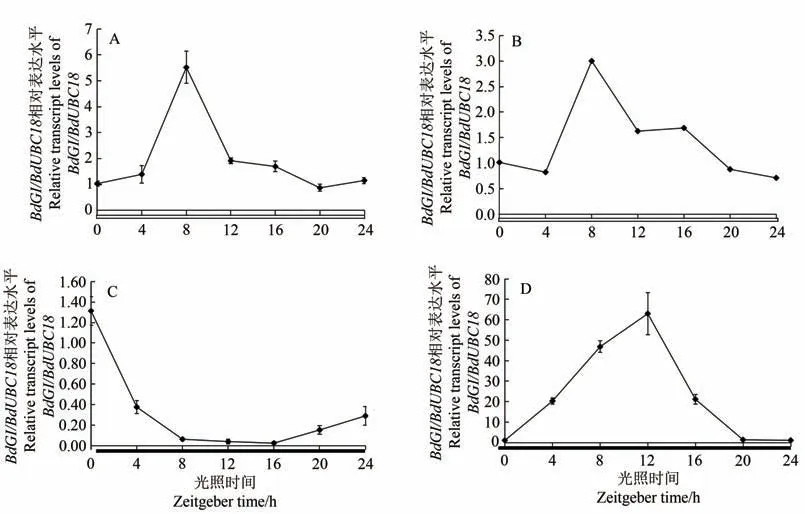
图2 全光照和全黑暗下BdGI基因的相对表达量Fig.2 BdGI gene relative expression under aways light and darkness
2.3 不同发育阶段BdGI的表达变化
取长日照下幼苗期、分蘖期、孕穗期和抽穗期4个时期的二穗短柄草叶片,qRT-PCR结果如图3所示。不同发育阶段BdGI的表达量变化趋势相似,均为先升后降,不同的是不同发育阶段BdGI的表达量峰值所处时间点不同,其中幼苗期与分蘖期均在ZT12 时间点(20:00)表达量到达峰值(图3-A、B);而分抽穗期与在孕穗期ZT8时间点(12:00)表达量到达峰值(图3-C、D)。
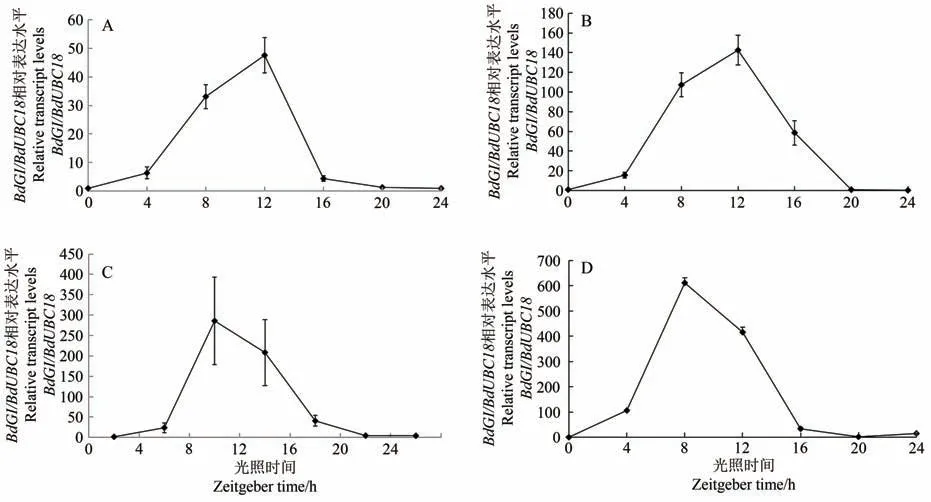
图3 不同发育阶段BdGI基因的相对表达量Fig.3 Relative expression of BdGI gene at different stages of development
2.4 酵母双杂交
酵母双杂交结果如图4 所示,阳性对照BD-P53/AD-T、阴性对照BD-lam/AD-T、自激活BD-ZTL/AD、试验组BD-ZTL/AD-GI 在二缺培养基(-LW:SD/-Leu/-Trp培养基)上都长出了克隆,说明转化成功。将SD/-Leu/-Trp培养基上的克隆挑取至SD/-Ade/-His/-Leu/-Trp平皿和SD/-Ade/-His/-Leu/-Trp(X-α-Gal)培养发现,除了共转化阳性对照质粒的菌株正常长出克隆和变蓝外,试验组BD-ZTL/AD-GI也可正常长出克隆及变蓝,说明BdZTL和BdGI有潜在蛋白互作关系。
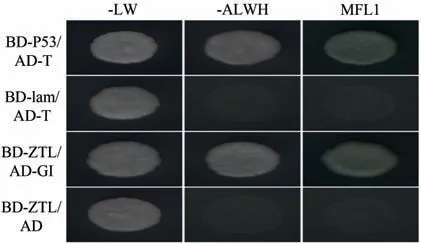
图4 GI与ZTL的双杂交验证结果Fig.4 Yeast two-hybrid validation results of GI and ZTL
2.5 免疫共沉淀
免疫共沉淀(co-immunoprecipitation)是以抗体和抗原之间的专一性作用为基础,研究两种蛋白在细胞内是否存在相互作用的常用方法,若二者有相互作用,则会沉淀出相应的条带。由图5可知,阳性对照IB:HA未能检测到条带,IB:MYC 检测到条带,说明GI-MYC 存在于阳性对照组Input中,表明细胞裂解液中存在目的蛋白,提取蛋白试验无误。Co-IP样品IP-HA两条胶均未检测出条带,说明IB:HA 与IB:MYC 均不能使HA 沉淀。Co-IP 样品IP-MYC 第一条胶未能检测到条带,IB:MYC检测到条带,说明IB:HA不能使GI-MYC沉淀,而IB:MYC能使GI-MYC 沉淀。由图6 可知,相应阳性对照检测到条带,说明ZTL-HA 与GI-MYC 均存在于阳性对照Input 中,证明目的蛋白存在。Co-IP 样品沉淀物用IB:HA 和IB:MYC 均可检测到蛋白条带,根据图5 可知HA 不能使GI-MYC 沉淀,能使ZTL-HA 沉淀,而IB:MYC 能使GI-MYC 沉淀,不能使ZTL-HA 沉淀。所以仅在GI蛋白和ZTL蛋白存在相互作用,且两者带着MYC 与HA 标签才能均被IB:HA与IB:MYC沉淀。利用IgG组进行沉淀试验,GI蛋白和ZTL蛋白未沉淀,排除蛋白与抗体非特异性结合的可能性。综上,BdGI与BdZTL之间存在相互作用。
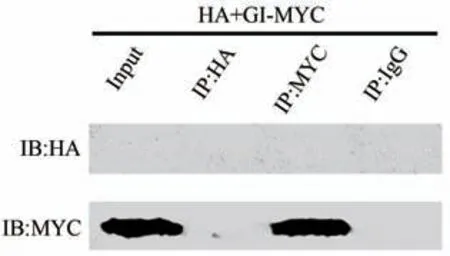
图5 BdGI与BdZTL免疫共沉淀试验结果(对照组)Fig.5 Co-immunoprecipitation results of BdGI and BdZTL(control group)
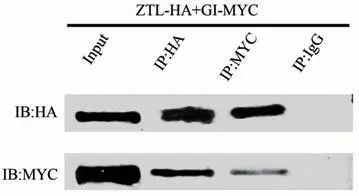
图6 BdGI与BdZTL免疫共沉淀试验结果(试验组)Fig.6 Co-immunoprecipitation results of BdGI and BdZTL(experimental group)
2.6 双分子荧光互补验证
在酵母双杂试验中,试验组在SD/-Ade/-His/-Leu/-Trp 平皿和SD/-Ade/-His/-Leu/-Trp(X-α-Gal)培养基中可以正常克隆且变为蓝色;在免疫共沉淀试验中,只有试验组才能沉淀出蛋白条带,说明BdGI 蛋白与BdZTL蛋白有潜在的互作关系。为了进一步验证其互作关系的真实性,本研究进行了双分子荧光互补试验,结果如图7所示。在激光共聚焦显微镜下观察发现试验组BdZTLnYFP/BdGI-cYFP 在细胞核中有黄色荧光信号;阴性对照BdGI-nYFP/35S-cYFP和35S-nYFP/ BdZTL-cYFP未能检测到黄色荧光信号。以上结果说明,BdZTL 和BdGI 可在细胞核内发生相互作用,且结果与酵母双杂交的结果相符,即BdGI与BdZTL存在相互作用。
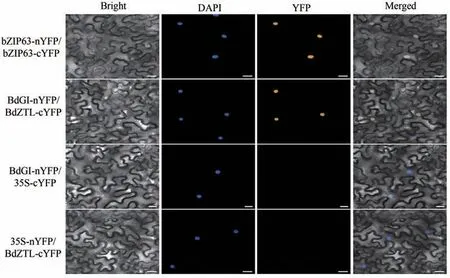
图7 BdGI与BdZTL的双分子荧光互补试验结果Fig.7 Bilolecular fluorescence complementary experimental results of BdGI and BdZTL
3 讨论
基因在生物体内表达受一定时间与空间的影响,从而表达出特异性,不同的表达模式显示出不同的功能。前人研究表明,在拟南芥中,GI能够调控植物开花,同时也参与调节昼夜节律[16],且在不同日照条件下,拟南芥GI表达量存在差异。本研究利用qRT-PCR检测BdGI不同光照处理下的表达模式,发现24 h 内在开灯后即能检测到BdGI的表达,BdGI在LD 和SD 下的表达峰值均出现在ZT8,即下午4∶00 左右,表明BdGI的表达具有一定的昼夜节律性,并且与拟南芥在长日照处理下相比,BdGI表达峰值出现的时间有所提前。值得注意的是,BdGI在不同光照条件下表达量的变化速率也存在不同,当处于长日照条件时,BdGI表达量在到达峰值后下降速度较慢,而短日照下降速度较快,说明BdGI受光照的影响且具有一定的自我调节能力以应对光照的变化。这一结果与小麦中的GI基因表达模式基本相似[17],仅在不同的植物内表达峰值出现的时间不同。采用连续光照的方式处理植株后,BdGI表达高峰出现的时间未发生改变,而长日照与短日照转全黑暗处理下BdGI的表达模式呈现不同的趋势。从长日照转全黑暗处理结果可以看出,长日照下BdGI表达量积累较多,短日照下BdGI表达量积累较少,而在黑暗时期BdGI表达量逐渐增加到达峰值,随后降低,表现出BdGI的表达受光周期调控,这与大豆[Glycine max(Linn.)Merr.]CmGI在光照转黑暗下的表达模式相似[18]。本研究还发现,不同发育阶段BdGI的表达量变化趋势大致相同,仅不同发育阶段下BdGI的表达量峰值所处时间点不同,其中幼苗期与分蘖期均在ZT12点表达量达到峰值,而抽穗期与在孕穗期在ZT8 点表达量达到峰值。值得注意的是,抽穗期的表达峰值最高,由此猜测二穗短柄草BdGI在抽穗期能发挥重要的作用。
GI 的表达受到生物钟的调控,生物钟的形成过程中有多种蛋白参与,通过转录和翻译后的调控作用,形成复杂的负反馈调节系统[19],其中CCA1、LHY(LATE ELONGATED HYPOCOTYL)和TOC1(TIMING OF CAB EXPRESSION 1)被认为是植物生物钟振荡器的核心元件,其调控方式为:早晨元件LHY和CCA1抑制TOC1的转录,晚间元件TOC1下调LHY/CCA1的积累[20-22]。ZTL属于F-box家族蛋白,其家族成员还包括FKF1(Flavin-binding,Kelch repeat,F-box1)、LKP2(LOV kelch protein2)[23-27],且都具有LOV结构域、F-box结构域、Kelch重复序列3个结构域[28],具有区分日照长度和控制光信号输入的能力[20,29]。ZTL mRNA 的积累不受生物钟的调控[23],但是ZTL 蛋白水平呈现出周期性节律变化,并在夜晚时其蛋白含量达到峰值[25],这与GI表达的昼夜节律高度同步,因此推测GI 蛋白与ZTL 蛋白存在某种相互作用,使得其表达模式基本一致。蓝光条件下,拟南芥中ZTL蛋白会与GI蛋白相互作用,起到稳定ZTL 蛋白的作用[24],同时解除ZTL 对TOC1 的抑制作用。傍晚过后LOV 功能域对蓝光的应答消失,ZTL 蛋白从GI 解离,靶向介导TOC1 的降解[24-27]。本研究通过酵母双杂、双分子荧光互补、免疫共沉淀等方式证实了二穗短柄草中BdGI蛋白与BdZTL 蛋白也存在相互作用,与拟南芥中的研究结果相符。对模式植物拟南芥研究发现,长日照条件下FKF1 蛋白和GI 蛋白的表达受生物钟的调控,当FKF1 响应蓝光应答,与GI蛋白发生相互作用时,光诱导产生的FKF1-GI 蛋白复合体会在长日照的下午积累到高水平,降解CO启动子上的CDF1 蛋白,解除对CO的抑制作用[30-33]。CO的表达上调确保了其靶标基因FT的表达;FT 蛋白可移动到茎的分生组织中,然后启动开花转变转录调控级联控制[34-36],进而促使植物成花。故推测二穗短柄草中BdGI 蛋白可能与BdZTL 共同调控TOC1表达,调控方式与FKF1/GI蛋白复合体的调控方式相似。
本研究利用酵母双杂筛选检测到BdGI 可能与BdZTL 有潜在的互作关系,由于酵母双杂交试验并非对所有蛋白都适用,而且出现假阳性可能性较大,所以再次利用双分子荧光互补方法与免疫共沉淀法检测进行验证,结果显示BdGI与BdZTL 存在互作关系。但对于BdGI 在光周期调控开花及稳定昼夜节律过程中行使何种职能;BdGI 与BdZTL 的相互作用如何调控下游基因的表达仍需进一步探索。
4 结论
本次研究利用qRT-PCR 技术分析了BdGI在不同光照处理下和长日照的不同发育阶段中的表达情况,结果表明BdGI的表达具有一定的昼夜节律并受光周期的调控;通过酵母双杂筛选出BdGI 的互作蛋白BdZTL,并通过双分子荧光互补与免疫共沉淀验证了BdGI与BdZTL互作的真实性。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
儿童时代·幸福宝宝(2021年1期)2021-03-29
小资CHIC!ELEGANCE(2019年40期)2019-12-10
音乐教育与创作(2019年8期)2019-05-16
星星·散文诗(2017年2期)2017-07-05
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年5期)2016-08-20
