
2015—2021年江苏省粳稻新品种(系)对水稻条纹叶枯病的抗性评价
2023-02-26 05:51兰莹杜琳琳林峰李晨羊周益军宋锦花许明周彤
江苏农业科学 2023年24期
关键词:粳稻
兰莹 杜琳琳 林峰 李晨羊 周益军 宋锦花 许明 周彤
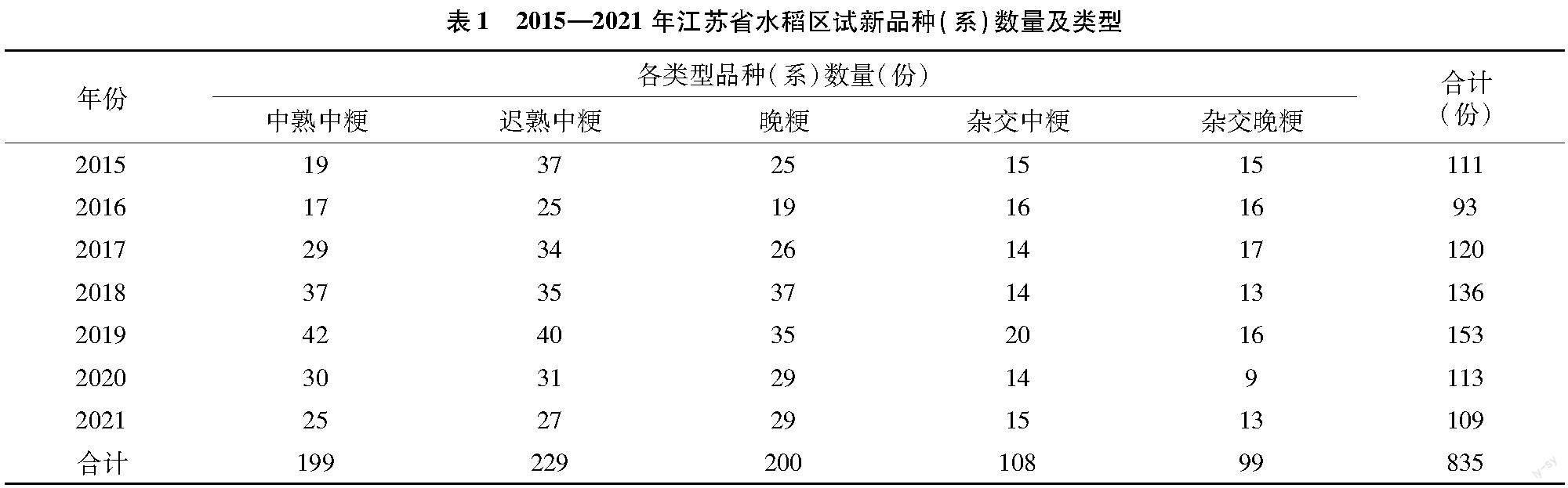


摘要:对2015—2021年度835份江苏省粳稻新品种 (系 )通过人工接种法进行水稻条纹叶枯病抗性水平鉴定评价,以期为生产上水稻品种布局和病害综合防控提供参考依据。统计结果表明,86%~100%的区试品种达到中感以上的抗性水平,高抗品种的比例从0(2016年)渐增到12.39%(2020年)和11.93%(2021年),抗病品种的比例在17.65%~48.62%,中感品种在39.45%~82.35%,2015年和2016年分别检测到1个和3个感病品种,2016年和2017年分别检测到10个和6个高感品种,2018年以后未发现感病和高感品种,说明经多年品种更替,江苏省区域试验粳稻品种对条纹叶枯病的抗性水平已经有了较大提升。2015—2021年抗病及高抗品种在各区组所占比例从高到低依次为中熟中粳、早熟晚粳、迟熟中粳、杂交晚粳和杂交中粳组,常规粳稻组如中熟中粳和早熟晚粳组,中感水平材料的数量仍较多,在病害暴发年份易成为生产上加重流行的因素。文中列出了筛选出的26份高抗品种,可作为抗性育种资源在生产上应用。
关键词:粳稻;水稻条纹叶枯病;人工接种鉴定;抗性评价;育种价值
中图分类号:S435.111.4+9 文献标志码:A
文章编号:1002-1302(2023)24-0100-05
水稻条纹叶枯病(rice stripe disease)是由水稻条纹病毒(rice stripe virus,RSV)引起的病毒病,传播介体为灰飞虱(small brown planthopper,SBPH)。发病株叶片有褪绿的条纹斑点,首先在幼嫩心叶引发枯死,使植株不抽穗或穗小,造成水稻严重减产。1897年该病最早在日本的栃木、群马等县被发现,之后在朝鲜、我国和乌克兰均有发生[1]。我国的苏南稻区在1963年发现该病害后,20世纪70年代和80年代又在北京、山东、云南等地区多次暴发性流行[2-3]。21世纪以来,水稻条纹叶枯病在江苏省为主的稻麦轮作区大规模流行,逐渐上升为区域性最重要病害,累计发生面积超过1 400万hm2,仅2004年就达到157 hm2,占当年全省水稻种植面积的79%,近3.3万hm2绝收,严重影响了农业生产安全[4]。
水稻条纹叶枯病毒在介体灰飞虱上传毒效率非常高,一经获毒就终身带毒,并经卵传给后代,是持久性传播的病毒[5]。江苏省自20世纪90年代起不断推广粳稻的种植面积,当时省内大部分主栽粳稻品种感病或高感水稻条纹叶枯病,容易造成毒源积累和病害扩散,对抗性品种进行选育和推广,是最为经济环保的防治措施[6]。
研究团队自2002年起连续承担江苏省水稻区域试验品种项目,对省内主栽和新育成品种进行水稻条纹叶枯病的抗性评价和筛选,鉴定出镇稻88、盐稻8号、徐稻3号、连粳4号、宁粳1号、南粳44等一系列拥有稳定抗性的主栽粳稻品种,在江苏、浙北生产上广泛应用[7]。然而近年来,水稻条纹叶枯病在江苏省粳稻种植区发生度逐渐趋轻,主栽品种也已发生较大的更替,对于新品种(系)的抗性水平呈怎样的分布趋势还未见报道。因此,本研究通过鉴定和评价2015—2021年江苏省区试粳稻品种对水稻条纹叶枯病的抗性水平,以期为生产上水稻品种布局和病害综合防控提供参考价值。
1 材料与方法
1.1 品种
参试粳稻新品种(系)共835份,均来自江苏省种子站,其類型及年度间分布见表1。
1.2 接种鉴定方法
2015—2021年人工鉴定水稻品种试验均在江苏省农业科学院病毒鉴定圃(118°52′E,32°02′N)完成。按照《水稻品种抗条纹叶枯病鉴定技术规范》(NY/T 2055—2011)将参鉴品种经浸种、催芽后,选取35粒发芽良好的种子均匀播于盛有自然肥力土壤的透明塑料杯(内径95 mm)中。用斑点免疫结合试验(DIBA)法筛选出带毒率≥80%的灰飞虱群体,待苗长至1.5叶龄期,选取处于2~4龄期的群体用于接种。人工接种鉴定的有效接种虫量以2头/苗为计算标准,用IVS表示,按以下公式计算。
IVS=N×PVS/nt。(1)
式中:N为接种灰飞虱数量;PVS为灰飞虱带毒率;nt为苗数。
根据公式(1)计算每杯接种灰飞虱数量,于 26~28 ℃条件下接入玻璃杯中,上午下午各赶虫1次,确保稻株均匀获毒,接种时间48 h后移走灰飞虱,并将秧苗移栽至15~30 ℃条件下培养。接种15 d后开始调查,每5 d调查1次,连续调查3次。
1.3 病害调查与评价
水稻条纹叶枯病的病级分级标准及症状详见表2。其中:2级以上直接记为病株;1级在 7 d 后再次调查,若表现出2级或更严重症状,则记为病株;0级和1级记为不发病。
各级别的水稻条纹叶枯病抗性分级根据发病率进行抗性评价,发病率计算公式为:发病率=病株数/调查总株数×100%。抗性等级标准详见表3。
2 结果与分析
2.1 江苏省不同年度水稻品种(系)对水稻条纹叶枯病的抗性分析
2015—2021年区试品种发病率的分布情况(图1、表4)显示,绝大部分品种的抗性水平都处于抗病和中感之间,高抗或高感的品种较少。2015—2021年,没有发现免疫品种;高抗品种的占比分别为9.91%、0、5.00%、0、1.96%、12.39%、11.93%;抗病品种的占比从2015年的41.44%降至2018年的17.65%,随后逐年递增,在2021年占比最高,达到48.62%;除2021年以外,其余年份占比最高的是中感品种,在46.90%~82.35%之间。感病品种在2015年和2016年分别检测到1个和3个,高感品种在2016年和2017年分别检测到10个和6个,2018年以后未发现感病和高感的品种(表4)。
2.2 江苏省不同类型粳稻品种(系)对水稻条纹叶枯病的抗性分析
对不同类型水稻品种(系)进行抗性综合指数评价分析,结果显示,2015—2021年, 抗病以上品种占比从高到低依次为中熟中粳组(62.0%)、早熟晚粳组(61.9%)、迟熟中粳组(61.6%)、杂交晚粳组(52.4%)和杂交中粳组(50.0%)。中感以上品种占比从高到低依次为早熟晚粳组(97.5%)、杂交中粳组(97.4%)、迟熟中粳组(96.4%)、中熟中粳组(95.9%)和杂交晚粳组(93.2%)(图2、表5)。虽然抗性以上品种比例基本占50%以上,一些常规粳稻组如中熟中粳和早熟晚粳组,中感水平材料的数量仍较多。
2.3 达到高抗水稻条纹叶枯病的江苏省主栽粳稻品种(系)
由表6可知,2020—2021年共鉴定出26份发病率低于5%的江苏省主栽水稻品种。其中:常规粳稻有W047、武粳8367、泗稻17-543、苏垦93002等16份材料,分属晚粳早熟、迟熟中粳、早熟晚粳、中熟中粳等区组。杂交稻有常优18-7、甬优1540、甬优1836等10份材料,分属杂交晚粳、杂交中粳2个区组。这些达到高抗水平的材料适合应用于生产并作为抗性育种资源。
3 讨论与结论
水稻条纹叶枯病20世纪90年代在江苏及周边省份大范围暴发时,国内许多研究团队对国内的主栽品种进行田间抗性鉴定,其结果显示水稻品种多数不抗水稻条纹叶枯病,且粳稻比杂交稻和籼稻更加感病[8]。本研究结果表明,经过多年主栽品种的更替,近年来江苏省区试粳稻品种对条纹叶枯病的抗性水平已经有了较大的提升。在不同区组中,高抗和抗性品种的比例也在逐步增加。2015—2021年,每年86%~100%的品种可以通过区试审定要求,达到中感以上的抗性水平。其中,2015年、2020年和2021年,有超过一半的品种达到抗病(R)水平(表4)。抗性品种的推广和种植减少了毒源积累和介体传播病毒的概率,降低了田间病毒病的发生。
对水稻抗条纹叶枯病抗源的筛选与鉴定试验始于1965年,Yamaguchi等研究发现,籼稻和爪哇稻品种中存在着丰富的抗性资源,而粳稻较感病[9]。1967年Washio等采用室内苗期接种鉴定了398份品种,发现大部分粳稻品种感病,多数籼稻、爪哇稻和粳型陆稻具有抗性,并筛出了Modan、Tadukan等抗性优良的品种[10]。目前水稻品种对条纹叶枯病的抗性鉴定方法分为田间鉴定和人工鉴定,周彤等引入了有效接种虫量的概念,即通过测算实际带病毒的灰飞虱数量作为接种强度的均一化指标,同时将感病对照和抗病对照的抗性表现也作为量化标准,并结合田间鉴定后的测产试验确定了品种的抗性分级标准(表2、表3)[6]。同时在重病区采用多年多点试验对数十份主栽粳稻品种进行了田间鉴定,结果显示供试粳稻品种在不同年份和不同地区间抗性水平存在一定的差异,其原因可能受到侵染时间、虫量、带毒率、栽培、气候等一系列因素的影响,而用苗期人工接种法鉴定出的水稻抗性品种,在多年大田生产中同样表现稳定的抗性水平[6-7,11]。近年来随着灰飞虱发生量和带毒率逐年递减,一些重病田已经不具备作为田间抗性鉴定的条件,因此自2015年起品种区域试验以人工接种结果作为参考依据。
为了能有效指导抗水稻条纹叶枯病的品种选育,研究者同时开展了大量抗性遗传研究。迄今为止,已报道了5个主要的水稻条纹叶枯病抗性QTL(Stv-bi、qSTV11IR24、qSTV11TQ、qSTV11KAS和qSTV11SG),均位于11號染色体上[12-16]。Wang等在2014年克隆了第1个水稻条纹叶枯病抗性基因STV11,可以编码具有磺基转移酶活性的蛋白,参与水杨酸介导的抗病毒途径,通过对214份不同地区年代的水稻品种序列分析发现,STV11存在明显的籼、粳分化,大部分籼稻中含有抗性基因STV11-R,而粳稻中一般都是感病基因位点STV11-S,这一发现为培育抗病粳稻提供了有用的基因资源[17]。Hu等研究发现,STV11编码的一个磺基转移酶能够将水杨酸(SA)磺化形成磺基水杨酸(SAA),通过上调植株体内SA的生物水平进而抑制病毒的复制[18]。Hayano-Saito等克隆了Stvb-i并发现其表达能够调控热激蛋白Hsp70参与的热响应调控,该途径与病毒的复制相关,Stvb-i可以减少病毒增殖,提升水稻持久抗病性[19]。利用上述抗性基因,育种家已经陆续培育出宁粳1号、连粳4号、徐稻3号、徐稻4号和盐稻8号等抗病优良的材料,为后续抗性品种的创制和选育打造了良好的基础。
抗病品种的大面积种植和绿色防控技术的推广使水稻条纹叶枯病的危害程度逐年减轻,因病害造成的损失被有效控制,但每年依旧有零星的病田分布。在本研究结果中,中熟中粳和早熟晚粳区组有大量中感水平的材料。在江苏省的丘陵稻作区、太湖稻作区、沿江沿海稻作区、淮北稻作区等多个稻作区内,中熟中粳和早熟晚粳是主要种植品种[20]。结合近年来对江苏省田间灰飞虱群体的调查,以上多个地区仍存在较大的虫口基数,一旦病毒病暴发,中感品种易成为生产上加重病害流行的因素。同时,生产上对条纹叶枯病抗性基因的应用目前还比较单一,需要不断挖掘新基因和新品种的抗病优势,培育优质的抗性材料,为保障粮食安全发挥重要作用。
参考文献:
[1]朱凤美,肖庆璞,王法明,等. 江南稻区新发生的几种稻病[J]. 植物保护,1964,2(3):100-102.
[2]林奇英,谢联辉,周仲驹,等. 水稻条纹叶枯病的研究 Ⅱ.病害的分布和损失[J]. 福建农学院学报,1990,19(4):421-425.
[3]陈声祥. 水稻病毒病发生现状及研究进展[J]. 浙江农业科学,1996,37(1):41-42.
[4]程兆榜,杨荣明,周益军,等. 江苏稻区水稻条纹叶枯病发生新规律[J]. 江苏农业科学,2002(1):39-41.
[5]陈 洁,吴丽娟,周 彤,等. 江苏省主栽水稻品种对条纹叶枯病与灰飞虱的抗性评价[J]. 南京农业大学学报,2010,33(4):105-108.
[6]周 彤,范永坚,程兆榜,等. 水稻抗条纹叶枯病鉴定方法的研究[J]. 植物保护,2008,34(6):77-80.
[7]周 彤,周益军,程兆榜,等. 粳稻品种对水稻条纹叶枯病的抗性鉴定及抗病品种镇稻88的遗传分析[J]. 植物保护学报,2007,34(5):475-479.
[8]杨荣明,刁春友,朱叶芹.江苏省水稻条纹叶枯病上升原因及防治对策[J]. 植保技术与推广,2002,22(3):9-10,12.
[9]Washio O,Ezuka A,Sakurai Y,et al. Studies on the breeding of rice varieties resistant to stripe disease:Ⅰ. Varietal difference in resistance to stripe disease[J]. Japanese Journal of Breeding Science,1967,17(2):91-98.
[10]Washio O,Toriyama K,Ezuka A,et al. Studies on the breeding of rice varieties resistant to stripe disease:Ⅱ. Genetic study on resistance to stripe disease in Japanese upland rice[J]. Japanese Journal of Breeding,1968,18(2):96-101.
[11]张梦龙,岳红亮,程新杰,等. 水稻条纹叶枯病抗性机制研究进展[J]. 江苏农业学报,2021,37(6):1608-1613.
[12]Hayano-Saito Y,Saito K,Nakamura S,et al. Fine physical mapping of the rice stripe resistance gene locus,Stvb-I[J]. Theoretical and Applied Genetics,2000,101(1):59-63.
[13]Wu X J,Zuo S M,Chen Z X,et al. Fine mapping of qSTV11TQ,a major gene conferring resistance to rice stripe disease[J]. Theoretical and Applied Genetics,2011,122(5):915-923.
[14]Wang B X,Jiang L,Zhang Y X,et al. Genetic dissection of the resistance to rice stripe virus present in the indica rice cultivar ‘IR24[J]. Genome,2011,54(8):611-619.
[15]Zhang Y X,Wang Q,Jiang L,et al. Fine mapping of qSTV11KAS,a major QTL for rice stripe disease resistance[J]. Theoretical and Applied Genetics,2011,122(8):1591-1604.
[16]Kwon T,Lee J H,Park S K,et al. Fine mapping and identification of candidate rice genes associated with qSTV11SG,a major QTL for rice stripe disease resistance[J]. Theoretical and Applied Genetics,2012,125(5):1033-1046.
[17]Wang Q,Liu Y Q,He J,et al. STV11 encodes a sulphotransferase and confers durable resistance to rice stripe virus[J]. Nature Communications,2014,5:4768.
[18]Hu J L,Huang J,Xu H S,et al. Rice stripe virus suppresses jasmonic acid-mediated resistance by hijacking brassinosteroid signaling pathway in rice[J]. PLoS Pathogens,2020,16(8):e1008801.
[19]Hayano-Saito Y,Hayashi K. Stvb-i,a rice gene conferring durable resistance to rice stripe virus,protects plant growth from heat stress[J]. Frontiers in Plant Science,2020,11:519.
[20]楊立炯,崔继林,汤玉庚. 江苏稻作科学[M]. 南京:江苏科学技术出版社,1990.
猜你喜欢
今日农业(2021年15期)2021-10-14
农民致富之友(2020年32期)2020-12-03
云南农业(2019年3期)2019-02-24
中国稻米(2018年5期)2018-01-21
河南农业(2017年7期)2017-07-25
农村百事通(2017年8期)2017-04-26
上海农业学报(2017年3期)2017-04-10
西南农业学报(2016年4期)2016-05-17
浙江农业科学(2016年11期)2016-05-04
湖南农业(2014年11期)2014-04-01
