
单核细胞增生李斯特菌新型转录调节因子lmo2486的分子特征及原核表达研究
2023-02-16 02:28朱孝珍刘昱诚张星星王立霞季春辉才学鹏孟庆玲
动物医学进展 2023年1期
朱孝珍,刘昱诚,张星星,王立霞,季春辉,郭 蕴,才学鹏,孟庆玲,乔 军*
(1.石河子大学动物科技学院,新疆石河子 832003; 2 .新疆农垦科学院畜牧兽医研究所,新疆石河子 832003;3.中国农业科学院兰州兽医研究所,甘肃兰州 730046)
单核细胞增生李斯特菌(Listeriamonocytogenes,LM)是一种兼性胞内寄生菌,该菌可引发人和多种动物的李斯特菌病,尽管李斯特菌病病例较少,但其病死率很高,被认为是引起公共卫生问题的主要病原菌[1]。成年人的常见症状为头痛、胃痛、发热、腹泻和呕吐,但在高危人群,比如孕妇、婴儿、老人和艾滋病、癌症患者中可能会出现严重症状,如脑膜炎、败血症、流产甚至新生儿死亡[1-4],对于人致死率高达20%~30%[5]。在土壤、动物饲料、水以及哺乳动物和鸟类的粪便中都能发现单核细胞增生性李斯特菌,该菌能通过污染各种肉类和蔬菜、乳制品和即食食品从而导致李斯特菌病暴发[1]。此外,该菌可在高盐、低温、酸碱、高压和其他极端条件下存活[6-7],有较强的环境适应性[8-9]。
LM感染、胞内生存和毒力受多种分子的调控,这些分子组成复杂的调控网络。现有研究发现,含有PspC结构域(噬菌体休克蛋白C)蛋白家族,广泛存在于多种病原微生物中。PspC蛋白是一种重要的转录调节因子,可参与细菌的应激反应系统及毒力相关基因的表达调控,可能与应激反应以及维持细菌细胞包膜的完整性密切相关[10]。然而,目前有关LM PspC家族成员的转录调节蛋白Lmo2486的生物学功能尚不明确。前期的生物学信息分析表明,Lmo2486蛋白具有PspC结构域。PspC结构域被认为是应激反应转录调节因子,可能与LM多个基因的表达调控密切相关[11-12]。同时,Lmo2486蛋白含有DUF4097结构域,该结构域为细菌黏附的可能结构元件。然而,lmo2486具体的生物学功能尚未深入研究。
为了进一步探究lmo2486基因的生物学功能功能,本研究对LM-SB5绵羊野毒株进行lmo2486基因克隆,对编码蛋白的结构进行分子特征分析,并进行了其遗传进化特征和表达研究,为揭示lmo2486在LM中对环境耐受和毒力的调控作用提供前期的研究基础。
1 材料与方法
1.1 材料
1.1.1 菌株、培养基及试剂 LM-SB5新疆野毒株,由石河子大学预防兽医学实验室从发病绵羊体内分离,鉴定后保存。大肠埃希氏菌(E.coli) 感受态细胞DH5α,由石河子大学预防兽医学实验室保存。
1.1.2 培养基及试剂 脑心浸液培养基(BHI),青岛高科技园海博生物技术有限公司产品;DNA标准DL 2 000、pMD19-T(simple)载体,TaKaRa公司产品;细菌DNA提取试剂盒、质粒小提试剂盒、琼脂糖凝胶DNA回收试剂盒,诺维赞生物公司产品;T4 DNA连接酶、限制性核酸内切酶HindⅢ、BamHⅠ、蛋白Marker,TaKaRa公司产品;IPTG,索莱宝公司产品;Trans8K DNA Marker、Transetta(DE3),北京全式金生物技术有限公司产品。
1.1.3 仪器设备 超净操作台,BOXUN公司产品;恒温水浴锅(XMTD-8222)、隔水式恒温培养箱(DNP-9082),上海精宏实验设备有限公司产品;移液器(2.5 μL、10 μL、20 μL、100 μL、200 μL、1 000 μL、5 L),Eppendorf公司产品;高压蒸汽灭菌锅(HVE-50),Hirayama公司产品;普通恒温培养箱(8000DH)、全自动酶标仪(Muitiskan GO)、高速冷冻离心机(Multifuge X1R),Thermo Fisher公司产品;PCR仪、凝胶成像仪、转膜仪,Bio-Rad公司产品;电子天平(TP-213),丹佛仪器(北京)有限公司产品;电泳槽(Power Wave XS2),北京君意东方电泳设备有限公司产品;电泳仪(DYY-2C)、SDS-PAGE凝胶电泳仪(DYY-6C),北京六一生物科技有限公司产品。
1.2 方法
1.2.1 引物设计与合成 根据GenBank登录的lmo2486基因序列(登录号:CP023861.1),设计扩增lmo2486基因特异性引物,上游引物lmo2486F:5′-ACAGACACCGGCAATAT-3′,下游引物lmo2486R:5′-CCGAATGCAGCTGATTCT-3′。由DNA Start软件分析的蛋白抗原表位集中区35~390,设计扩增lmo2486抗原集中区的特异性引物,32a1:CGGATCCATTTATATTATCATCACCAT,32a2:CCAAGCTTAATTCCATTTCCTACTTT,在引物32a1/2的5′端分别添加BamHⅠ和HindⅢ保护性碱基及限制性酶切位点(斜体及下划线部分)。送往北京华大基因公司合成。
1.2.2 LMlmo2486基因的克隆及测序 挑取LM-SB5单菌落于BHI培养基中,置于37 ℃振荡培养14 h~16 h,按照细菌基因组DNA提取试剂盒的说明书提取LM-SB5分离株基因DNA,以该DNA为模板,以设计的lmo2486基因特异性上、下游引物lmo2486F/R扩增lmo2486基因,PCR的反应体系为:PCR Mixture 10 μL,H2O 7 μL,上、下游引物各0.5 μL,DNA模板2 μL。 反应条件为:95 ℃预变性5 min;94 ℃ 30 s,52 ℃ 30 s,72 ℃ 1 min 30 s,72 ℃ 10 min,共30个循环;4 ℃保存。将PCR产物用10 g/L琼脂糖凝胶在0.5×TBE电泳缓冲液中进行电泳,与DNA标准DL 2 000相比,将目的片段用琼脂糖凝胶DNA回收试剂盒回收纯化。回收后的目的片段lmo2486基因与pMD19-T(simple)载体4 ℃过夜连接,然后转化到E.coliDH5α感受态细胞内,通过菌液PCR筛选阳性转化子,将阳性克隆送至华大基因生物公司测序。
1.2.3lmo2486基因编码蛋白质的生物信息学分析 通过Motif Scan在线软件分析糖基化位点、磷酸化位点等。利用在线软件ProtParam预测Lmo2486蛋白的理论分子质量及等电点;通过Protscale分析蛋白质的亲水性、疏水性;通过SignalP进行蛋白质的信号肽预测;利用软件SOPMA预测Lmo2486蛋白的二级结构;应用SWISS-MODEL预测蛋白质的三级结构;使用DNA Star预测蛋白的抗原表位集中区域;利用SMART预测并分析蛋白的结构域。
1.2.4lmo2486基因核苷酸序列遗传进化分析 将LM-SB5lmo2486基因与GenBank中公布的LM不同的菌株和PspC家族和DUF4097家族其他种属细菌的基因核苷酸序列进行同源对比,应用MEGA5.0软件对lmo2486基因核苷酸序列进行遗传进化分析,并采用邻接法(Neighbor-Joining,NJ)构建系统进化树。
1.2.5 重组载体pET-32a(+)-lmo2486K构建 用引物32a1/2扩增lmo2486基因抗原表位集中区的片段并命名为lmo2486K,构建pMD19-T-lmo2486K,转化至DH5α中,筛选阳性克隆,用BamHⅠ和HindⅢ内切酶同时双酶切pMD19-T-lmo2486K重组质粒和pET-32a(+)载体质粒并将其回收,然后使用T4连接酶将pMD19-T-lmo2486K重组质粒和pET-32a(+)载体4 ℃过夜连接,转化至DE3中,进行菌液PCR和双酶切鉴定,将验证正确的质粒命名为pET-32a(+)-lmo2486K。
1.2.6 Lmo2486K的诱导表达及重组蛋白鉴定 将pET-32a(+)-lmo2486K重组质粒的转入DE3感受态细胞内,使用IPTG诱导蛋白表达,然后用120 g/L的SDS-PAGE验证蛋白的表达情况。同时,以小鼠抗His单克隆抗体为一抗,带有His标签的山羊抗鼠IgG为二抗,进行Western blot分析,鉴定表达的重组蛋白。
2 结果
2.1 lmo2486基因的扩增、克隆及测序
用特异性引物PCR扩增lmo2486基因,产物经10 g/L琼脂糖凝胶电泳检测得到1 212 bp的目的条带(图1)。胶回收与pMD19-T(simple)载体连接,转化到DH5α后通过菌液PCR筛选阳性转化子(图2),进行测序分析,结果显示该基因序列与GenBank中公布的序列一致。
2.2 lmo2486基因核苷酸及其编码氨基酸序列分析
由GenBank中提供的序列可知,LM-SB5lmo2486基因全长1 212 bp,编码403个氨基酸。lmo2486蛋白含3个N-糖基化位点(137-140、201-204、317-320),1个cAMP和cGMP依赖性蛋白激酶磷酸化位点(277-280),10个酪蛋白激酶Ⅱ磷酸化位点(8-11、65-68、66-69、193-196、206-209、241-244、279-282、321-324、348-351、396-399),7个蛋白激酶C磷酸化位点(43-45、139-141、144-146、176-178、261-263、288-290、348-350),5个N-豆蔻酰化位点(15-20、16-21、26-31、297-302、344-349)(图3)。
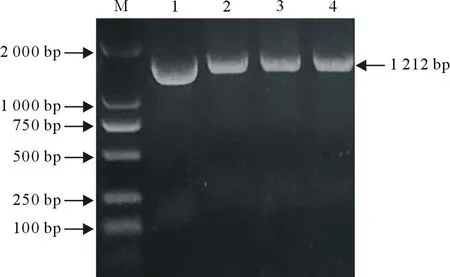
M.DNA标准DL 2 000;1.阳性对照;2~4.菌液PCR的产物

3个N-糖基化位点(阴影部分),1个cAMP和cGMP依赖性蛋白激酶磷酸化位点(加粗部分),10个酪蛋白激酶Ⅱ磷酸化位点(下划线),7个蛋白激酶C磷酸化位点(双下划线),5个N-豆蔻酰化位点(方框)
2.3 Lmo2486蛋白的分子结构特征分析
通过在线软件分析表明,Lmo2486蛋白分子质量为64.6 ku,属于亲水性蛋白。利用在线软件SOPMA分析可知,Lmo2486蛋白主要由α-螺旋和无规则卷曲构成(图4A)。在线软件TMHMM Server v.2.0分析Lmo2486蛋白形成一个跨膜区(34-56)。SignalP 4.1 分析表明,该蛋白无信号肽。SWISS-MODEL预测该蛋白可形成桶状的三级结构(图4B)。SMART预测并分析发现,该蛋白质具有PspC 蛋白域(3-63)和DUF4097 蛋白域(140-361)(图4C)。
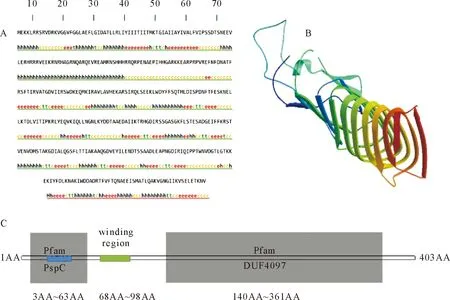
A.Lmo2486二级结构;B.Lmo2486三级结构;C.Lmo2486含有的主要结构域(PspC蛋白域位置:3~63,跨膜区:34~56,盘绕区:68~98,DUF4097蛋白域位置:140~361)
2.4 lmo2486基因序列的系统进化分析
将LM-SB5lmo2486核苷酸序列与LM不同分离株核苷酸序列以及同为PspC家族和DUF4097家族的不同菌种的基因进行同源性比对,结果显示,LM-SB5的lmo2486核苷酸序列与LM3株、02-6680株(4b型)、SLCC2378株(4e型)、ATCC 19117株(4d型)株、ICDC-LM188株、1/2b 10-0811株属于同一分支,与标准株EGD-e株的亲缘关系较近。与大肠埃希氏菌、阿尔多耶尔森氏菌( NZ-CP009781.1)、小肠结肠炎耶尔森氏菌株IP26014、亚历山大耶尔森氏菌株(NZ-CGBL01000009.1 )及马桑耶尔森氏菌株CCUG 53443属于不同分支,亲缘关系较远(图5),表明lmo2486基因在LM中高度保守。
2.5 lmo2486表达载体pET-32a(+)-lmo2486K的鉴定
lmo2486抗原集中区片段长度为1 074 bp。将pMD19-T-lmo2486K双酶切,得到2 692 bp和1 074 bp大小的条带(图6)。重组质粒pET-32a(+)-lmo2486K双酶切得到5 900 bp的载体条带和1 074 bp的目的条带(图7),表示成功构建了pET-32a(+)-lmo2486K重组质粒。
2.6 pET-32a(+)-lmo2486K重组质粒的诱导及蛋白的表达
将pET-32a(+)-lmo2486K转化至DE3感受态中,使用IPTG在16 ℃中振荡诱导Lmo2486的抗原集中区蛋白表达,经120 g/L SDS-PAGE检测出该重组蛋白的分子质量约是64.6 ku(图8),并在20 h~24 h时表达量较多。Western blot以鼠抗His单克隆抗体为一抗,带有His标签的山羊抗兔IgG为二抗,在64.6 ku处有特异性反应条带(图8),与预期值相符,分析lmo2486基因有着一定的反应原性,Lmo2486抗原集中区重组蛋白在大肠埃希氏菌BL21(DE3)中表达成功。

图5 基于lmo2486基因核苷酸序列进行系统进化分析(NJ法)

M.DNA标准DL 8 000;1~2.pMD19-T-lmo2486K双酶切产物

M.DNA标准DL 8 000;1~5.pET-32a(+)-lmo2486K双酶切产物
3 讨论
LM是一种食源性胞内寄生的革兰氏阳性菌,也是李斯特菌属中致病力最强的细菌,可感染人和动物,并突破机体的血脑及血胎等生理屏障,引起脑膜炎和流产等多种疾病[13]。LM拥有复杂的调节网络,其中包括LM毒力因子的转录调控和对环境应激耐受力的调节等,其中pfrA是许多毒力基因的转录调控因子,sigB可调控多种环境应激相关基因的表达,在LM应激反应中发挥着重要的功能[14-17]。LMlmo2486基因是一种含有PspC结构域的新型转录调控因子,可能参与LM在致病力和环境应激时的转录调控过程。同时,该蛋白DUF4097结构域是构成细菌黏附素的重要元件,可能参与LM黏附宿主细胞的过程,推测与细菌毒力密切相关。然而,目前对LMlmo2486的研究较少,至今其生物学功能尚不清楚。
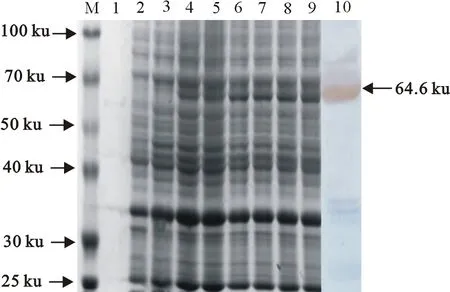
M.蛋白分子质量标准;1.蛋白上清液;2.pET-32a(+)空载体;3~9.IPTG诱导Lmo2486K蛋白0、4、8、12、16、20、24 h的表达产物;10.重组蛋白 pET-32a(+)-lmo2486K
PspC家族蛋白是一类应激反应调节因子,广泛存在于在革兰氏阴性肠杆菌科大肠埃希氏菌、小肠结肠炎耶尔森氏菌、肠炎沙门氏菌和鼠伤寒沙门氏菌中,主要参与细菌的应激反应和毒力的调节。目前研究发现,细菌的存活取决于它们感知和响应细胞包膜紊乱的能力。噬菌体休克蛋白(Psp)系统在包膜应激反应中起关键作用。在大肠埃希氏菌中,PspC家族蛋白参与应激反应系统以维持细菌细胞膜的完整性和功能[10,18]。PspC和PspB是两种细胞质膜蛋白,可以形成整合复合物,在小肠结肠炎耶尔森氏菌和大肠埃希氏菌中作为Psp反应的感觉激活剂[19-20]。在乳酸乳球菌中,YthA蛋白是PspC家族的应激反应性转录调节因子,在氨基酸生物合成、嘧啶代谢和胞外多糖的生物合成中发挥了调控作用,同时YthA有助于耐酸性[18]。在小肠结肠炎杆菌中,小肠结肠炎耶尔森氏菌的Psp系统对毒力至关重要,PspC 缺失突变后毒力显著降低[19,21-22],其生长也明显比野生型慢[21,23-24]。霍乱弧菌的Psp反应在压力源存在的情况下,内膜蛋白PspC可参与感知损伤[25]。PspC在肺炎球菌菌临床分离株中普遍存在,且PspC蛋白可以保护小鼠免受致命肺炎链球菌的攻击[26]。
本研究成功克隆了lmo2486基因并对其编码的蛋白进行了分子特征分析,发现其含有PspC结构域和DUF4097结构域。系统进化分析发现,该基因在LM中具有高度保守性。在此基础上,通过SDS-PAGE和Western blot证实重组蛋白Lmo2486K具有一定的反应原性,为揭示LMlmo2486对LM在环境应激和致病力调控作用奠定了前期的研究基础。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
现代畜牧科技(2021年9期)2021-10-13
江西农业学报(2021年4期)2021-04-20
生物信息学(2020年1期)2020-05-16
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
天津科技大学学报(2016年1期)2016-02-28
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年2期)2015-02-06
