
RVA谱特征值的影响因素及其与稻米食味品质的关系综述
2023-02-15 01:44岳红亮张梦龙程新杰宛柏杰朱静雯唐红生孙明法
江苏农业科学 2023年1期
岳红亮, 张梦龙, 程新杰, 刘 凯, 宛柏杰, 朱静雯, 唐红生, 孙明法
(江苏沿海地区农业科学研究所,江苏盐城 224000)
水稻是我国重要的粮食作物,播种面积常年稳定在3 000万hm2,占粮食作物总播种面积的25%以上。随着我国种植产业结构的调整,“优质”已成为水稻育种的首要目标,快速黏度分析仪(rapid visco analyzer,RVA)是目前水稻淀粉品质测定中重要的仪器之一。RVA最初是为了快速判定小麦穗发芽导致品质下降而开发的加热搅拌黏度仪,因其可以用少量的样品简便且迅速地测定黏度,1992年田縁勝洋应用RVA对稻米米粉样品进行黏度测定及其测定条件的分析,发现RVA可以用来测定稻米的黏滞性[1]。淀粉黏滞性谱别称RVA谱,是指将一定质量的米粉加入到一定体积的水中并搅拌均匀,搅拌中的米粉浆在快速黏度分析仪中经过一定时间的加热和冷却处理,淀粉黏滞性发生一系列变化而形成的特征曲线。RVA谱模拟了米饭的蒸煮过程,揭示了稻米淀粉在蒸煮过程中黏性随温度和时间的变化规律。本试验介绍了RVA的测定步骤及原理,RVA谱特征值QTL的定位研究、影响稻米RVA谱特征值的因素以及RVA谱特征值与稻米品质的关系,探讨了RVA谱特征值对稻米食味品质的评价效果,以期为稻米品质的深入研究提供参考,并为RVA谱的进一步推广应用奠定基础。
1 RVA的测定步骤及原理
RVA谱的测定可以分为6个阶段:(1)向称有3.0 g待测米粉样品的测试罐中加入25.0 mL去离子水,用桨叶将水和米粉搅拌均匀;(2)RVA以 960 r/min 快速搅拌10 s,然后降至160 r/min保持整个过程;(3)RVA将米粉浆从50 ℃均匀加热至95 ℃;(4)在95 ℃下保持2.5 min;(5)RVA将米粉浆从95 ℃均匀降至50 ℃;(6)RVA在50 ℃下保温1.4 min。整个测定程序时间为12.5 min(表1)。
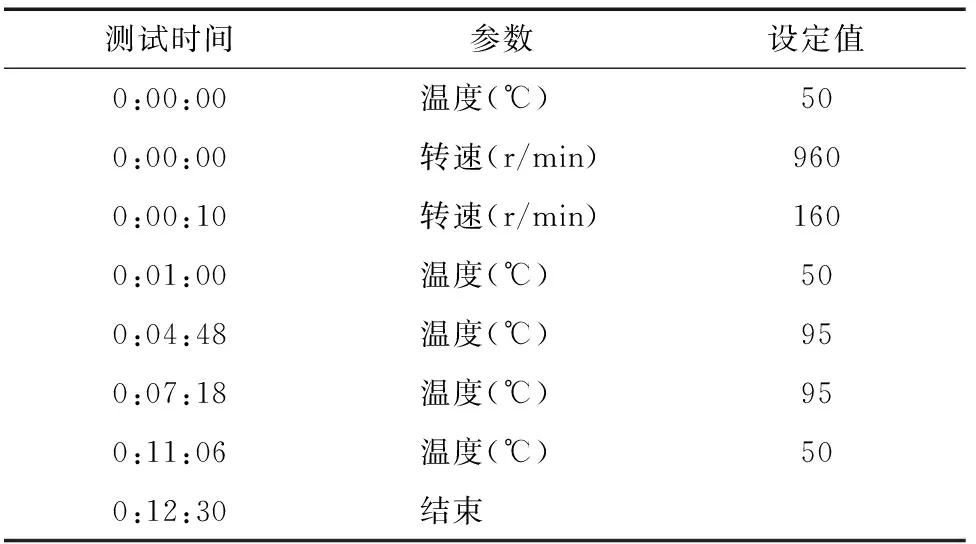
表1 快速黏度分析仪测定程序
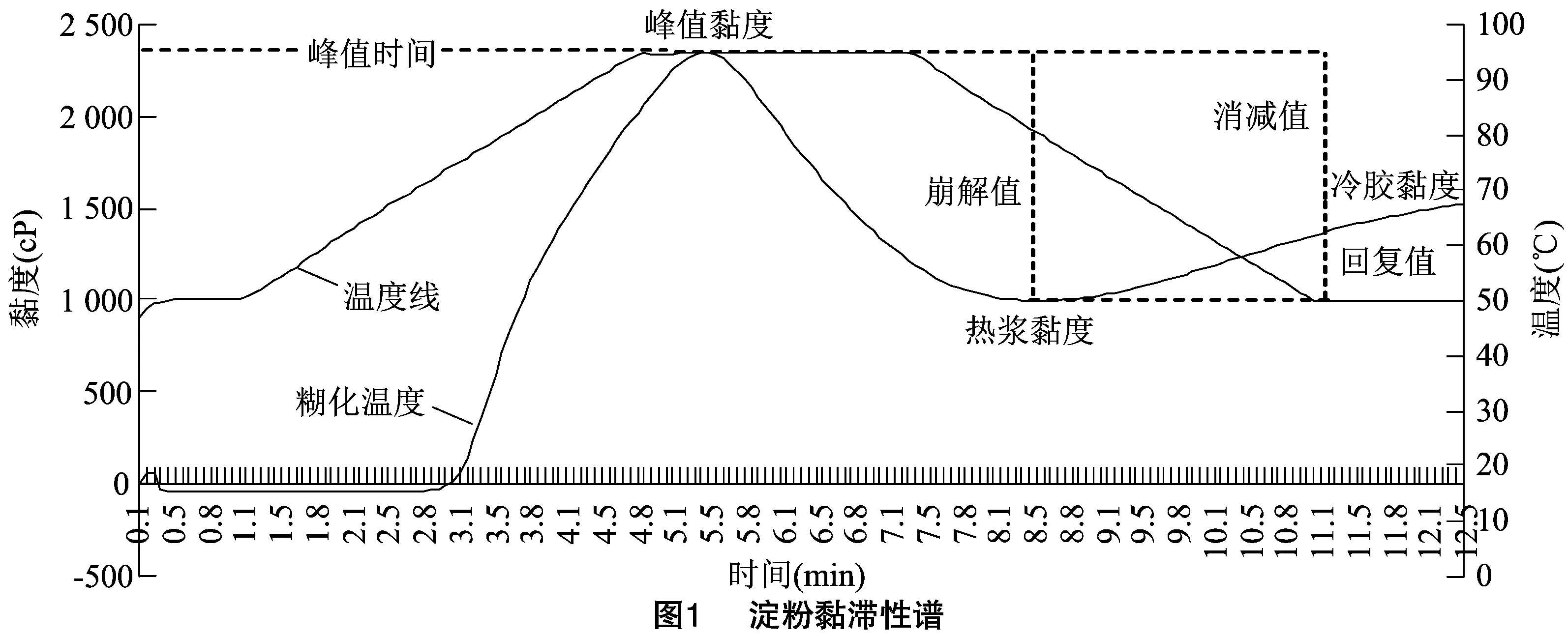
在RVA谱黏度曲线测定过程中,当淀粉与水搅拌混合时,水会进入淀粉内部,并与蛋白质等成分结合,但在温度低于50 ℃时,淀粉还未开始糊化,淀粉颗粒的膨胀很小,淀粉黏性基本没有变化。随着温度的升高,首先支链淀粉开始糊化,淀粉黏度达到峰值黏度后,由于淀粉颗粒水合物膨胀破裂导致直链淀粉和分子量较小的支链淀粉浸出,淀粉黏度下降。持续高温使得淀粉颗粒的结晶区域融化,更多的淀粉颗粒破裂,黏度进一步降低。随着温度的降低,淀粉凝胶冷却回生淀粉颗粒重新排列,形成晶体化的结构,淀粉黏度再次增加。在这一阶段,淀粉黏度与直链淀粉含量有关,较低的回复值表明直链淀粉含量较低[2-3]。
RVA谱中可以得到的一级参数有峰值黏度(peak viscosity,PKV)、热浆黏度(hot viscosity,HPV)、冷胶黏度(cool viscosity,CPV)、峰值时间(peak time,PeT)、糊化温度(pasting temperature,PaT)以及由一级参数通过计算而来的崩解值(breakdown viscosity,BDV)、消减值(setback viscosity,SBV)、回复值(consistency viscosity,CSV)等,其计算方式分别为BDV=PKV-HPV、SBV=CPV-PKV、CSV=CPV-HPV,黏度值以cP(centi poise)为单位(图1)。其中峰值黏度又称为最高黏度,热浆黏度又称为最低黏度,冷胶黏度又称为最终黏度,糊化温度又称为起浆温度、成糊温度[4],崩解值又称为衰减值,回复值又称为回冷值、回生值[5-6]。
2 RVA谱特征值QTL的定位研究
RVA谱特征值是由多基因控制的数量性状,且受基因和环境共同作用,对其研究工作较复杂。Zhang等通过构建染色体片段置换系群体,对表观直链淀粉含量相近的水稻群体和亲本进行研究,在第2、第5、第7、第8号染色体上鉴定出qPKV2-1、qSBV2-1、qPKV5-1、qHPV5-1、qCPV5-1、qPKV7-1、qHPV7-1、qCPV7-1、qSBV7-1、qPKV8-1等10个稳定的QTL,其贡献率范围为-95.6%~47.1%[7]。Shar等通过双单倍体群体在3个种植季节对RVA谱特征值的遗传位点进行分析,检测到4个稳定调控CPV、CPV、SBV、PeT的QTL均位于RM6 775~RM3 805区间[8]。包劲松等构建了双单倍体群体及分子连锁图谱,发现稻米RVA谱特征值主要受Wx基因控制,且在第1、第5号染色体上存在共同调节BDV和SBV的微效QTL,在第12号染色体上存在调控PKV和BDV的微效QTL[9]。张巧凤等利用F2群体对RVA谱特征值进行遗传分析,发现与包劲松等的结论[9]相似,并发现PeT可能由2对主基因控制,且还受微效多基因的影响;此外,通过构建重组自交系还检测到位于第2号染色体RM341~RM475标记之间的qHPV2、qCSV2和位于RM573~RM250标记之间的qCPV2;在第2号染色体上与支链淀粉合成有关的基因Sbe3和SssⅡ-2与RVA谱特征值有密切关系[10]。张杰等通过回交重组自交系群体在定位区间C470-R1 944、G264-G177和R2 829-R2 401发现4个尚未报道过的基因位点,分别是qCPV1、qBDV4、qPKV7、qHPV7[11]。Yao等利用重组自交系群体,在第2、第6、第7、第11号染色体上检测到分别位于RM7 581-STS、RM190~RM217、RM11~RM1 279和RM2 064~RM144区间的4个含有稳定表达的QTL的多效性区域[12]。
3 影响水稻RVA谱特征值的因素
3.1 淀粉对RVA谱特征值的影响
淀粉作为稻米最主要的成分,以淀粉粒的形式存在于细胞中,在含水量为14%的精米中,淀粉含量占总质量的75%以上[13]。根据淀粉的分子结构不同,可将淀粉分为直链淀粉和支链淀粉2类。岳红亮等对江苏省内粳稻品种进行研究,发现直链淀粉含量(amylose content,AC)较低的粳稻品种PKV和BDV较高,HPV、SBV和PeT较低[14]。赵春芳等对江苏省普通粳稻与东北粳稻RVA谱进行比较,发现两者RVA曲线相似,CPV近似或略高于PKV,而江苏省半糯粳稻的RVA曲线与前两者有显著差异,具体表现为PeT提前,HPV较低,CPV显著低于PKV,因而具有较高的BDV和较低的SBV[15]。朱艳菊对415份按不同AC值分类的水稻品种进行RVA谱特征值比较,发现HPV、CPV、CSV随AC值的升高而增大,且AC值在 15%~22%时相关性最好,PKV、BDV随AC值先增大后减小,SBV随AC值先减小后增大[16]。张蓉对仅表观直链淀粉含量不同的水稻株系RVA谱进行研究,发现随着表观直链淀粉含量的升高,RVA谱的PKV、HPV、CPV也逐渐升高,PeT延迟;对糯-极低(1.20%~3.29%)、低(13.13%~13.91%)、中(21.39%~22.84%)、高(25.26%~25.93%)4组表观直链淀粉含量相似的水稻品种RVA谱进行比较,发现具有相近表观直链淀粉含量的水稻品种,其RVA谱也有较大差异[17]。可见,在一定范围内直链淀粉含量的升高会减小PKV和BDV,增大HPV、CPV、CSV、SBV、PeT等特征值,但直链淀粉含量差异还不足以完全解释淀粉对RVA谱特征值的影响。
赵春芳等对支链淀粉结构与RVA谱特征值进行研究,发现南粳系列水稻品种支链淀粉短链A链[6≤支链淀粉链长聚合度(DP)≤12]含量与HPV、CPV、CSV呈显著负相关,支链淀粉B1链(13≤DP≤24)含量与HPV、CPV呈显著正相关[18]。李丁鲁等对长江下游地区部分优质粳稻品种与越光稻米支链淀粉结构进行比较,发现支链淀粉的短链部分(5≤DP≤12)的占比和支链淀究短/长键[Fa(5≤DP≤12)/Fb3(37≤DP≤58)]与PKV和BDV均呈显著正相关[19];蔡一霞等发现长链部分(44≤DP≤47和DP>100)与PKV和BDV呈极显著负相关[20]。赵营等认为,支链淀粉含长链的占比越高,淀粉粒越不易充分糊化,PKV和BDV将降低,影响米饭口感;支链淀粉中短链含量高,利于淀粉粒的糊化,易形成较高的PKV和BDV,使米饭的口感较好[21]。对不同类型水稻品种按AC值分组后进行研究,发现支链淀粉结构主要与淀粉的起始成糊温度和相对结晶度相关,与淀粉RVA谱特征值关系不密切[22]。周慧颖等认为,在低AC值水稻品种中,支链淀粉的平均链长、平均外链长与起始糊化温度、最高糊化温度、终结糊化温度、SBV呈正相关,与PKV、BDV呈负相关;支链淀粉A ∶B值(每条B链上所具有的A链数量)与起始糊化温度、SBV呈负相关,与PKV、BDV呈正相关[23]。张超等认为,在直链淀粉含量相同的前提下,支链淀粉链长越长,分支结构越多,平均链长越长,双螺旋结构越多,结晶度、PaT越高,BDV越小[24]。可见,支链淀粉中短链部分的含量越高越利于淀粉的糊化,PKV和BDV值也会越高。
3.2 蛋白对RVA谱特征值的影响
陈能等认为,蛋白是稻米中的第二大成分,稻米中的大部分蛋白以贮藏性蛋白形式存在,一般占糙米干质量的8%~11%[25]。张启莉等认为,按溶解性可将其分为碱溶性谷蛋白、醇溶性醇溶蛋白、盐溶性球蛋白、水溶性清蛋白(别称白蛋白),谷蛋白占贮藏蛋白的75%~90%,醇溶蛋白、球蛋白、清蛋白所占比例分别为1%~5%、2%~10%、2%~5%[26]。
石吕等认为,蛋白含量高的米饭质地较硬,口感较差,还会对RVA谱特征值产生显著影响[27]。谢新华等认为,随着稻米种蛋白含量的增加,RVA谱曲线呈整体下降趋势,RVA谱的PKV、HPV、BDV、CPV、CSV、SBV随蛋白含量的增加呈显著或极显著降低[28]。郭涛等认为,随着稻米蛋白含量增加,淀粉RVA谱曲线也呈整体下降趋势,不同的是只有PKV、HPV、CPV均呈下降趋势,BDV和SBV先升高后降低,PaT和PeT无显著变化[29]。谢新华等通过中性蛋白酶与超声波结合去除糙米中蛋白,去除蛋白质后的米粉RVA谱与未去除蛋白的相比,RVA谱曲线整体下降,糙米中的蛋白使RVA特征值的PKV、HPV、CPV等值升高,去除蛋白后RVA谱曲线上升段的斜率降低[30]。谢黎虹等在米粉中分别添加一定量的二硫苏糖醇和蛋白酶,发现PKV、HPV、CPV有不同程度的降低,并认为蛋白通过水合作用和二硫键形成的蛋白网络结构改变了淀粉的糊化特性[31]。可见,在一定范围内蛋白含量增加会显著降低PKV、HPV、CPV、BDV、SBV等特征值,造成淀粉RVA谱曲线整体下降。当人为去除米粉中的蛋白、降低米粉中蛋白含量时也会造成RVA谱曲线整体下降,且这种现象在直链淀粉含量越低时情况越显著。
张欣等对水稻4种蛋白组分进行分离并与RVA特征谱进行分析,发现醇溶蛋白含量和总蛋白含量与PKV、BDV呈极显著负相关,与SBV呈显著和极显著正相关;谷蛋白含量仅与SBV呈显著负相关,谷醇比与PKV、BDV呈极显著正相关,与SBV呈显著负相关[32]。周显青等对籼糯米逐步去除蛋白,发现糯米粉的PKV和BDV能够显著降低,其中去除谷蛋白影响最大,其次是球蛋白、醇溶蛋白;去除球蛋白能显著降低米粉的CSV;去除球蛋白和谷蛋白后,米粉PeT延长、PaT升高[33]。陈书强等认为,粳稻不同粒位间籽粒的总蛋白及其4种组分含量与PKV、HPV、BDV、SBV的相关性显著[34]。吴洪恺等通过对谷蛋白相对于醇溶蛋白含量差异较大的2种基因型LGCLGC(谷蛋白含量较醇溶蛋白低)和基因型lgclgc(谷蛋白含量较醇溶蛋白高)稻米的RVA谱特征值进行研究,发现LGCLGC型稻米的总蛋白含量与BDV和SBV存在显著的负相关和正相关;lgclgc型稻米的总蛋白含量与SBV和CSV都存在极显著的负相关,且LGCLGC型稻米RVA谱特征值的BDV和CSV分别显著和极显著大于lgclgc型[35]。郭涛等对低谷蛋白品种圣稻LG03和其背景亲本圣稻735进行研究,发现谷蛋白含量降低会导致PKV、HPV、CPV、BDV、PaT降低和PeT减少,CSV和SBV增加[36]。因此,研究蛋白对RVA谱特征值的影响不仅需要从稻米总蛋白含量上进行分析,还要对4种蛋白组含量及各组分之间的比例进行研究。
3.3 耕作措施及环境对RVA谱特征值的影响
稻米食味品质是一个受自身遗传因素和生长环境等多因素控制的复杂性状。同一品种的RVA谱特征值在不同环境和栽培条件下均有一定的差异[37-38]。因此,研究RVA谱特征值在不同种植条件下的性状表现对改善稻米品质具有重要意义。对半糯粳稻进行地点和播期试验,发现除PKV外,HPV、CPV、BDV、SBV、CSV、PeT等在不同地点间的差异均达到极显著水平;PKV、HPV、CPV、CSV、PeT与播期相关性显著;推迟播种会导致粳稻PKV、HPV、CPV、PeT、CSV上升[39]。对不同播期和种植地点的南粳46 RVA谱特征值进行研究,发现随着播期的推迟,PKV、HPV、CPV、CSV、PaT呈先降后升趋势,BDV先升后降,PeT无明显变化规律。随着纬度的降低,PKV、HPV、BDV、CPV总体上均呈先降后升的变化趋势,SBV和PeT降低,PaT升高[40]。对不同生态类型的粳稻进行播期与种植地点研究,发现品种对RVA谱特征值的影响最大,不同种植环境下RVA谱特性差异明显,CPV和CSV呈现北高南低的趋势,SBV随纬度的升高呈增加趋势。PaT和PeT在不同纬度间差异较小。随着播期的推迟,PKV、HPV、CPV、PeT呈减小趋势,而SBV和CSV呈增加趋势,BDV呈先升后降的趋势,PaT表现为先降后升的趋势。RVA谱特征值在不同种植地点随播期的变化趋势基本一致,且在江苏省北部地区受播期的影响较显著,同一品种在适宜种植区域内均以早播为佳[41]。收获期对RVA的影响因品种不同和种植环境不同有较大差异[42-44]。
徐正浩等对水稻不同灌溉方式进行研究,发现水胁迫环境下会降低稻米的PKV、HPV、CPV、BDV,增加SBV[45]。刘立军等认为,水稻结实期轻度干-湿交替处理可以提高稻米的BDV,降低SBV,而重度干-湿交替处理结果[46]则相反。相较于深水和浅水的供水方式,湿润灌溉会获得较高的PKV和HPV[47],与淹水管理和旱栽管理相比,节水灌溉和间歇灌溉方式有利于获得更高的PKV、HPV、BDV,降低CPV、SBV、CSV,获得较优的稻米品质[48]。对水稻氮肥施肥期进行研究,发现水稻进入抽穗期尽早施入穗肥,可以获得更高的BDV和较低的SBV[49]。从拔节期到齐穗期,增施镁肥可以显著降低稻米的CPV、CSV、SBV,增加PKV、HPV、BDV;然而穗肥时期镁锌配施会降低PKV、HPV、BDV,提高SBV和PaT,导致食味品质变差[50-51]。与常规栽培处理相比,生物菌肥的施用能在不改变稻米直链淀粉含量和蛋白含量的情况下显著提高稻米的PKV、BDV,降低SBV[52]。遮阴避光处理对水稻淀粉糊特性有显著影响,且在生育中后期的影响大于生育前期,但不同品种对遮阴的反应也有明显差异[53]。随着海拔高度的升高,稻米的PKV、CSV、PaT会降低,灌浆结实期间日均温度降低和日照时数减少导致PKV、HPV、CPV显著下降;秧龄和移栽方式发生改变,RVA谱特性也会发生变化,PKV和CPV均随秧龄的增加而升高,双苗手插处理的PKV、HPV、CPV均高于单苗优化定抛处理[54]。与常规耕作方式相比,有机耕作能提高稻米PKV、CSV、PaT等主要糊化特性[55]。
3.4 其他因素对RVA谱特征值的影响
除了稻米自身所含的淀粉、蛋白和农田耕作管理等因素,稻米中的脂肪含量以及稻米的储藏条件、蒸煮工艺等对RVA谱特征值也有很大影响。在不同储藏条件下,储藏温度、储藏时间和稻米含水量对稻米RVA特征值均有显著影响,其中温度为主要因素,随着储藏时间的延长,稻米的PKV和BDV在储藏期先升高后降低,高温(30、35 ℃)储藏条件下,稻米的PKV和BDV在后期降低更快;CPV、CSV、SBV、PaT在整个储藏期内呈增加趋势,高温储藏下CSV、SBV、PaT的变化明显大于其他储藏温度(25、20、15 ℃)[56]。RVA特征值在储藏1~5个月内变化较小,而6~7个月期间变化较大,籼稻类储藏过程中变化较大的特征值主要为PKV、BDV、PeT,粳稻类为HPV、CPV、CSV,而粳糯类为PKV、BDV、CSV、PaT,大部分特征值变化差异达到显著或极显著水平;且不同类型稻谷在储藏期间RVA谱特征值发生明显差异的时间点不同,粳糯类水稻品种在储藏期间发生明显差异的时间点明显早于粳稻和籼稻类[57]。Wongsaipun等认为,PKV和CPV可以作为模型参数来判断稻米的贮藏时间,且这种方法对糯米的预测结果更加准确[58]。因此,在稻米储存和保鲜过程中应对不同类型的水稻品种提供适宜的储藏条件,同时储藏时间不宜过长。吴洪恺等认为,稻米米粉经脱脂处理后PKV和HPV显著高于未脱脂米粉,说明米粉脂肪含量升高会降低米粉黏度[59]。对大米进行微波处理能够降低PKV、HPV、CPV、CSV,且PaT和SBV均随着微波功率和处理时间的增加而升高;微波功率(400、640 W)和处理时间(60、90 s)对RVA特征值均有显著影响,800 W的微波功率及120 s的微波时间影响达极显著水平,原因可能是微波处理对淀粉颗粒的结构和形态有修饰作用,使淀粉颗粒间空隙变大,大米在蒸煮时可渗入更多水分,蒸煮品质得到改善[60]。
4 RVA谱特征值与食味品质的关系
RVA谱模拟了米饭的蒸煮过程,将加热过程中糊状米粉黏性随温度和时间的变化以特征值的形式反映出来。因此,稻米RVA谱特征值与稻米食味品质必然有很大相关性[61]。
目前评价稻米食味品质的指标除RVA谱特征值外,主要有直链淀粉含量、蛋白含量、胶稠度和食味值,其中食味值又分为米饭食味值和米粒食味值。马会珍等认为,不仅RVA谱特征值之间相关性显著,且RVA谱特征值与直链淀粉含量、蛋白含量、米饭食味值相关性也显著[62-68]。其中多数研究结果显示,直链淀粉含量与HPV、CPV、SBV、CSV、PeT、PaT等显著或极显著正相关,与BDV显著或极显著负相关,与PKV的相关性在不同的研究中结果不一致;蛋白含量与CPV、CSV、PeT、PaT显著或极显著正相关,与BDV显著或极显著负相关,与PKV、HPV、SBV的相关性在不同的研究中结果不一致;米饭食味值与BDV、PKV显著或极显著正相关,与SBV、CSV、CPV、PeT、PaT显著或极显著负相关[62-68],与HPV的相关性在不同的研究中结果不一致,这些差异可能是由不同研究者所采用的品种类型不同以及是否进行分组分析造成的。舒庆尧等认为,食味较好的水稻品种大多表现为BDV>100 RVU,而SBV<25 RVU,且多数为负值;食味较差的品种大多表现为BDV>35 RVU,而SBV>80 RVU[69]。Allahgholipour等认为,除BDV和SBV外,PKV和CSV和对食味品质有较好的评价效果[70-71]。Champagne等认为,感官评价中米饭内聚性、黏性、适口性和质构分析仪中的黏附性与RVA谱特征值有较高的相关性,其中BDV与米饭黏性的相关性最强[72]。因此,可以通过CPV、CSV、PeT、PaT、BDV等判断稻米直链淀粉含量和蛋白含量,通过BDV、PKV以及SBV、CSV、CPV、PeT、PaT等鉴定米饭食味值。
5 结论与展望
黏度特性是稻米食味品质的重要性状,在稻米食味品鉴活动中,黏度特性作为评价稻米食味品质等级的重要参考指标[73-74],对优质稻米的选育和品鉴都具有十分重要的指导意义。RVA特征值是反映稻米黏度特性的重要参数指标,近年来随着对稻米食味品质研究的深入,对RVA特征值影响因素的研究以及调控RVA特征值主效和微效基因的定位工作都取得了较好的进展,RVA特征值与米饭食味值的相关性也得到较明确的研究结论。这些都为RVA特征值在稻米品质鉴定及育种中的应用提供了更加准确的理论依据,并为RVA谱的进一步推广应用奠定了基础。
随着分子技术的不断发展和完善,分子标记辅助选择技术也越来越多地应用在水稻黏度基因鉴定和水稻育种的研究中。应当充分利用黏度特性对稻米黏性基因进行挖掘,明确不同基因位点与RVA谱特征值的相关性,通过分子标记辅助选择快速准确地培育出携带不同主效和微效基因的种质资源,加速种质资源的创新,通过与基因紧密连锁的标记进行辅助选择,提高育种的准确性,缩短育种年限,加速品质育种进程。
猜你喜欢
粮油食品科技(2020年6期)2020-12-08
红楼梦学刊(2020年2期)2020-02-06
中国粮油学报(2019年4期)2019-07-12
现代装饰(2018年4期)2018-05-22
中国塑料(2016年2期)2016-06-15
实用手外科杂志(2015年2期)2015-08-28
饲料博览(2015年4期)2015-04-05
北京航空航天大学学报(2014年1期)2014-12-19
食品科学(2013年22期)2013-03-11
食品科学(2013年22期)2013-03-11
