
腹内侧前额叶在内隐认知重评中的因果作用*
2023-02-13 02:12高可翔张岳瑶李思瑾袁加锦张丹丹
心理学报 2023年2期
高可翔 张岳瑶 李思瑾 袁加锦 李 红 张丹丹,3
(1 四川师范大学脑与心理科学研究院,成都 610066) (2 深圳大学心理学院/磁共振成像中心,深圳 518060)(3 深港脑科学创新研究院,深圳 518055)
1 引言
情绪调节对个体维持心理健康、适应社会生活十分重要(Gross & John,2003)。广义的情绪调节是指在情绪发生过程中通过改变认知、行为和环境等来影响情绪的产生、体验和表达(Gross,1998)。情绪调节可通过不同的方式、使用各种策略达成(Braunstein et al.,2017)。以往研究主要关注外显(有意识的、主动的)情绪调节,而近来一些研究发现,情绪调节过程也可以是内隐(无意识)的,且极少或不需要消耗以外侧前额叶(lateral prefrontal cortex)为代表的认知控制资源(Gyurak et al.,2011;Koole et al.,2015;Koole & Rothermund,2011)。内隐情绪调节能有效降低负性情绪体验及相关生理反应(李红,袁加锦,2018;Mauss et al.,2007;Wang & Li,2017;Williams et al.,2009;Yang et al.,2015;Yuan,Ding,et al.,2015;Yuan,et al.,2019;Zhang et al.,2020)。然而与外显情绪调节不同,目前我们对内隐情绪调节的认知及脑机制的了解还非常有限。
情绪加工神经模型(Etkin et al.,2015;Phillips et al.,2008;Rive et al.,2013)指出,外显情绪调节需要调用负责认知控制的外侧前额叶皮层,包括背外侧前额叶(dorsolateral prefrontal cortex)和腹外侧前额叶(ventrolateral prefrontal cortex)。而内隐情绪调节则节省认知控制资源,它主要依赖内侧前额叶(medial prefrontal cortex)和前扣带回皮层(anterior cingulate cortex,ACC)工作。不少研究者认为,腹内侧前额叶(ventromedial prefrontal cortex,vmPFC)可能是内隐情绪调节的关键脑区。vmPFC 在情绪加工和调节过程中不仅通过与杏仁核、颞叶、腹侧纹状体的功能连接,提取情绪相关的先验信息,对情绪内容进行评估并更新刺激的情感价值,同时还负责整合腹侧系统和背侧系统的信息,调控杏仁核的活动从而改变情绪刺激诱发的神经和行为反应(Delgado et al.,2016;Diekhof et al.,2011;Hiser & Koenigs,2018;Motzkin et al.,2015;Roy et al.,2012)。目前共有两项脑调控-脑观测的证据支持vmPFC 在内隐情绪调节过程中的关键作用。这两项研究同时采用了功能核磁共振成像(functional magnetic resonance imaging,fMRI)和经颅直流电刺激(transcranial direct current stimulation,tDCS)技术。其中一项研究发现,采用tDCS 激活内侧前额叶(包括vmPFC 和亚属前扣带皮层)之后,实验组被试在观看负性视频片段时报告的负性情绪强度显著低于tDCS 伪刺激组报告的评分(Abend et al.,2019)。另一项发现,采用tDCS 激活vmPFC 条件相比于伪刺激条件,被试在最后通牒游戏中被诱发愤怒情绪时,vmPFC 激活增加,而愤怒情绪体验脑区ACC 和前脑岛的激活减弱,且被试对不公平提议的接受率增加,自我报告的愤怒程度降低,随后的攻击行为降低,此结果提示vmPFC 在减少愤怒体验和愤怒表达方面具有重要作用(Gilam et al.,2018)。需要注意的是,这两项研究仅考察了激活vmPFC 所带来的情绪改善,没有对情绪调节进行实验性操纵,因此难以保证被试情绪的改善是由内隐情绪调节带来的。针对此问题,本研究将内隐情绪调节作为核心自变量进行操纵,建立内隐情绪调节和情绪改善间的因果关系,并通过 tDCS 激活vmPFC,观察被试内隐情绪调节效果的改变,为揭示内隐情绪调节的核心脑区提供因果性证据。
最常用的内隐情绪调节诱发方式是通过启动(priming)建立情绪调节目标,使个体对该情绪调节目标进行自动化追求(automatic goal pursuit) (Custers&Aarts,2010)。例如,Yang 等人(2015)使用含有情绪调节语义的目标词来启动“调节情绪反应”这一目标,被试每次从两个备选词中选择一个与目标词的语义相匹配的选项,从而通过语义加工过程为被试“植入”情绪调节目标。该内隐情绪调节的启动方式称为“词语匹配任务”。又如,Yuan 等人(2019)要求被试从打乱顺序的五个词中挑选四个词组成一个语法通顺的句子,而句子中每次都含有一个情绪调节策略的相关词(例如“接受”、“顺其自然”),这样被试也可以通过语义加工被植入某种具体的情绪调节策略(例如“接受策略”,acceptance)。该内隐情绪调节的启动方式称为“句子整理任务”。已有研究发现,以启动方式诱发的内隐情绪调节,不但能有效降低负性情绪体验,且对外侧前额叶的认知资源消耗较少(Mauss et al.,2007;Williams et al.,2009;Wyczesany et al.,2021)。例如,Zhang 等人(2020)发现,内隐情绪调节可降低杏仁核和脑岛等情绪体验脑区的激活,同时不会引起额顶控制脑网络的明显激活。事件相关电位(event-related potential,ERP)证据也显示,使用内隐情绪调节下调负性情绪时,反映负性情绪强度的顶区LPP 波幅减小,且不会诱发额区P3 波幅的明显增强,后者被认为是认知控制卷入的指标(Yuan,Long,et al.,2015;Yuan,2019)。
与外显情绪调节相比,内隐情绪调节在某些情境下或在某些人群中具有较明显的优势。例如Williams 等人(2009)发现,在高压力损害认知资源的情境下,极少消耗认知资源的内隐认知重评(cognitive reappraisal)是比外显认知重评更适合的调节方式。更重要的是,对认知控制资源严重受损的抑郁症等精神疾病患者而言,虽然他们外显情绪调节策略(例如认知重评)的使用效率和使用频率均低于健康个体(Campbell-Sills et al.,2006;Garnefski et al.,2007;Garnefski & Kraaij,2006;Gross & John,2003),但患者的内隐情绪调节功能没有明显受损。例如,研究者发现抑郁被试在内隐启动的分心(distraction)策略下能有效降低在挫折任务中的消极情绪,但让被试执行外显分心策略时却不能达到此情绪调节效果(李红,袁加锦,2018)。由于内隐情绪调节具有上述优点,阐明内隐情绪调节的认知神经机制具有重要的理论价值和临床指导意义。
本研究针对内隐情绪调节的潜在核心脑区vmPFC,将tDCS 组别作为被试间变量,比较tDCS激活组和伪刺激组之间内隐情绪调节效果的差异。情绪调节可以通过多种策略实现,其中认知重评策略最具环境适应性,该策略强调认知的改变,即通过重新解释情绪刺激的内容(例如,从更加积极的角度看待问题)从而调节情绪反应(Gross & John,2003;Ochsner et al.,2012;Suri et al.,2015)。本研究使用含有认知重评相关词的句子整理任务,启动认知重评目标,诱发被试对负性情绪认知重评的自动化追求(Williams et al.,2009;Yuan,Ding,et al.,2015)。实验中,采用图片浏览任务诱发被试的负性情绪,并记录脑电数据。行为层面,我们采集被试观看每张图片后的情绪自我报告评分以及任务结束半小时后的图片效价评分,用以考察内隐情绪调节及阳极激活vmPFC 的即时效果和短时间内的持续效应(Mauss & Robinson,2009)。电生理层面,本研究采用 ERP 的顶区晚正成分(late positive potential,LPP)作为情绪反应的客观指标。LPP 通常在情绪刺激呈现500 ms 后出现,在顶区达到最大值(Foti & Hajcak,2008;Hajcak & Nieuwenhuis,2006)。该成分反映了刺激的情绪强度(Hajcak et al.,2010;Liu et al.,2012),在情绪调节相关研究中被用于反映情绪调节的效果(Schönfelder et al.,2013)。例如,以往研究表明,下调负性情绪伴随着LPP 波幅的降低(He et al.,2020;Yuan,Long,et al.,2015),且LPP 波幅和主观负性情绪评分之间存在显著相关(Li et al.,2022;Zhao et al.,2021)。
基于已有发现,本研究假设:vmPFC 是内隐情绪调节的核心脑区,内隐认知重评可有效下调负性情绪体验。在观测指标上,被试在内隐认知重评启动条件比基线条件的情绪评分更加正性、顶区LPP波幅明显下降;相比于伪刺激组,采用tDCS 激活vmPFC 可提高内隐认知重评的调节效果。本研究不但能深化我们对内隐情绪调节脑机制的理解,还可为采用tDCS 提高内隐情绪调节能力、改善抑郁症等人群的情绪调节障碍提供临床指导。
2 方法
2.1 被试
本研究招募在校本科生和研究生共80 名。所有被试均为右利手,无神经系统疾病及脑损伤史,参与实验时身体健康,视力或矫正视力正常。实验前签署了知情同意书,实验后获取报酬100 元。本研究方案得到了深圳大学伦理委员会的批准。根据相关研究(Abend et al.,2019;Gilam et al.,2018)报告的效应量大小(平均=0.24),使用G*Power 3.1.7 对样本量进行估计(α=0.05,β=0.2,方差分析:重复测量,被试内-被试间交互作用),两组被试各含7 名即可达到80%的统计检验力(power)。因此本研究的样本量满足要求。
实验当天被试填写以下量表:贝克抑郁量表第二版(Beck Depression Inventory Second Edition,BDI-II;Beck et al.,1996)、斯皮尔伯格特质焦虑量表(Trait form of Spielberger’s State-Trait Anxiety Inventory,STAI-T;Spielberger,et al 1983)、情绪调节问卷的认知重评子量表(Emotion Regulation Questionnaire,ERQ-R;Gross & John,2003)。随后将被试随机分配到阳极刺激组和伪刺激组。经检验,两组在年龄、性别、抑郁水平(BDI-II)、特质焦虑水平(STAI-T)、认知重评策略的使用倾向(ERQ-R)上均无显著差异,如表1 所示。

表1 本研究两组被试的人口学特征
2.2 实验过程
本研究为2 (启动类型:认知重评/基线) × 2(tDCS 组别:阳极刺激/伪刺激)的混合实验设计。其中启动类型为被试内变量,tDCS 组别为被试间变量。
主实验由两个block 组成,分别对应“基线”和“认知重评”两种启动条件(见图1A)。两个block 的先后顺序在被试间平衡,block 间被试进行60 min的无关任务。每个block 包含句子整理(60 个试次)和图片浏览(30 个试次)两种任务。每个block 开始前,被试接受电流强度为1.5 mA 的tDCS (阳极刺激组的电流持续10 min,伪刺激组只在开始时给予1 min 的电流)。tDCS 结束后立刻开始实验。被试首先进行10 试次的句子整理任务,紧接着进行5试次的图片浏览任务,反复6 轮。认知重评启动紧跟简短的图片浏览任务,这样可以保证较强的启动效应。
句子整理任务用于启动内隐情绪调节。在句子整理任务中(图1B)被试需在30 s 内从5 个打乱顺序的词语中挑选4 个词,组成一个语法正确的句子(Mauss et al.,2007)。被试需使用电脑小键盘将4 个词的序号依次输入屏幕中的灰框内。认知重评block 中,每个整理好的句子均包含一个与认知重评策略相关的词或短语,基线block 中则不包含任何与情绪调节相关的词或短语。研究表明,整理包含认知重评词义的句子,可启动对情绪情境进行认知重评的自动化目标追求,从而达到内隐情绪调节的效果(Williams et al.,2009;Yuan,Ding,et al.,2015)。
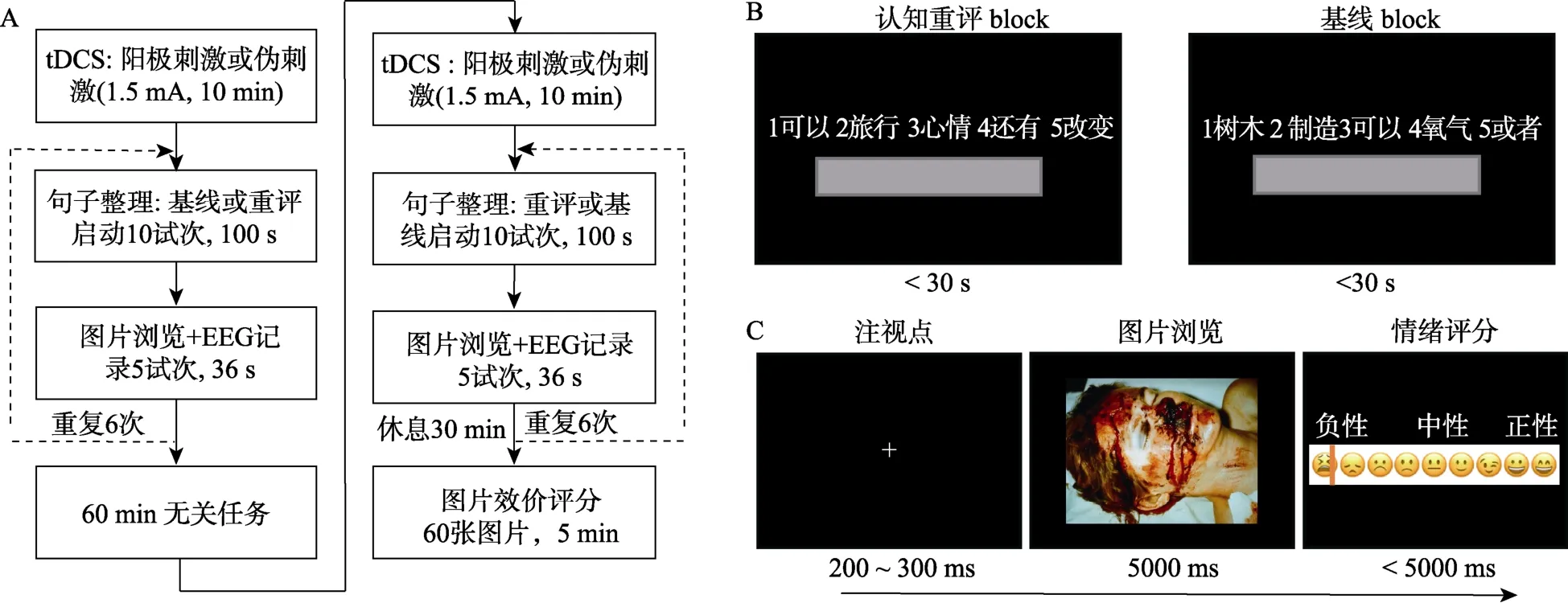
图1 实验过程。A,任务流程图。B,句子整理任务的材料示例。C,图片浏览任务的单试次流程。
图片浏览任务用于诱发被试的负性情绪。在图片浏览任务中(图1C),每张负性图片呈现5 s,在此期间被试需要仔细观察图片并体验情绪,图片消失后,在0~1 连续标尺中报告自己当前的情绪效价。情绪评定通过鼠标单击表情轴完成,越靠近表情轴左端(0 分)代表情绪越负,越靠近轴的右端(1 分)代表情绪越正,鼠标点击后屏幕上会出现红色竖条(提示点击位置),被试需在5 s 内完成情绪评定。试次间的空屏间隔为1 s。
主实验结束半个小时后,被试需要对图片浏览中的60 张图片进行9 点效价评分(1=非常消极;9=非常积极),以考察内隐情绪调节及tDCS 的持续效应。
为了确保内隐情绪调节操纵的有效性,实验结束后对每名被试进行了问卷调查,考察他们是否猜到了实验目的,以及在实验中是否主动进行了情绪调节。
2.3 实验材料
从国际情感图片系统(International Affective Picture System,IAPS;Lang et al.,1997)和中国情绪图片系统(Chinese Affective Picture System,CAPS;白露 等,2005)共挑选负性图片60 张。额外招募20名同质被试对这60 张图片的效价和唤醒度进行评定。随后这些图片被平均分配到两个block 中。独立样本t检验(双尾)表明,图片的效价和唤醒度在两个block 间均无显著差异,图片的效价为(t(58)=0.13,p=0.893;2.54 ± 0.11 vs.2.52 ± 0.14),唤醒度为(t(58)=-0.50,p=0.622;6.31 ± 0.16 vs.6.44 ±0.19) (1~9 点评分)。实验过程中,所有图像以相同的亮度和对比度显示在LCD 屏幕的中央,视角为3.0× 3.5°。
实验共使用10 个认知重评相关语句和10 个情绪调节无关语句。认知重评相关句中含有一个与认知重评策略相关的启动词。例如,“旅行可以改变心情”中的“改变”。情绪调节无关句中不含有任何与情绪调节策略相关的词汇。例如,“树木可以制造氧气” (图1B)。额外招募20 名同质被试对这20 个句子的效价和熟悉度进行评定。独立样本t检验(双尾)表明,认知重评相关句和情绪调节无关句在效价和熟悉度上均无显著差异,句子的效价为(t(18)=0.26,p=0.794;6.18 ± 0.10 vs.6.13 ± 0.17),熟悉度为(t(18)=0.96,p=0.342;5.99 ± 0.12 vs.5.83 ± 0.11)(1~9 点评分)。这20 个句子在实验中重复使用6 次,每次使用时随机设置新的词序并加入不同的填充词(或称为干扰词),因此被试在不同的试次中对相同的句子也需要重新进行语义加工和句子整理。
2.4 tDCS 参数设置
本研究采用离线tDCS 刺激方案,主要原因有二:在线的tDCS 会在脑电数据中造成电流伪迹;电流刺激过程可能引起电极周围皮肤的不适感,影响被试执行实验任务。
采用无线tDCS 系统(NeuStim,博睿康科技股份有限公司,中国常州)。靶点脑区为vmPFC。参照同样选择vmPFC 为靶点脑区的tDCS 研究,本研究将阳极电极置于国际脑电10/20 系统中的Fz 点和Fpz 点之间的位置,阴极电极置于下巴中央,同时电流强度设置为 1.5 mA (Bulteau et al.,2022;Junghofer et al.,2017;Winker et al.,2018,2019,2020;Yin et al.,2021)。根据Junghofer 等人所进行的基于有限元的tDCS 电流正向建模表明,这种电极位置和电流参数设置可使电流有效到达vmPFC,并最大限度地减少对其他脑区的影响(见Junghofer等人2017 研究的图1)。
通过一对用盐水浸泡过的3 cm × 3 cm 大小的海绵电极施加1.5 mA 的恒定电流。为减少电刺激引起的不适,在刺激的开始和结束阶段分别设置了30 s 的电流逐渐增强(fade in)和逐渐减弱(fade out)的过程。阳极刺激组的被试接受1.5 mA 电流10 min。伪刺激组被试只在这10 min 中的最初1 min 接受逐渐增强又逐渐减弱的电流。已有研究表明,1 min 的电流刺激会让被试体验到与真刺激组相似的头皮感觉,但不会显著影响后续实验任务中大脑的神经活动(Mungee et al.,2014;Riva et al.,2015)。
被试在实验结束后报告自己在接受tDCS 电刺激过程中的负性体验,包括电刺激引起的生理难受程度和负性情绪程度等。1~9 点评分,分值越高表示负性体验越强烈。
2.5 脑电信号采集与分析
脑电信号采集使用32 通道无线放大器(NeuSen.W32,博睿康科技股份有限公司,中国常州)。采样率250 Hz,电极阻抗低于10 kΩ。在线记录时以左侧乳突作为参考电极,离线分析时以“全脑平均电位值”进行重参考。
使用Matlab (v2020a)软件对脑电数据进行预处理。以图片开始出现作为零时刻,依次进行以下处理:重参考、滤波(0.01~30 Hz)、分段(-200~5000 ms)、基线矫正(-200~0 ms)、剔除波幅超过±150 μV 的试次。顶区LPP 成分的波形使用Pz 及其周围电极(P3、P4、Pz、CP1 和CP2)的平均波幅计算,时间窗根据课题组已有文献选取(Li et al.,2022;Zhao et al.,2021):以经典的P3 成分结束为起点,内隐情绪调节结束为终点,即图片呈现后的1~5 s 作为LPP 时间窗。除了事前关注的顶区LPP,我们在脑电数据中还发现枕区的P1 成分受实验变量的显著影响。对于枕区P1 成分(事后分析),采用O1、O2、P7、P8 的平均波幅来衡量,时间窗根据已有文献选取为 100~150 ms (Clark & Hillyard,1996;Gu et al.,2020;Zhang et al.,2022;Żochowska et al.,2022)。本研究关注启动类型和tDCS 组别对P1 波幅的影响,不考察P1 波幅的左右半球差异。
2.6 统计方法
统计分析采用SPSS Statistics 20.0 (IBM,Somers,USA)。描述性统计量表示为“均值 ± 标准误”。显著性水平设定为0.05。采用作为方差分析的效应值。对自我情绪评分、图片效价评分、顶区LPP 波幅、枕区P1 波幅分别进行两因素重复测量方差分析,被试内因素为“启动类型”,被试间因素为“tDCS 组别”。采用双尾皮尔逊相关对因变量进行探索性相关分析。
3 结果
3.1 行为结果
80 名被试中,仅5 名被试事后报告说他们察觉到了实验目的或在实验中主动进行了情绪调节(阳极刺激组3 名,伪刺激组2 名)。这说明大部分被试(94%)在实验中处于内隐情绪调节状态。有否剔除此5 名被试并不对统计结果产生明显影响。考虑到本研究主要关注tDCS 效应(即组间差异),本部分报告了所有80 名被试的实验结果。
两组被试对tDCS 电流刺激的负性体验均较弱,且不存在组间差异(表2)。自我情绪评分的结果表明,启动的主效应显著,F(1,78)=34.4,p〈 0.001,=0.306:认知重评启动条件下的情绪评分(0.30 ± 0.01)比基线条件更加正性(0.26 ± 0.01),该结果说明了内隐情绪调节启动的有效性。tDCS 组别的主效应不显著,F(1,78)=1.6,p=0.211,=0.020:阳极刺激组的情绪评分(0.29 ± 0.01)比伪刺激组稍偏正性(0.27 ± 0.01)。最主要的发现是,启动类型和tDCS 组别的交互作用显著:F(1,78)=5.7,p=0.019,=0.068 (图2A)。简单效应分析表明,认知重评启动条件下阳极刺激组的情绪评分(0.32 ± 0.01)比伪刺激组更偏正性(0.28 ± 0.01),F(1,78)=4.2,p=0.043,=0.051;但此组间差异在基线启动条件下不显著,F(1,78)=0.2,p=0.670 (阳极刺激组:0.27 ± 0.02,伪刺激组:0.26 ± 0.02)。探索性分析发现,情绪评分结果与被试的特质焦虑和抑郁水平成反比(表3,图2C)。
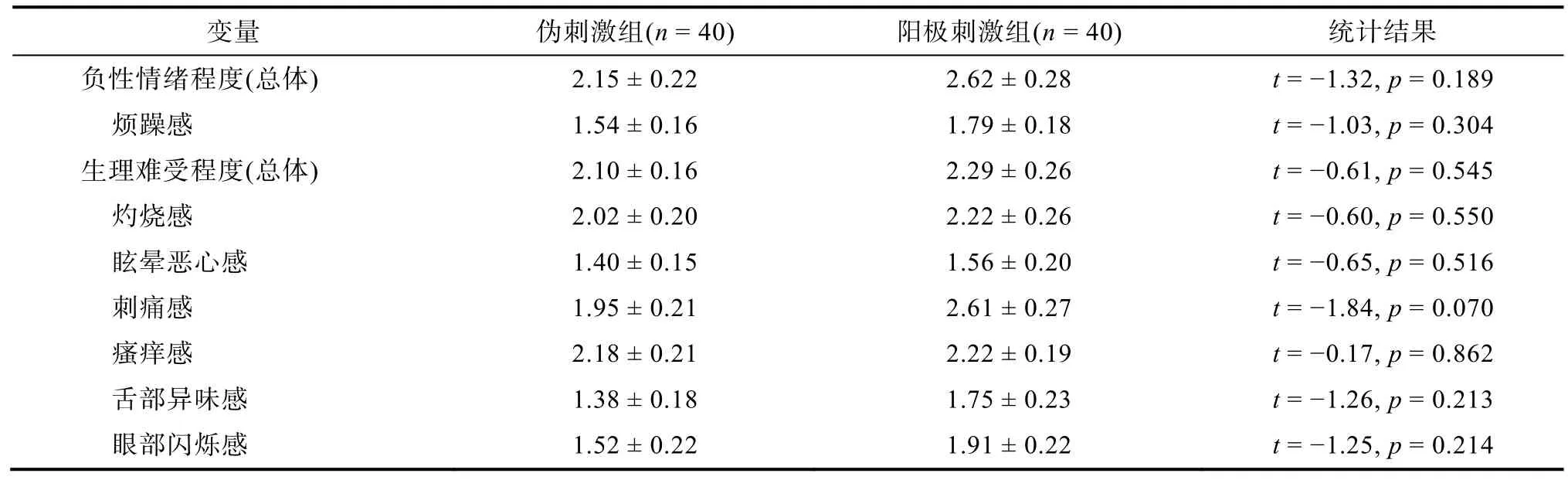
表2 两组被试对电流刺激的负性体验
主任务结束半个小时后的图片效价评分结果表明,启动类型的主效应边缘显著,F(1,78)=3.8,p=0.055,=0.046:认知重评启动条件下的效价评分(2.96 ± 0.10)比中性启动条件(2.90 ± 0.94)更偏正性。tDCS 组别的主效应显著,F(1,78)=9.2,p=0.003,=0.106:阳极刺激组的效价评分(3.38 ±0.14)比伪刺激组(2.75 ± 0.14)更加正性(图2B)。
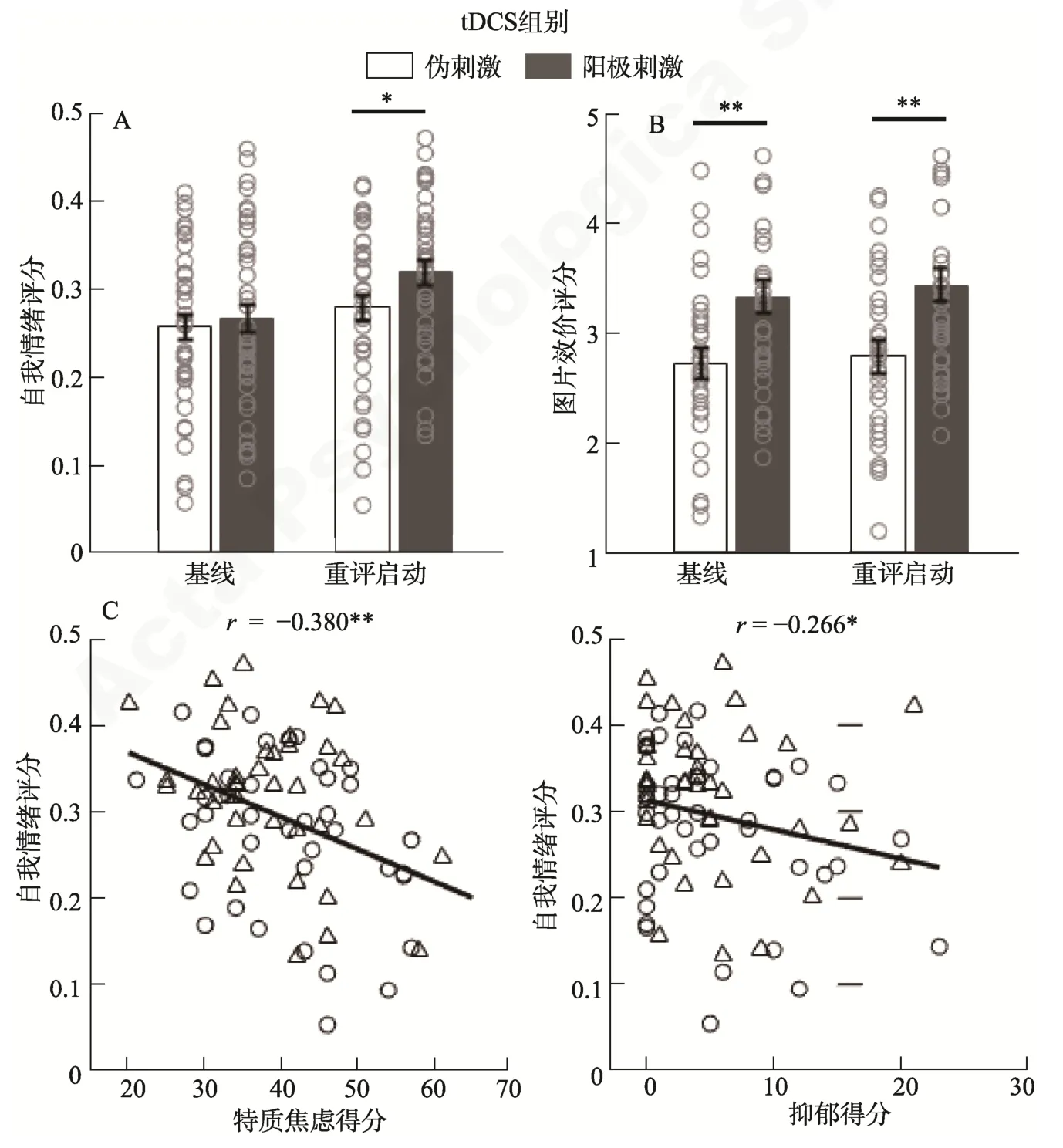
图2 行为结果。A,自我情绪评分。0~1 计分:0 分和1 分分别代表极端负性和极端正性,0.5 分代表中性。B,图片效价评分。9 点计分:1 分和9 分分别代表极端负性和极端正性,5 分代表中性。图中的误差条(error bar)代表标准误。小圆圈表示单个被试的数据。C,自我情绪评分与特质焦虑和抑郁得分的相关关系。为节省空间,图中仅画出总体样本(n=80)在重评启动条件下的相关。完整的相关结果见表3。小圆圈和小三角分别表示伪刺激组和阳极刺激组单个被试的数据。** p 〈 0.01,* p 〈 0.05。
3.2 脑电结果
LPP 波幅结果表明,启动的主效应显著,F(1,78)=70.3,p〈 0.001,=0.474:认知重评启动条件下的 LPP 波幅(1.39 ± 0.18)显著低于基线条件(2.50 ± 0.16)。tDCS 组别的主效应显著,F(1,78)=7.9,p=0.006,=0.092:阳极刺激组的LPP 波幅(1.51 ± 0.22 µV)低于伪刺激组(2.39 ± 0.22 µV)。最主要的发现是,启动类型和tDCS 组别的交互作用显著:F(1,78)=5.7,p=0.019,=0.068 (图3AC)。简单效应分析表明,认知重评启动条件下F(1,78)=11.0,p=0.001,=0.124,阳极刺激组的LPP 波幅(0.80 ± 0.26 µV)显著低于伪刺激组(1.99 ±0.26 µV),但此组间差异在基线启动条件下不显著,F(1,78)=3.1,p=0.080 (阳极刺激组:2.22 ± 0.22 µV,伪刺激组:2.78 ± 0.22 µV)。探索性分析发现,自我情绪评分结果与LPP 波幅成反比(表3,图3B)。
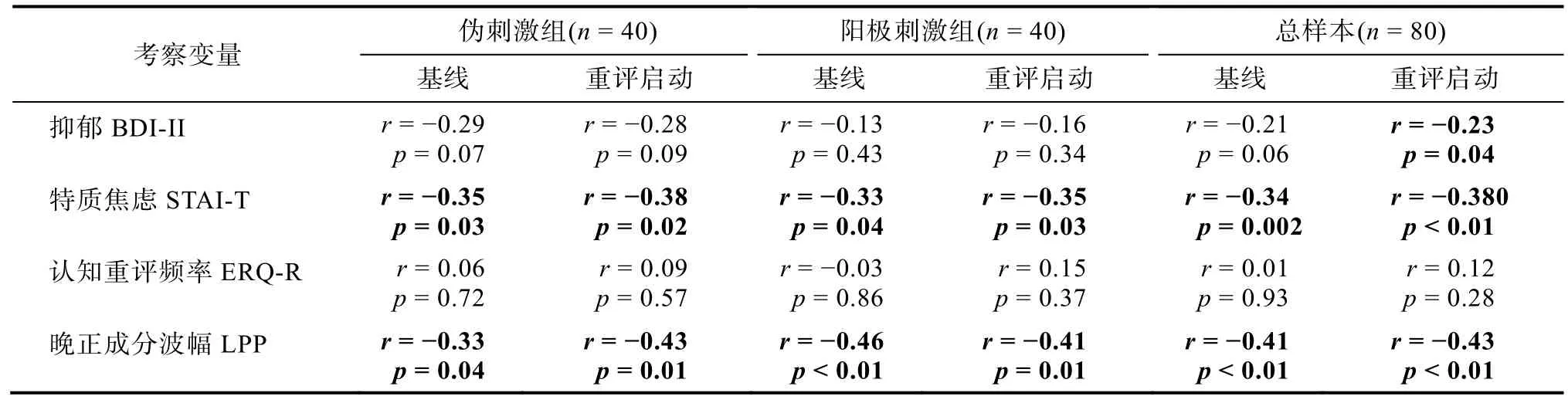
表3 自我情绪评分与被试特质及LPP 波幅的相关
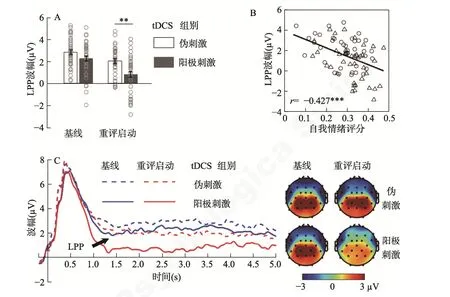
图3 顶区晚正成分(LPP)。A,LPP 平均波幅。图中的误差条(error bar)代表标准误。小圆圈表示单个被试的数据。B,自我情绪评分与LPP 平均波幅的相关。为节省空间,图中仅画出总体样本(n=80)在重评启动条件下的相关。完整的相关结果见表3。小圆圈和小三角分别表示伪刺激组和阳极刺激组单个被试的数据。*** p 〈 0.001,** p 〈0.01。C,LPP 波形图和地形图。波形为Pz、P3、P4、CP1 和CP2 的平均波幅。地形图为时间窗1~5 s 的平均值。
除了在假设阶段关注的LPP 波幅,我们还意外发现双侧枕区的P1 成分也显著受到实验变量的影响(图4):tDCS 组别的主效应显著,F(1,78)=6.1,p=0.016,=0.072:阳极刺激组的P1 波幅(4.71 ±0.40 µV)低于伪刺激组(6.12 ± 0.40 µV)。启动的主效应不显著,两个自变量的交互作用不显著。
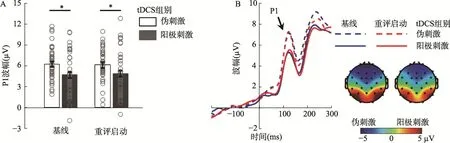
图4 枕区P1 成分。A,P1 平均波幅。图中的误差条(error bar)代表标准误。小圆圈表示单个被试的数据。* p 〈 0.05。B,P1 波形图和地形图。该波形为O1、O2、P7 和P8 的平均波幅。地形图为时间窗100~150 ms 的平均值。地形图中,由于未发现启动的主效应,合并“启动类型”的两个水平。
4 讨论
4.1 腹内侧前额叶激活的即时效应
本研究使用tDCS 以考察vmPFC 在内隐情绪调节中的因果作用。与预期一致,我们发现阳极激活以vmPFC 为代表的内侧前额叶导致了内隐情绪调节效果的显著增强。具体来说,我们发现内隐认知重评能同时下调负性情绪和脑电LPP 波幅,并且这种下调作用在激活vmPFC 的被试组中更显著。
本研究主要的贡献在于:我们通过操纵vmPFC的激活程度以考察该脑区在内隐情绪调节中的因果作用。启动类型和tDCS 组别的交互作用表明,vmPFC 在由启动诱发的内隐情绪调节中发挥了因果作用,激活vmPFC 可明显增强内隐认知重评对负性情绪的下调效果。虽然以往研究已提示vmPFC是内隐情绪调节的重要脑区(Braunstein et al.,2017;Etkin et al.,2015;Phillips et al.,2008;Rive et al.,2013),然而采用神经调控技术激活vmPFC,并对比考察启动和非启动的内隐情绪调节过程(对内隐情绪调节进行直接操纵),本研究尚属首次。值得注意的是,我们发现的情绪调节效应的组间差异仅在认知重评启动条件下出现,在基线条件下没有组间差异。与我们的发现不同,Abend 等人(2019)在没有启动内隐情绪调节的情况下(类似于我们的基线即中性启动条件),使用tDCS 激活vmPFC 并观察到了被试负性情绪的降低。我们认为这种不一致的结果可能与tDCS 电极位置的摆放不同有关。本研究中tDCS 的电极放置于前额顶部(Fz 点和Fpz 点之间)和下巴,由电场模拟图可以看出(Junghofer et al.,2017),如此设置电极位置可以使电流调控的脑区主要集中在vmPFC,同时可以减弱流经vmPFC 邻近脑区(例如背内侧前额叶和眶额叶)的电流强度。而Abend 等人(2019)的研究将两个电极分别放置于前额(鼻根上方)和后脑(枕骨隆突下方),这就使整个大脑的矢向轴(包括:vmPFC、ACC、后扣带回、内侧枕区等)都接受到了电流刺激,因此Abend 等人观测到的类似于情绪调节的效应很可能不是直接由vmPFC 受到调控引起的。
除了证明vmPFC 是内隐情绪调节的关键脑区,我们还意外发现,激活vmPFC 能在情绪刺激加工的早期阶段影响对刺激的注意。具体来说,我们发现在负性图片呈现后100~150 ms 的早期注意阶段,vmPFC 被激活的实验组比伪刺激组的P1 波幅更小。P1 成分反映了大脑对视觉刺激的早期注意,P1波幅越大代表被试对刺激投入的注意越多(Clark & Hillyard,1996)。例如,由于人们对负性情绪信息存在注意偏向,不少研究发现负性图片比正性和中性图片诱发更大的 P1 波幅(Carretié et al.,2004;Delplanque et al.,2004;Smith et al.,2003)。因此,本研究发现激活vmPFC 引起P1 波幅减小,表明tDCS阳极组被试对负性情绪刺激的早期视觉注意加工减弱。该结果提示,vmPFC 除了参与较晚期的情绪调节过程,还会在情绪信息的早期视觉加工阶段减少对负性刺激的注意。与我们的发现一致,Junghofer 等人(2017)结合脑磁图和tDCS 技术发现,激活vmPFC 导致早期(约100 ms)视觉注意相关的枕叶和颞枕皮层对负性刺激的响应减弱。
4.2 腹内侧前额叶激活的持续效应
最后本研究还发现,半小时后再次观看图片时,内隐情绪调节和阳极激活vmPFC 会持续影响被试对图片的再体验。具体来说,启动类型的主效应边缘显著,认知重评启动条件下的图片效价评分比基线条件更偏正性;tDCS 组别的主效应显著,vmPFC被激活的实验组比伪刺激组的效价评分更正性。以往研究发现,外显情绪调节会影响情绪刺激的再次体验(Erk et al.,2010;Hermann et al.,2017,2021)。例如,MacNamar 等人(2011)发现,对负性图片进行认知重评,30 min 后情绪调节效果依然存在,体现为与对照条件相比,认知重评条件的图片效价评分更正、LPP 波幅更小。同时,Walter 等人(2009)发现,间隔10 min 再次观看之前经过情绪调节的负性图片时,杏仁核的激活减少。因此,本研究结果表明,内隐情绪调节同样具有持续的改善情绪的潜力。但是也需要注意,本研究中我们发现的内隐情绪调节的持续效应较弱:启动类型对图片效价评分的主效应仅为边缘显著。这可能是因为短时的启动效应持续效果有限,后续研究应考虑使用多次训练的方式(例如连续训练一周,每天训练半小时)替代“启动”,从而提高内隐情绪调节的持续效果(Hopp et al.,2011)。此外,我们还发现tDCS 组别对图片效价评分有显著的主效应。先前有研究发现,激活vmPFC能影响被试再次面对负性刺激时的情绪反应,这可能与负性刺激的恐惧消退过程有关(Dittert et al.,2018;Guhn et al.,2014)。因此我们的发现可能表明,vmPFC 的激活促进了对负性图片的恐惧消退过程。
4.3 腹内侧前额叶在内隐情绪调节中的作用
综合已有研究和本文的结果我们认为,vmPFC在调节负性情绪时可能具有至少三个方面的作用。首先,vmPFC 在视觉加工的早期阶段通过影响对负性刺激的注意(Wolf et al.,2014)来调控自下而上的信息输入。接着,vmPFC 作为信息整合和价值编码的核心脑区对信息进行整合,并计算刺激的情感价值(Roy et al.,2012;Winecoff et al.,2013)。具体来说,vmPFC 通过与杏仁核、颞叶、腹侧纹状体的功能连接,提取相关的先验信息对刺激的情感价值进行评估(D’Argembeau et al.,2008;Sharot et al.,2007;Torrisi et al.,2018),并在情绪性信息加工的中晚期更新刺激的情感价值(Quirk et al.,2006;Roy et al.,2012;Winecoff et al.,2013)。最后在情绪反应阶段,vmPFC 作为抑制负性情绪反应的脑区,通过与杏仁核间的负向功能连接,自上而下地调节杏仁核的反应从而影响情绪表达(Diekhof et al.,2011;Johnstone et al.,2007;Motzkin et al.,2015;Urry et al.,2006)。例如,在恐惧消退过程中,vmPFC 负责消除编码在杏仁核中的条件性恐惧信号(Gottfried & Dolan,2004;Sotres-Bayon & Quirk,2010),并通过影响情绪信息的记忆促进再次暴露的恐惧消退(Maren & Quirk,2004;Raij et al.,2018)。值得注意的是,vmPFC 的上述功能在外显情绪调节过程中需要外侧前额叶的调制和监管(He et al.,2020;Li et al.,2022;Ochsner et al.,2002;Ochsner & Gross,2005;Smith & Lane,2015;Zhao et al.,2021),而在内隐情绪调节过程中vmPFC 可以相对独立的工作(Mauss et al.,2007;Zhang et al.,2020),几乎不会消耗外侧前额叶的认知资源(Yang et al.,2015;Yuan et al.,2019)。
4.4 主客观情绪测量指标的相关性
本研究的另一个贡献在于,我们明确了自我情绪评分在测量内隐情绪调节效果中的有效性。自我情绪评分是情绪体验的直接测量,常用于检验外显情绪调节的效果(Mauss & Robinson,2009),但其在内隐情绪调节研究中的应用存在争议。一些研究者提出,单侧的情绪评分(如只评负性或正性)可能会引起被试对实验目的的猜测,从而降低评分结果的可靠性(Haeffel & Howard,2010;Polivy & Doyle,1980)。因此一些内隐情绪调节研究并没有使用自我情绪评分作为调节效果的指标(Mocaiber et al.,2010;Payer et al.,2012;Williams et al.,2009)。本研究中,我们采用双侧评分方式(评分范围从负性到正性)来避免评分任务可能会引起的被试对实验目的的猜测。我们的结果表明,内隐认知重评启动可以有效降低自我情绪评分,并且自我情绪评分与客观指标——顶叶LPP 波幅呈显著的负相关关系,这表明自我情绪评分在测量内隐情绪调节效果中具有较高的准确性(He et al.,2020;Li et al.,2022;Schupp et al.,2000;Shafir et al.,2015,2016;Zhao et al.,2021)。
4.5 研究展望
下一步研究可以从以下4 个方面展开。第一,本研究验证了vmPFC 在内隐情绪调节过程中的因果作用,但测量的指标仅限于自我情绪评分和脑电信号。先前研究表明情绪调节也会引起皮肤电导反应(Raio et al.,2013)、心率变异性(Williams et al.,2009)以及瞳孔直径(Robinson et al.,2021)等外周生理指标的变化。未来研究可以考虑通过多种测量手段验证内隐情绪调节的效果。第二,本研究招募的被试均为健康大学生,激活vmPFC 促进内隐情绪调节的潜在临床应用价值还需要来自抑郁症等临床样本的实验支持。第三,本研究主要关注单次tDCS 干预和句子整理任务启动后的短暂的内隐情绪调节效应,未来研究可以考察多次tDCS 治疗并结合反复的启动训练对内隐情绪调节能力的长期提高效果。第四,本研究虽然采用 tDCS 瞄准了vmPFC,但无法避免电流同时流经vmPFC 的周边脑区(例如背内侧前额叶、眶额叶等),这影响了本研究结论的准确性。我们期盼有更多的方法学研究,致力于探讨如何更精确的采用tDCS 瞄准vmPFC 等较深层的脑区。已有研究提示,采用多电极模式可以较好地将电流集中到目标靶区,同时减少非目标脑区的电流密度(Martínez-Pérez et al.,2020;Sergiou et al.,2022)。后续研究可采用此方法进行尝试。
5 结论
为揭示vmPFC 在内隐情绪调节中的因果作用,本研究采用tDCS 阳极激活以vmPFC 为代表的内侧前额叶,并使用句子整理任务启动内隐认知重评。研究结果表明,激活vmPFC 能显著促进内隐情绪调节,在内隐认知重评启动条件下,vmPFC 激活导致观看负性情绪图片的被试报告的负性情绪更弱、被负性情绪图片诱发的顶区LPP 波幅更小。同时我们发现,激活vmPFC 能减弱被试对负性刺激的早期注意分配。本研究表明,以vmPFC 为代表的内侧前额叶是内隐情绪调节的关键脑区,临床可以通过激活vmPFC 并配合内隐情绪调节认知训练,提高情绪障碍等精神病患者对负性情绪的调节能力。
猜你喜欢
安徽医科大学学报(2022年11期)2022-11-01
心理学探新(2022年1期)2022-06-07
癫痫与神经电生理学杂志(2022年6期)2022-02-10
浙江大学学报(理学版)(2021年1期)2021-01-26
价值工程(2018年25期)2018-09-26
猪业科学(2018年5期)2018-07-17
健康管理(2017年3期)2017-04-20
西南交通大学学报(2016年4期)2016-06-15
华北电力大学学报(自然科学版)(2016年3期)2016-04-25
中国当代医药(2015年8期)2015-03-01
