
膳食纤维对大鼠钙平衡及骨生物力学性能的影响
2023-02-12 03:02覃香香周玉恒陈海珊蔡爱华刘金磊
食品科学 2023年1期
覃香香,周玉恒,陈海珊,蔡爱华,刘金磊
(广西壮族自治区中国科学院广西植物研究所,广西植物功能物质与资源持续利用重点实验室,广西木质纤维素生物炼制工程技术研究中心,广西 桂林 541006)
钙不仅是构成骨骼最主要的无机元素,同时广泛参与机体各项生理活动的调节,如凝血、神经元传递、肌肉收缩、细胞内信号传导、激素分泌、调节心律和血压等[1-2]。整个生命周期中充足的钙摄入对于骨骼健康和正常生理代谢十分必要[3-6]。钙摄入不足可增加人体罹患骨质疏松以及各种慢性代谢性疾病的风险,儿童缺钙还会引起骨骼发育不良[7]。据统计,目前中国人均每日钙摄入量仅为345.03 mg,远低于800 mg的每日需要量,钙摄入不足人口比例超过95%,在中国居民缺乏的营养素中排名第一[8]。
除了需要摄入足够的钙,钙的吸收和利用是衡量补钙效果的重要参数。有研究表明单纯地摄入钙和VD无法降低老年人骨折发生的比例[9-10]。影响钙吸收和利用的因素包括性别、年龄、药物、疾病、食物等。其中,食物的各种营养成分是影响钙肠道吸收和体内生物利用度的重要因子[11]。尤其是被誉为人体第七营养素的膳食纤维,不仅影响钙的吸收和利用,也影响体内骨转换、骨密度,是骨代谢中一个重要的调节因子[12-14]。
膳食纤维是指不能被人体消化液所消化,但是却能够被共生在肠道(主要是结肠)的微生物所发酵,并对机体代谢产生调节作用的植物成分(通常是多糖)。世界各国的卫生组织都对国民的膳食纤维摄入量给出指导(推荐每日摄入25~40 g膳食纤维),但在膳食纤维种类选择上没有进一步的建议。膳食纤维是一大类在结构单元、聚合度、化学键种类、分支程度和支链基团等结构上有很大差异的化学物质,它们在分子质量、溶解程度、黏稠度、生物可降解性等理化性质上都表现出显著不同[15]。一些研究发现不同结构的膳食纤维对脂代谢、糖代谢产生不同的影响[16-18]。但目前针对谷类、水果类、根茎类等食品中出现频率最高的一些大分子膳食纤维的比较研究仍然缺乏。
本实验通过7 种不同的纯化膳食纤维或膳食纤维混合物,以小麦麸皮作为参照,研究它们在钙的肠道吸收、血清钙水平、尿钙排泄、骨钙含量、骨最大载荷等方面的差异,分析其差异规律和可能原因,为进一步研究缺钙和骨质疏松的膳食预防提供理论依据。
1 材料与方法
1.1 动物、材料与试剂
70 只SPF级雄性SD大鼠(Rattus norregicus)购于湖南斯莱克景达实验动物有限公司(生产许可证号:SCXK(湘)2019-0004;使用许可证号:SYXK(桂)2020-0002),鼠龄2 个月,体质量约(200±20)g,适应性饲养2 周。饲养条件:温度25 ℃、相对湿度40%~60%,自由摄食、饮水,光照/黑暗周期为12 h/12 h。
蔗渣木聚糖和蔗渣纤维素为自制,制备方法参考文献[19-20];麦麸为普通市售;菊粉 青海威德特种糖业有限公司;魔芋胶 河南正兴食品添加剂有限公司;果胶烟台安德利果胶股份公司;氯化镧 天津市精细化工研究所;Ca、Mg(质量浓度1000 μg/mL)单元素标准溶液 国家标准物质研究中心;HNO3(优级纯)广州化学试剂有限公司;HClO4(优级纯) 天津政成化学制品有限公司。其余试剂均为分析纯。
1.2 仪器与设备
AG-201型电子式万能试验机 日本岛津公司;NOVAA400P原子吸收分光光谱仪 德国耶拿分析仪器股份有限公司;DK-98-11实验电炉 天津市泰斯特仪器有限公司;SX2-2.5-10A箱式电阻炉 上海索域试验设备有限公司;DB-1AB电热板 邦西仪器科技(上海)有限公司;AC9000全自动电解质分析仪 江苏奥迪康医学科技股份有限公司。
1.3 方法
1.3.1 动物饲养
将SPF级雄性SD大鼠随机分成7 组,每组10 只,每组大鼠在基础饲料中分别加入50 g/kg的纤维素、木聚糖、果胶、菊粉(果聚糖)、魔芋(葡甘露聚糖)、麦麸(对照组)、混合饲料(m(木聚糖)∶m(纤维素)∶m(果胶)∶m(菊粉)∶m(魔芋)=6∶0.5∶1∶2∶0.5)。基础饲料为美国营养学会(American Institute of Nutrition,AIN)所推出适用于成年期啮齿动物的纯成分饲料改良配方AIN-93M,每千克干饲料含钙5 g。
1.3.2 样品采集
大鼠饲养8 个月后进行尾部采血,尿液和粪便用代谢笼收集,收集前先将大鼠在代谢笼适应性饲养24 h,之后收集48 h粪便、尿液,并记录饲料48 h摄入量。大鼠饲养9 个月后进行宰杀,剥离后肢左右股骨。
1.3.3 钙吸收率测定
天然的吸附材料来源广泛,但会有吸附率低、吸附中药材里有效成分的缺点,可以针对这两点为参照对材料进行结构修饰后,增大其吸附量。使其不仅能应用于水体中重金属的脱除,还能对中药水提液的重金属离子达到理想的吸附效果。
收集48 h的粪便,70 ℃烘干48 h,然后研钵研碎成粉,称取1 g粪便干粉样品,放入瓷坩埚(40 mL),置于电炉上加热碳化至无烟,放入箱式电阻炉进行灰化。灰化条件为600 ℃持续8 h。待电阻炉冷却取出瓷坩埚,向瓷坩埚加入1 mL质量浓度100 g/L的HNO3溶液溶解,并用超纯水定容至100 mL,测定前用质量浓度10 g/L HNO3适当稀释,用原子吸收光谱仪测定422.8 nm波长处吸光度,以钙质量浓度为纵坐标,吸光度为横坐标绘制标准曲线,标准曲线方程为y=18.074x+0.2853(R2=0.9981)。饲料的处理方法和钙含量检测方法同粪便,计算48 h饲料摄入总量所含的钙总量,并换算钙日平均摄入量。钙吸收率计算如公式(1)~(3)所示。
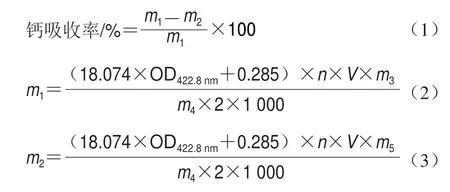
式中:m1为钙日平均摄入量/mg;m2为粪钙日平均排出量/mg;n为检测时样品稀释倍数;V为样品总体积/mL;m3为48 h内摄入的饲料总量/g;m4为预处理过程取样量/g;m5为48 h内粪便总量/g。
1.3.4 血钙浓度测定
剪尾采血1.0 mL,静置30 min,3000 r/min离心10 min,取上清液,采用电解质分析仪测定血清中的总钙、游离钙、结合钙浓度。
1.3.5 钙保留率测定
将48 h内收集的所有尿样混合均匀,10000 r/min离心10 min,量取尿液总体积,取1 mL澄清尿液,用质量浓度10 g/L HNO3稀释,并用原子吸收光谱测定其中的钙含量,计算每日尿钙日排出量。钙保留率计算如公式(4)所示。
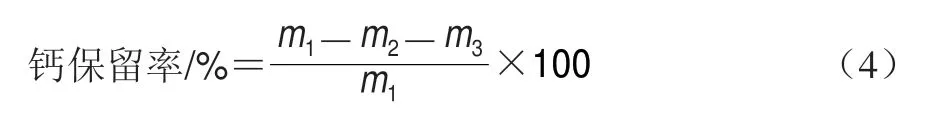
式中:m1为钙日平均摄入量/mg;m2为粪钙日平均排出量/mg;m3为尿钙日平均排出量/mg。
剥离的左股骨用超纯水洗净表面,放置烘箱烘至恒质量,然后放入坩埚内,置于箱式电阻炉中进行灰化,灰化温度为600 ℃,灰化时间为6 h,灰化结束后取出,在坩埚中加入5 mL质量浓度100 g/L的HNO3溶液溶解,再用超纯水定容至100 mL,适当稀释至检测值在标准曲线范围内,用原子吸收光谱仪测定骨钙含量。
1.3.7 右股骨三点弯曲实验
大鼠饲养9 个月后,处死后剥离后肢左、右股骨,右股骨用浸有生理盐水的纱布裹住,置于-20 ℃下保存。实验前取出冰冻的右股骨,解冻至室温25 ℃,在电子式万能试验机上进行生物力学性能测试。测试条件:跨距为20 mm、加载速率为5 mm/min,记录载荷变形曲线,在曲线上读出骨最大载荷参数。
1.3.8 原子吸收光谱法检测
取0.1 mL待测溶液,加入2 mL氯化镧溶液和7.9 mL超纯水,摇匀,原子吸收光谱仪进样检测,仪器工作条件见表1。
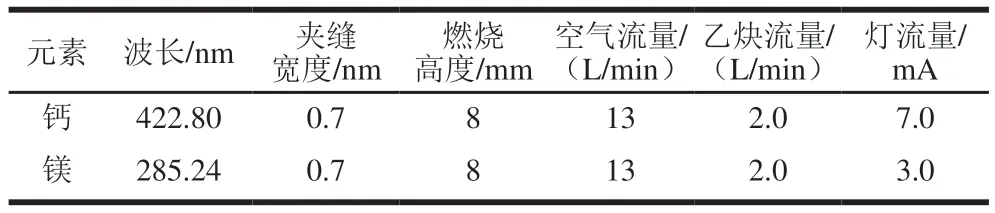
表1 原子吸收光谱仪工作条件Table 1 Working conditions of atomic absorption spectrometer
1.4 数据统计与分析
实验数据以平均值±标准差表示,利用SPSS 19.0统计软件进行单因素方差分析,P<0.05为差异显著。
2 结果与分析
2.1 不同膳食纤维对肠道钙吸收的影响
膳食钙是机体钙的唯一来源,钙的肠道吸收对机体钙平衡的维持至关重要[21]。与钙的肠道吸收有关的参数主要包括钙摄入量、粪钙排出量、钙吸收量、钙吸收率。从表2可以看出,木聚糖组钙日平均摄入量显著高于麦麸组(P<0.05),并高于其他各组,但由于木聚糖有促排便作用(数据未显示),导致木聚糖组粪钙日平均排出量大于其他组,且钙吸收量(P<0.05)和钙吸收率都显著或极显著低于麦麸组(P<0.01);在本课题组之前研究中发现魔芋组单位质量的粪便中钙含量偏高[20],本实验中虽然魔芋组排便量不大,但是粪钙日平均排出量也偏高,且钙吸收量和钙吸收率极显著低于麦麸组(P<0.01)和其他实验组。总地来说,麦麸组、纤维素组的钙吸收量、钙吸收率高于混合组、木聚糖、果胶、菊粉、魔芋组,其中麦麸组钙吸收量和木聚糖、果胶、魔芋组之间具有显著性差异(P<0.05、P<0.01),钙吸收率与木聚糖组和魔芋组具有极显著性差异(P<0.01)。

表2 不同膳食纤维对钙肠道吸收的影响Table 2 Effects of different dietary fibers on intestinal absorption of calcium
2.2 不同膳食纤维对血清钙水平的影响
从表3可以看出,血清游离钙浓度在不同膳食纤维组间无显著性差异(P>0.05),但是纤维素、木聚糖、果胶、菊粉组血清结合钙浓度和总钙浓度均显著小于麦麸组(P<0.05、P<0.01)。血清钙(主要是游离钙)的稳定对于维持细胞正常的生理功能至关重要,正常情况下,机体血钙水平受到甲状旁腺激素、降钙素和1,25-(OH)2D3的精确调节,通过骨骼、肾脏、小肠来共同维持血钙的动态平衡,保证机体正常生理功能的进行[21]。由表3可知,麦麸组在结合钙(游离钙的临时储库)浓度和总钙浓度上高于其他纯化的膳食纤维组,因此,麦麸更有利于稳定血钙。

表3 不同膳食纤维对血清钙水平的影响Table 3 Effects of different dietary fibers on serum calcium levels
2.3 膳食纤维对尿钙流失和钙保留率的影响
肾脏是调节钙平衡的一个重要的器官,其能够通过重吸收来减少钙的尿排泄。图1表明,相对麦麸组,纤维素组和木聚糖组可以显著或极显著减少尿钙流失(P<0.05、P<0.01);而果胶组尿钙日排出量极显著高于麦麸组(P<0.01),且明显高于其他各组。
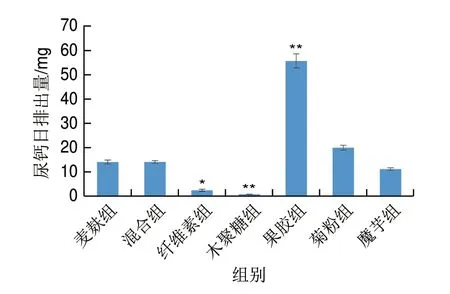
图1 不同膳食纤维对尿钙的影响Fig.1 Effects of different dietary fibers on urinary calcium levels
钙保留是钙摄入、钙的肠道吸收、尿钙排泄共同作用的结果。从图2可以看出,果胶组和魔芋组钙保留率极显著低于麦麸组(P<0.01),尤其是果胶组由于尿钙流失过多,出现了负钙平衡。总地来说,麦麸、混合物、纤维素组钙保留率高于木聚糖、果胶、菊粉、魔芋组。
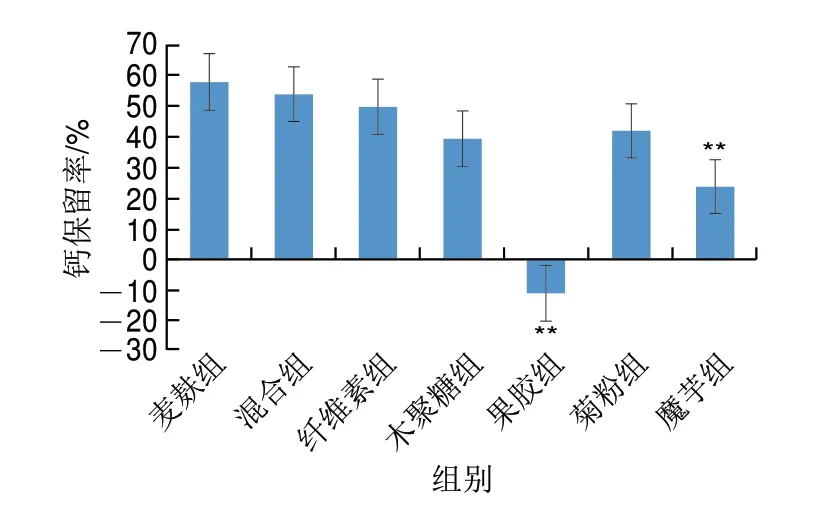
图2 不同膳食纤维对钙保留率的影响Fig.2 Effects of different dietary fibers on calcium retention
2.4 膳食纤维对骨钙含量和骨生物力学性能的影响
骨钙含量是钙平衡的结果,由表4可知,钙保留率比较高的麦麸组、混合组、纤维素、木聚糖组、菊粉组骨钙含量也比较高,除了果胶组骨钙含量显著低于麦麸组以外(P<0.05),其余各组与麦麸组并没有显著性差异(P>0.05)。
骨生物力学性能是骨强度的最直观体现,而骨最大载荷是骨生物力学性能的最主要评价指标,从表4可以看出,骨钙含量与麦麸组没有显著差异的木聚糖组(P>0.05),其骨最大载荷显著高于麦麸组(P<0.05),由此可见,骨强度不仅与骨钙含量有关,还与骨微结构、骨转化率、矿化程度、微损伤累积及修复、胶原矿物基等其余骨质量因素有关[22]。本实验结果同时表明,果胶组尿钙的过度流失会引起负钙平衡,使得骨钙含量大幅度降低,最终导致骨生物力学性能下降(图2、表4)。

表4 不同膳食纤维对骨钙含量和骨最大载荷的影响Table 4 Effects of different dietary fibers on bone calcium content and maximum load
3 讨论
3.1 膳食纤维结构对钙吸收的影响
食物中的钙盐以游离钙的形式从消化道进入人体体液循环的过程称为钙吸收。90%的钙吸收发生在小肠,其次是结肠。膳食纤维由于非消化的特性,可以完整地穿过小肠,到达结肠后被共生在肠道的菌群分解并代谢产生各种短链脂肪酸(short-chain fatty acids,SCFAs),降低了肠道环境的pH值。因此,有研究将一些膳食纤维促进钙吸收的作用归因于酸化肠道、SCFAs促进肠道上皮生长从而增加吸收面积等因素[23-24]。以此类推,钙吸收率应该和SCFAs的产量成正比。然而,本研究出现与上述理论不一致的结果,以纤维素和木聚糖为例,纤维素是β-葡聚糖的高度结晶体,具有最少的降解率和SCFAs产量;而木聚糖是多种单糖的不定型聚合物,虽然不溶于水,却具有很高的亲水性,在水中通常是呈现均匀的乳浊液状态,这种物理特性使得木聚糖具有极高的酶解可及性,很容易被肠道微生物利用,在大鼠的肠道末端的消化率可达到85%[25],因而具有远高于纤维素的SCFAs产量[26],但它们在钙吸收量和钙吸收率上却呈现出和SCFAs产量相反的结果。其他组别也出现类似的趋势,即低发酵型膳食纤维如麦麸、纤维素的肠道钙吸收量和钙吸收率高于高发酵型膳食纤维如木聚糖、果胶、菊粉、魔芋、混合组(表2)。Mneo等[27]利用Ussing chamber技术进行研究发现,在肠道中单纯降低pH值不会影响到钙吸收,因此,降低肠道pH值似乎不是膳食纤维促钙吸收的主要机制,单纯从结肠的发酵和SCFAs产量方面并不能完全解释膳食纤维对钙吸收的差异。
可溶性或分散性良好的膳食纤维如木聚糖、果胶、魔芋、菊粉、混合组比难溶的纤维素、麦麸在肠道中更容易与肠道内容物充分接触,它们本身的结构特点及理化性质增加了与矿质离子相互作用的可能性。例如,木聚糖的结构中除了木糖、阿拉伯糖、半乳糖等中性糖以外还含有葡萄糖醛酸基、乙酰基、未脱除干净的木质素酚基基团,果胶是半乳糖醛酸聚合物,本身带有阴离子基团且易溶于水,它们对矿质元素都具有吸附螯合和包裹的双重作用;魔芋是葡萄糖和甘露糖的聚合物,菊粉是果糖的聚合物,二者虽然不带电荷,但其大分子可溶物呈现的凝胶性能对肠道营养物质具有隔离、包裹作用。这种理化特性能够减少例如淀粉、脂肪等营养物与消化酶的接触,从而降低营养物质的吸收[28],这也是膳食纤维抗糖抗脂的机制之一,对于矿质元素也可能起着同样的作用。
麦麸和纤维素具有相似的钙吸收量和钙吸收率,它们都是难溶性膳食纤维。有研究报道了麦麸所含的植酸具有抗营养作用,是引起钙吸收减少的原因之一[29],但本研究结果中,麦麸组钙吸收率并不比那些不含植酸的纯化膳食纤维低。植酸一般存在于小麦坚硬的糊粉层细胞胞浆中[30-31],与坚固光滑的小麦皮层黏在一起,即使经过肠道消化过程可能也很难被释放出来,工业上提取也要经过比较剧烈的条件才能将植酸脱离附着物[32],这可能是麦麸组没有受到植酸干扰的原因。也有学者认为麦麸减少钙吸收的研究通常是短期的结果,从长期的适应性上看并没有对钙吸收产生负面影响。总体而言,钙吸收受到影响的因素很复杂,膳食纤维由于种类众多,理化性质表现出很大的差异,不同的膳食纤维对钙吸收的影响可能存在不同的机制。但作为吸收钙的主要部位,在小肠肠道内理化性质的不同所导致的钙吸收差异可能是比结肠发酵产酸更重要的一个机制。
3.2 不同膳食纤维对钙的体内利用的影响
在钙的体内利用阶段,膳食纤维的作用更多的是通过代谢产物进行间接的调节,只有代谢产物才能穿过肠道细胞屏障进入血液中,影响更远端的器官。目前已知的主要代谢产物是各种SCFAs[23],尤其是丁酸被认为具有可以抑制骨吸收、降低骨的溶解流失的作用[33]。有研究表明,粪便中总SCFAs浓度从大到小依次是混合组>木聚糖>魔芋>麦麸>菊粉>果胶>纤维素,丁酸浓度从大到小依次是木聚糖>菊粉>果胶>混合组>魔芋≈麦麸>纤维素[26],而本研究中骨钙含量从大到小依次是混合组≈纤维素≈菊粉≈麦麸≈木聚糖≈麦麸>魔芋>果胶,骨最大载荷从大到小依次是木聚糖>纤维素≈麦麸≈混合组≈菊粉≈魔芋胶>果胶。显然,不能完全以SCFAs或丁酸浓度来解释膳食纤维之间在骨钙含量和骨生物力学性能上的差异。Weaver等[34]的研究也证明骨的性能与SCFAs产量关系并不密切,与本研究结果一致。除了代谢产物,肠道微生物也是膳食纤维发挥作用的重要机制和当今研究的热点之一[35-37]。然而,无论是肠道微生物还是肠道代谢产物,膳食纤维如何影响肠道微生态,肠道微生态的改变又是如何精确联系到远端器官功能改变,它们联系的分子机制是什么,仍有待进一步的研究。
3.3 纯化膳食纤维能否作为麦麸的替代品
禾本科谷物是人类的主食之一,禾本科谷物膳食纤维是人们所需膳食纤维的最主要来源,粮食的精加工使得谷物膳食纤维成为现代饮食最缺乏的膳食纤维。与此同时,众多纯化的膳食纤维补充剂作为商品进入市场。以人们自然饮食中最典型的麦麸作为膳食纤维参照,比较几种不同结构的纯化膳食纤维对钙平衡及其影响,结果显示麦麸组在钙吸收率和钙保留率上均高于除纤维素以外的单一纯化膳食纤维,混合组比单一纯化膳食纤维更接近麦麸组。关于膳食纤维的补充原则,张厚德等[38]详细地论述了平衡膳食纤维对健康的重要性,本实验结果也进一步证明,单一纯化膳食纤维在维持钙平衡上仍然存在一定的缺陷,复合型膳食纤维更合理。
4 结论
不同结构的膳食纤维对钙的肠道吸收、钙在机体内的利用、钙在骨骼中的沉积、尿钙排泄量、钙保留和骨的生物力学性能等的影响存在差异,低发酵性膳食纤维有利于钙的吸收和更高的钙保留率。在纯化的膳食纤维中,木聚糖比麦麸更能显著提高骨的生物力学性能,而果胶的作用相反。
猜你喜欢
农家参谋(2021年6期)2021-08-03
山西冶金(2021年3期)2021-07-27
微生物学杂志(2020年5期)2020-12-23
农村农业农民·B版(2019年8期)2019-10-08
农村.农业.农民(2019年16期)2019-01-08
大众健康(2018年11期)2018-11-28
百科知识(2018年22期)2018-11-27
华声文萃(2018年8期)2018-09-18
饮食科学(2016年9期)2016-11-18
饮食科学(2016年3期)2016-07-04
