
重组大肠杆菌利用醋酸及乳酸为碳源合成 PHB
2023-02-10 06:18:22肖萌蒋莹崔逸璇SadafRiazMaurycyDaroch
北京大学学报(自然科学版) 2023年1期
肖萌 蒋莹 崔逸璇 Sadaf Riaz Maurycy Daroch
北京大学学报(自然科学版) 第59卷 第1期 2023年1月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 59, No. 1 (Jan. 2023)
10.13209/j.0479-8023.2022.100
深圳市基础研究项目(GXWD20201231165807007-20200806170221001)资助
2022-02-10;
2022-02-24
重组大肠杆菌利用醋酸及乳酸为碳源合成 PHB
肖萌 蒋莹 崔逸璇 Sadaf Riaz Maurycy Daroch†
北京大学深圳研究生院环境与能源学院, 深圳 518055; † 通信作者, E-mail: m.daroch@pkusz.edu.cn
以廉价的工业副产物醋酸及乳酸为碳源, 对产 PHB 重组大肠杆菌进行培养, 考察醋酸及乳酸的添加对重组大肠杆菌生长及 PHB 产量的影响。将来自的 PHB 合成操纵子 phaCAB 基因簇克隆至 pBAD 载体, 得到产 PHB 菌株, 以阿拉伯糖为诱导剂, 在大肠杆菌中进行重组表达。分别使用 LB 及 M9 培养基, 对重组菌株进行培养, 研究其生长速度及 PHB 产量, 探索产 PHB 重组大肠杆菌最适培养基。以添加 0.04g/L 乳酸、1.2g/L 乳酸、0.02g/L 醋酸、0.6g/L 醋酸、0.04g/L乳酸+0.02g/L 醋酸、1.2g/L 乳酸+0.4g/L 醋酸的 M9 培养基(均含 2g/L 葡萄糖)为实验组, 以 M9 培养基(含 2g/L 葡萄糖)为对照组, 考察醋酸及乳酸的添加对重组大肠杆菌生长及 PHB 产量的影响。分别取第 6, 12, 24和 36 小时的培养液, 分析其葡萄糖、醋酸及乳酸含量的变化。结果表明, 低氮型 M9 培养基更适合产 PHB 重组大肠杆菌在低糖培养环境中生长。在葡萄糖消耗殆尽后, 大肠杆菌能够以醋酸及乳酸为碳源进行代谢, 因此在培养基中添加一定浓度的醋酸及乳酸能够有效地提高重组菌株产 PHB 能力, 在乳酸添加量为 1.2 g/L 时, PHB 产量达到最高(1.43 g/L), 比对照组提高 78%。
PHB; 乳酸; 醋酸; 大肠杆菌
聚羟基丁酸酯(polyhydroxybutyrate, PHB)是一种常见的生物聚合物, 由于具有疏水性、完全的生物降解性和良好的生物相容性等优点, 广泛用做传统塑料的替代品[1]。自然界中的 PHB 往往是微生物在碳源丰富且氮、硫、磷等其他营养元素缺乏时积累的[2]。是目前研究最深入的高 PHB 产量微生物[3], 其野生型菌株的 PHB 产量可达细胞干重的 78%, 来源于的 PHB 合成基因 phaCAB 广泛用于重组大肠杆菌菌株中生产PHB。Napathorn 等[4]的研究表明, 通过菌株优化、基因工程及发酵条件优化等手段, 大肠杆菌发酵产PHB 的产量可高达大肠杆菌细胞干重的 93.3%。与野生型 PHB 生产菌株相比, 重组大肠杆菌产 PHB具有许多优势, 如大肠杆菌培养周期短, 可利用的碳源广泛, PHB 产量相对较高, 并且由于缺乏解聚酶而在发酵过程中不会降解 PHB[5]。
重组大肠杆菌生产 PHB 往往以葡萄糖为碳源, 具有一定的经济局限性, 因此选用廉价的碳源进行PHB 的生物合成可有效地降低成本。醋酸是廉价易得的工业副产物, 在许多工业过程中都有生成(如石油副产物提取和木质纤维素的水解工艺等[6]), 目前醋酸的市场价格远低于葡萄糖[7]。在低葡萄糖条件下, 大肠杆菌内的乙酰辅酶 A 合成酶()相关基因代谢调控能帮助大肠杆菌对培养环境中的醋酸进行利用, 生成大肠杆菌代谢的重要前体物质乙酰辅酶 A[8]。乳酸通常是利用工业、农业及食品废弃物进行生物发酵而得, 其生产成本低且来源广泛[9]。当培养环境中乳酸含量较高时, 细菌体内的 D-乳酸脱氢酶()可发生逆催化反应, 催化乳酸生成丙酮酸, 参与细菌代谢[10]。因此, 使用醋酸及乳酸作为产 PHB 重组大肠杆菌的碳源, 是低成本生产PHB 的有效方法。此外, 传统重组大肠杆菌生产PHB 的研究常使用 LB 培养基作为培养介质。LB培养基中的胰蛋白胨组分富含大量氮源, 且 LB 培养基不直接含葡萄糖, 不利于 PHB 的合成。这是由于在低氮条件下, 大肠杆菌的 TCA 循环会被抑制, 从而使得乙酰辅酶 A 增加[11], 乙酰辅酶 A 是大肠杆菌合成 PHB 的唯一前体。高含量的乙酰辅酶 A 有利于 PHB 的产生, 因此选择低氮含量的培养基可以提高重组大肠杆菌的产 PHB 能力。LB 培养基中的主要碳源供体为酵母提取物, 其碳源通常以可分解代谢的氨基酸的形式存在[12], 与氨基酸相比, 醋酸及乳酸更接近大肠杆菌合成 PHB 的代谢通路, 理论上以醋酸及乳酸为碳源更利于产物 PHB 的合成。
本文将来自的 PHB 合成操纵子 phaCAB 基因簇克隆至 pBAD 载体, 以廉价的阿拉伯糖为诱导剂, 在大肠杆菌中进行重组表达, 探究不同培养基对重组菌株产 PHB 的影响, 同时探究重组菌株在低糖且含不同浓度乳酸及醋酸条件下的生长情况及 PHB 产量, 为低成本、绿色生产 PHB的工程化应用提供理论基础。
1 材料和方法
1.1 实验材料
1)菌株和质粒: 克隆菌株采用DH5α, 表达菌株采用BL21(DE3), 采用非连接式克隆 pBAD LIC cloning vector (8A)载体, 来自的 PHB 合成操纵子phaCAB 基因簇经密码子优化后, 由华大基因公司合成。
2)主要试剂: 限制性核酸内切酶购于Thermo 公司, pfu 高保聚合酶、Taq DNA 聚合酶、非连接酶依赖型单片段快速克隆试剂盒、DNA 凝胶回收试剂盒及质粒小量提取试剂盒购于 Vazyme公司。
3)培养基: LB 培养基的配制参照 Lin 等[13]的方法。M9 培养基粉末购于山东拓普生物工程有限公司。终浓度为 2g/L 葡萄糖的培养基: 配 1g/的葡萄糖母液进行过滤灭菌后, 以 1:500 的体积比添加至经高压灭菌的培养基中。
1.2 实验方法
1.2.1 产 PHB 重组大肠杆菌的构建
图 1 展示预期的重组大肠杆菌代谢路线。本研究使用的菌株及质粒及引物列于表 1。将 pBAD 质粒通过内切酶进行酶切, 得到线性质粒。使用带同源臂的引物 phaCAB_F 及 phaCAB_R 对 pha CAB 操纵子进行 PCR 扩增, 通过快速克隆试剂盒, 将所得带同源臂的 phaCAB 基因簇(3693bp)与线性pBAD 载体进行连接, 然后转化至大肠杆菌 DH5α感受态, 进行重组质粒复制。质粒测序由广州艾基生物公司完成。将测序正确的质粒转入大肠杆菌BL21(DE3)感受态细胞, 得到重组菌株。图 2 为质粒构建示意图。
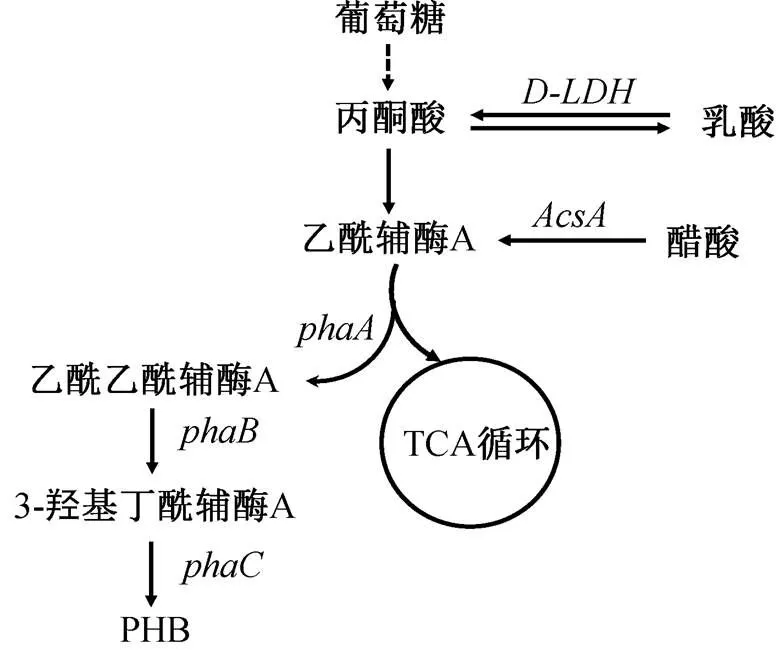
D-LDH: D-乳酸脱氢酶; AcsA:乙酰辅酶 A 合成酶; phaA: β-酮硫裂解酶; phaB:乙酰基乙酰辅酶 A 还原酶; phaC:PHB 聚合酶
1.2.2 重组菌株__的蛋白表达
将重组菌株在 LB 培养基中进行过夜培养, 然后以 2% (体积比)的接种量接种于含 50ng/μL 氨苄青霉素抗生素的 LB 液体培养基的锥形瓶中。37°C 和 200rpm 条件下培养至 OD600约为 0.4~0.6, 加入终浓度为 1% (质量体积比)的阿拉伯糖作为诱导剂, 于 30°C 振荡表达 12 小时。培养结束后, 取 20mL 培养液, 于 4°C 和 10000r/min条件下离心 15 分钟, 收集诱导菌体, 用 2mL 预冷的 20mM Tris-HCl 缓冲液(pH8.0)重悬收集菌体, 冰浴中进行超声破碎处理(5min/样品, 每个循环运行 5 秒后暂停 9 秒)。破碎完成后, 在 4°C 和 12000r/ min 条件下离心 20 分钟, 获得含 phaC, phaA 及phaB 蛋白的上清粗蛋白酶液。使用所得上清粗酶液进行 SDS-PAGE 分析。由于 SDS-PAGE 不能清楚地确定蛋白的表达情况, 将 SDS-PAGE 凝胶切下, 送样至华大基因公司进行蛋白 Q-E 质谱鉴定。

表1 本研究所用质粒、菌株及引物
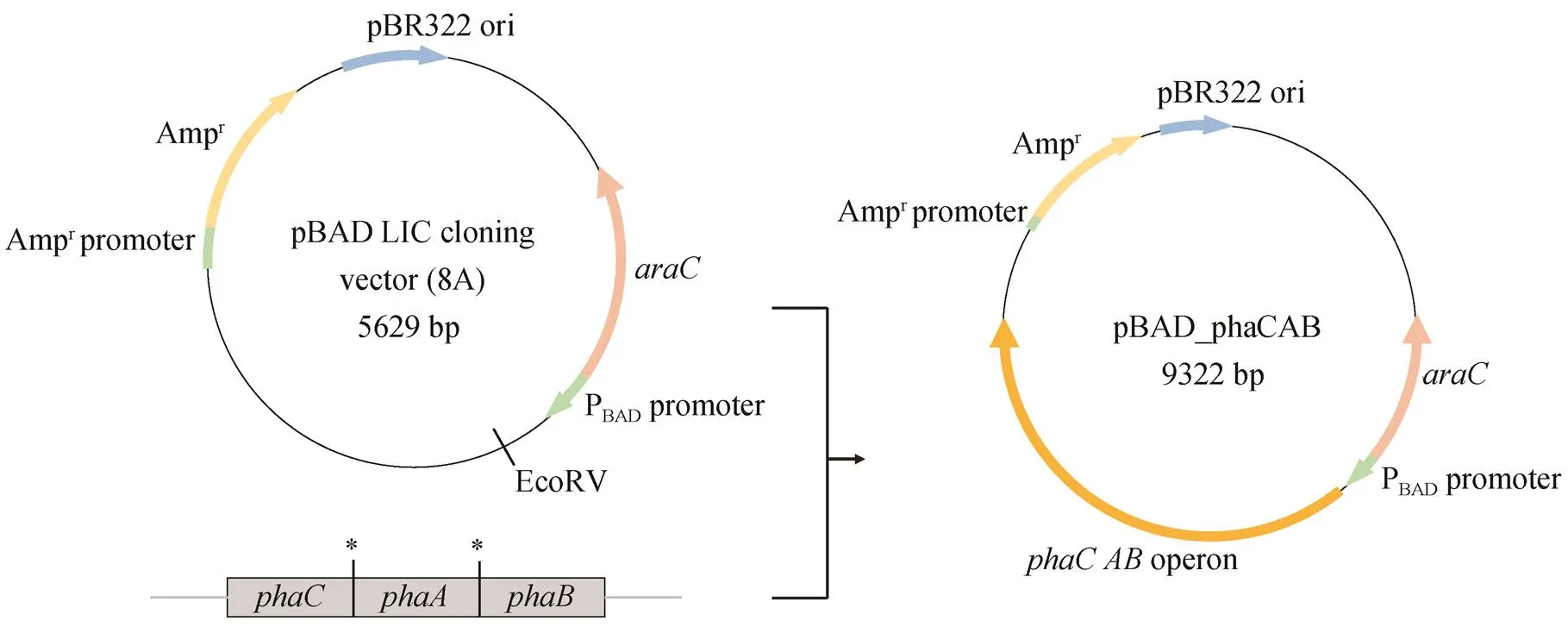
*表示终止密码子
1.2.3 PHB 的胞内形态检测
取 0.5mL 过夜的表达重组菌液, 离心分离后收集菌体, 采用 PBS 缓冲液(pH=7.0)悬浮菌体, 然后加入电镜固定液, 将样品送至中科百测公司进行透射电镜检测。
1.2.4 重组菌株的培养基优化
将重组菌株在常规 LB 培养基中于 37°C 和 200rpm 条件下过夜培养, 得到预培养液。分别采用 LB 培养基与 M9 液体培养基(培养基中均含终浓度为 2g/L 的葡萄糖)对重组菌株进行培养。以 2%的接种量, 将预培养液分别接种于含两种 50mL 液体培养基的 250mL 锥形瓶中, 在37°C 和 200rpm 条件下培养 6 小时后, 培养温度转为 30°C, 并加入终浓度为 1%的阿拉伯糖进行诱导表达。在此期间, 分别取培养 6, 12, 24 和 36 小时的菌液测定 OD600 值, 并对两种培养基中培养 36 小时后的重组菌株分别进行 PHB 含量检测。每组实验均设置 3 组平行样品。
1.2.5 添加醋酸和乳酸的影响
分别在含 2g/L 葡萄糖的 M9 培养基中加入不同起始浓度的乳酸、醋酸及二者的组合, 实验组设置为 0.04g/L 乳酸(LL)、1.2g/L 乳酸(HL)、0.02g/L 醋酸(LA)、0.6g/L 醋酸(HA)、0.04g/L 乳酸+0.02g/L醋酸(LL+LA)及1.2g/L乳酸+0.4g/L醋酸(HL+HA), 以含 2g/L 葡萄糖的常规 M9 培养基为对照组。按照1.2.4 节所述方式, 对重组菌株 B 进行培养, 取培养6, 12, 24 和 36 小时的菌液进行 OD600值的测定, 并测定培养基中葡萄糖、乳酸及醋酸的含量, 最终对培养 36 小时后的重组菌株 PHB 含量及产量进行测定。每组实验均设置 3 组平行样品。
1.2.6 重组菌株 PHB 的产量测定
通过 GC-MS 对 PHB 进行定量分析[14–15], 以PHB 质量/细胞干重作为 PHB 的含量, 以 1L 重组菌株培养液所含总 PHB 质量作为产量。在 GC-MS 检测之前, 需要对 PHB 进行衍生化反应[16], 操作步骤如下: 取表达完成的菌液进行离心分离后, 收集菌体, 冷冻干燥 48 小时, 称取25mg 干燥菌体, 置于高压反应管中, 依次加入 2mL 氯仿、2mL 和 7%甲醇–硫酸溶液及 200μL 的 3%甲醇–苯甲酸溶液, 在 105°C 条件下反应 8 小时, 期间每 2 小时取出, 均匀摇晃 30 秒。反应完成后, 向反应液中加入 2mL 去离子水, 充分振荡摇匀。此过程中, 利用甲醇在氯仿与水中的溶解度不同, 将甲醇等杂质萃取到水相中, 静置 1 小时后, 上层为水相, 下层为氯仿层, 取下层进行 GC-MS 检测。PHB的 GC-MS 色谱测定条件如下: 采用安捷伦气质联用仪 8890-5977B; 采用安捷伦公司 HP-5MS (30m× 0.25mm×0.25μm)毛细色谱柱; 载气为氦气, 1mL/ min 恒流模式; 进样口温度为 250°C; 采用不分流模式。升温程序: 初始温度为 80°C, 保持 1 分钟, 以10°C/min 的速率升温至 140°C, 保持 2 分钟, 再以4°C/min 的速率升温至 250°C, 保持 8 分钟。
1.2.7 葡萄糖、乳酸及醋酸的检测
重组菌株生长过程中的葡萄糖消耗量、乳酸及醋酸的变化量由 HPLC 进行检测[17–18]。取 1mL 重组菌株培养液, 离心分离后取上清液, 过滤后进行 HPLC 检测。高效液相色谱测定条件如下: 采用安捷伦高效液相色谱仪 1271, 安捷伦 Hi-plex H (4.6×250 mm)色谱柱, 柱温 60°C, 流动相为 5mM 硫酸溶液, 流速 0.6mL/min。葡萄糖采用 RID 检测器检测, 醋酸及乳酸采用 DAD 检测器进行检测, 检测波长为 210 nm。
2 结果
2.1 重组菌株 BL21_pBAD_phaCAB 的蛋白表达
重组菌株在 1%阿拉伯糖诱导下的蛋白表达情况如图 3 所示。根据预测, 重组蛋白的 phaC 理论大小为 60kDa, phaA 理论大小为 40kDa, phaB 理论大小为 26kDa。通过 SDS-PAGE 结果可以看出, 与未诱导表达的重组细胞上清粗酶液相比, 诱导表达后的细胞上清粗酶液拥有一个明显的 phaB 归属条带, 但 phaC 和 phaA 的表达情况难以通过 SDS-PAGE 结果判断。从理论上讲, phaCAB 操纵子均受同一启动子的调控, 且其基因片段排列顺序依次为 phaC, phaA 和 phaB, 由于 phaB 处于下游, 一定是在 phaC和 phaA 之后表达。因此, 猜测细胞上清粗酶液中存在 phaC 和 phaA 目的蛋白, 但浓度较低, 无法从SDS-PAGE 结果中看出。如图 3 所示, 将猜测归属于 phaC, phaA 和 phaB 的 SDS-PAGE 条带(接近其大小范围的条带)分别切下后, 进行蛋白质谱 Q-E 鉴定。检测结果(表 2)显示, 对于 3 种目标蛋白, 其蛋白覆盖率分别为 phaC 15%, phaA 62%和 phaB 73%。且每一个所检测样品均能匹配到原始蛋白序列中的特有多肽片段。检测结果清晰地表明, 3 种目的蛋白均成功表达。
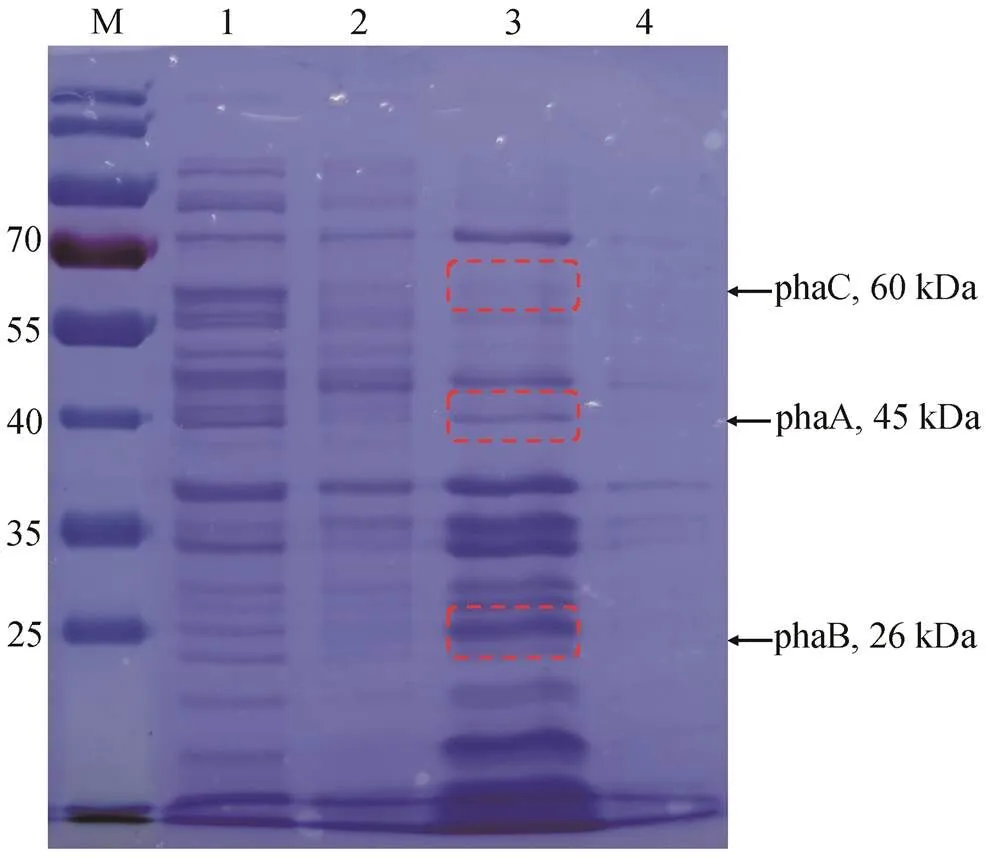
M:蛋白标准; 1:未诱导重组菌株破壁细胞上清; 2:未诱导重组菌株破壁细胞下沉; 3:诱导重组菌株破壁细胞上清; 4:诱导重组菌株破壁细胞下沉
2.2 重组菌株 BL21_pBAD_phaCAB 产 PHB的透射电镜分析
PHB 在大肠杆菌中的积累是以胞内颗粒的形式存在的[19]。重组大肠杆菌经诱导表达后的透射电镜结果如图 4 所示。样品采用包埋切片的方式进行处理, 通过图片可清晰看见重组大肠杆菌胞内存在密集的球形颗粒, 该胞内颗粒即为 PHB。这一结果证明, 重组蛋白在大肠杆菌中实现了功能化。通过图 4(a)可以看到, 重组菌株所产 PHB 粒径最大可达 200 nm, 整体而言, PHB 粒径分布于 50~200nm 之间。根据图 4(b)来看, PHB 颗粒主要以聚集而非分散的分布形式存在于大肠杆菌胞内, 占重组细胞横切面面积的 30%以上。图 4(c)和4(d)为野生型大肠杆菌 BL21 的细胞形态, 可以清楚地看到细胞内部无明显聚集颗粒, 进一步证明了重组菌株__的功能性。
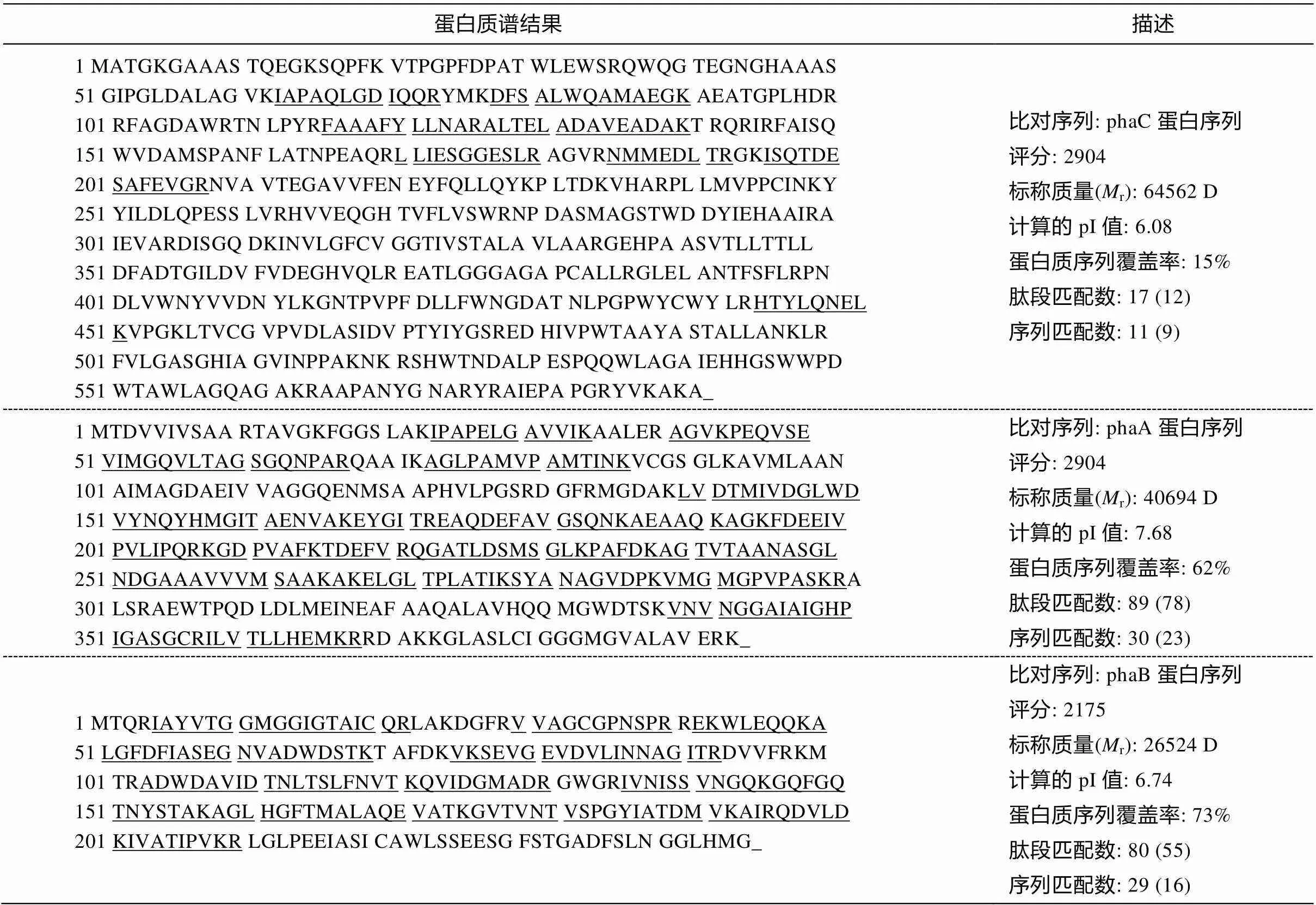
表2 重组菌株BL21_pBAD_phaCAB诱导表达蛋白phaC, phaA和phaB的质谱分析
说明: 评分的计算公式为‒10×Log, 其中是随机事件的概率;r为参考蛋白质质量; 计算得出的 pI 值表示参考蛋白质等电点的理论值; 下划线标记的肽序列表示与参考蛋白质为同源序列。
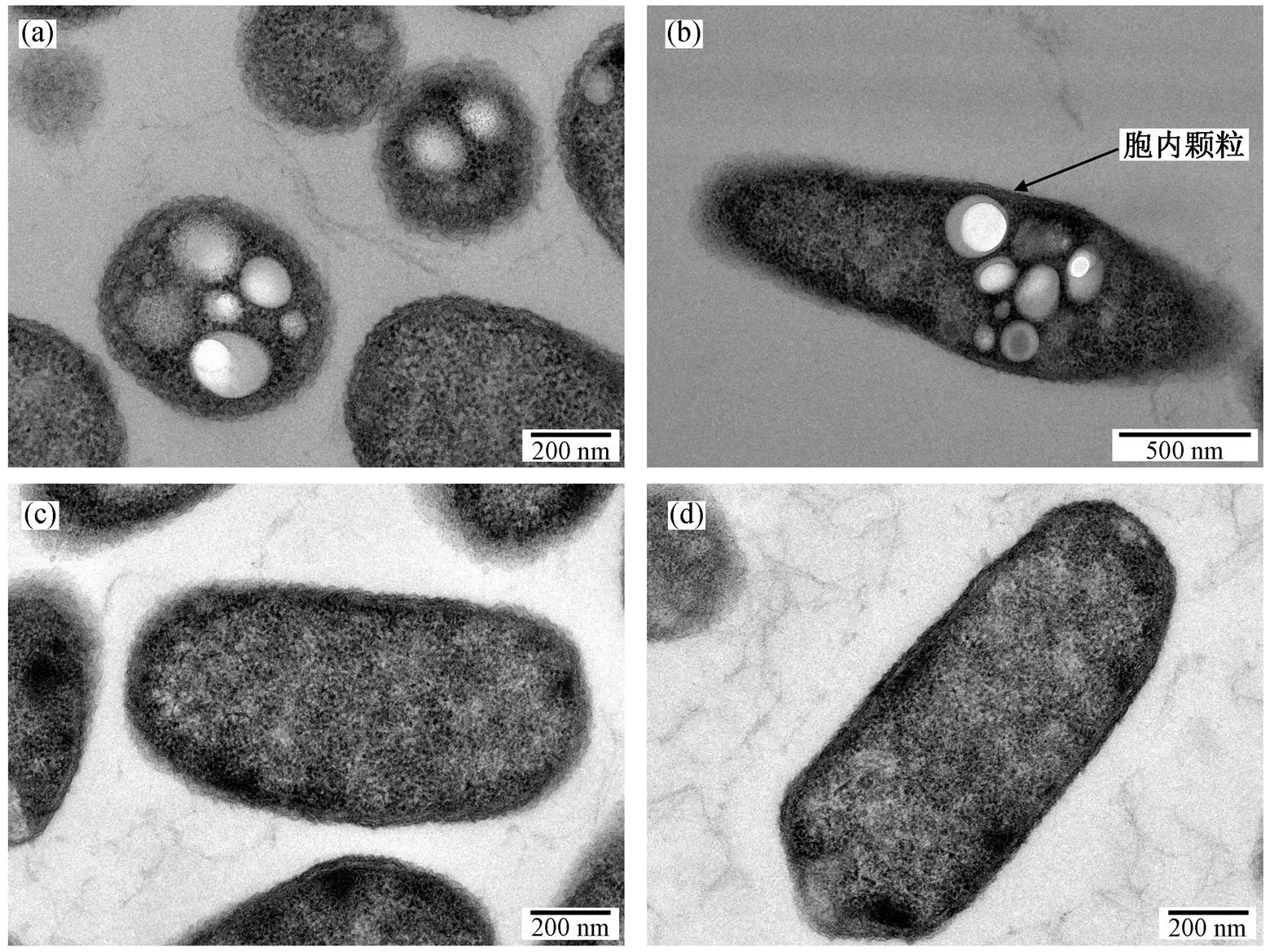
(a)~(b):重组菌株BL21_pBAD_phaCA的胞内形态; (c)~(d):野生型大肠杆菌菌株BL21的胞内形态
2.3 培养基对重组菌株 BL21_pBAD_phaCAB生长及 PHB 含量的影响
图 5 比较重组菌株在 LB 和 M9 培养基中(均含2 g/L 葡萄糖)的生长曲线及其 PHB 终含量。如图5(a)所示, 在 LB 培养基中培养的重组菌株在 6~12小时处于生长对数期, 能够以较快的速度生长至OD600 大于 1, 12 小时后细胞趋于稳定生长, OD600值最终稳定在 1.1 左右。在 M9 培养基中培养的重组菌株, 12 小时前的生长速度与 LB 培养基中的细胞相当, 但对数生长期持续约 18 小时, 细胞 OD600 值能够达到 2.6, 远大于 LB 培养基中的细胞密度。如图 5(b)所示, 以 LB 为培养基培养的重组菌株的 PHB 终产量为 11.3%, 远低于以 M9 为培养基的重组菌株 PHB 产量29.3%。
2.4 醋酸和乳酸对重组菌株 BL21_pBAD_phaCAB产 PHB 的影响
图 6 展示重组菌株在含不同起始浓度醋酸及乳酸的 M9 培养基(含 2g/L 葡萄糖)中的生长曲线及 PHB 终含量。如图 6(a)所示, 在高含量乳酸(HL)的培养基中, 重组菌株在前 20 小时的生长速度比其他条件下更快; 在高起始乳酸及醋酸含量(HL+HA)的培养基中, 细胞能够在 24 小时内达到最高的稳定 OD 值, 约为 3.3, 高于对照组的2.6; 在添加高浓度醋酸(HA)的实验组中, 细胞生长速度及最高稳定 OD 值均大于对照组。如图 6(b)所示, 在高含量乳酸(HL)及高含量醋酸(HA)中生长的重组细胞能够获得分别为 42%和 40.8%的 PHB 含量, 远高于对照组的 29.7%。然而, 生长速度最快且终 OD 值最高的 HL+HA 实验组获得较低的 PHB终含量, 仅为 29.9%。此外, 终 OD 值小于对照组的 LA 实验组获得 29.4%的 PHB 终含量, 也是唯一一组 PHB 终含量小于对照组的实验组。表 3 总结各种条件下的 PHB 终含量、产量及产率, 通过计算得出, 在 HL 条件下可获得的 PHB 产量为1.43g/L, 即36 小时内的产率为 0.040g/(L∙h), 比对照组的产量(0.8g/L)提高 78%。
2.5 培养基中的葡萄糖、乳酸及醋酸的含量变化
本研究在利用含不同起始浓度的醋酸及乳酸的低糖 M9 培养基(含 2g/L 葡萄糖)对重组菌株进行培养期间, 分别于第 6, 12, 24和 36 小时对培养基中的葡萄糖、乳酸及醋酸含量进行测定, 观察细胞对 3 种物质的代谢情况, 图 7显示实验组 HL, HA, HL+HA 及对照组在 0~36 小时中的葡萄糖、乳酸及醋酸含量变化。可以看出, 重组细胞在这 4 种条件下的葡萄糖消耗均有相同的趋势: 在前 6 小时大量消耗葡萄糖, 6 小时以后培养基中的葡萄糖含量极低, 6~12 小时期间细胞消耗葡萄糖速度放缓, 直到第 36 小时, 培养基中几乎无葡萄糖残余。此外, 前 12 小时, 4 种条件下的醋酸含量均呈增加趋势, 直到第 12 小时, 培养基中的醋酸含量达到约 0.6g/L, 而 12 小时以后, 在葡萄糖含量极低的状态下, 醋酸被细胞作为碳源参与代谢, 含量逐渐降低, 直到第 36 小时, 培养基中几乎无醋酸残留。在不添加乳酸的培养条件下, 重组细胞在生长的前 6 小时均有乳酸的分泌, 终浓度达到约 0.4g/L, 而细胞从第 6 小时就开始缓慢消耗培养基中的乳酸, 直到第 36 小时培养基中的乳酸含量接近 0。值得注意的是, 初始乳酸添加量相同的 HL 及HL+HA 实验组的乳酸含量变化趋势有所不同。HL 实验组中, 乳酸含量始终呈现逐渐降低的趋势, 而 HL+HA 实验组中, 前 6 小时, 重组细胞在含高起始浓度乳酸的培养基中仍然呈现乳酸含量增加的趋势, 表明重组细胞在此条件下的乳酸消耗量小于生成量。
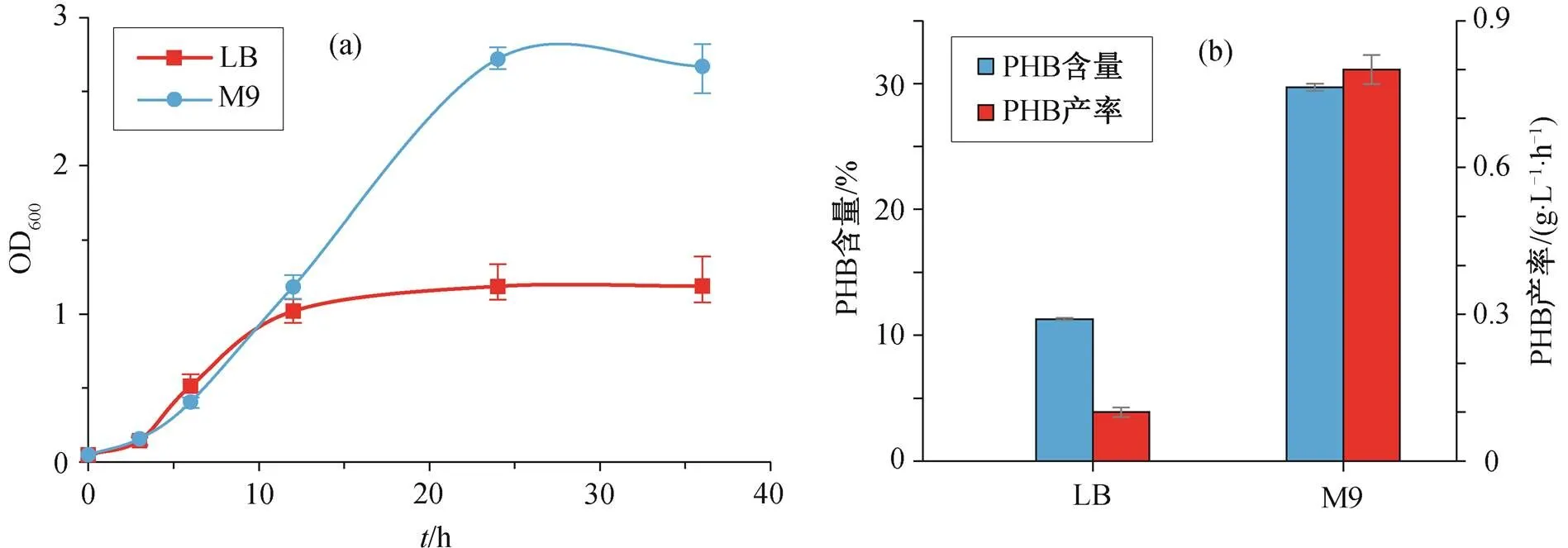
图5 重组菌株BL21_pBAD_phaCAB在LB和M9培养基中的生长曲线及PHB终含量
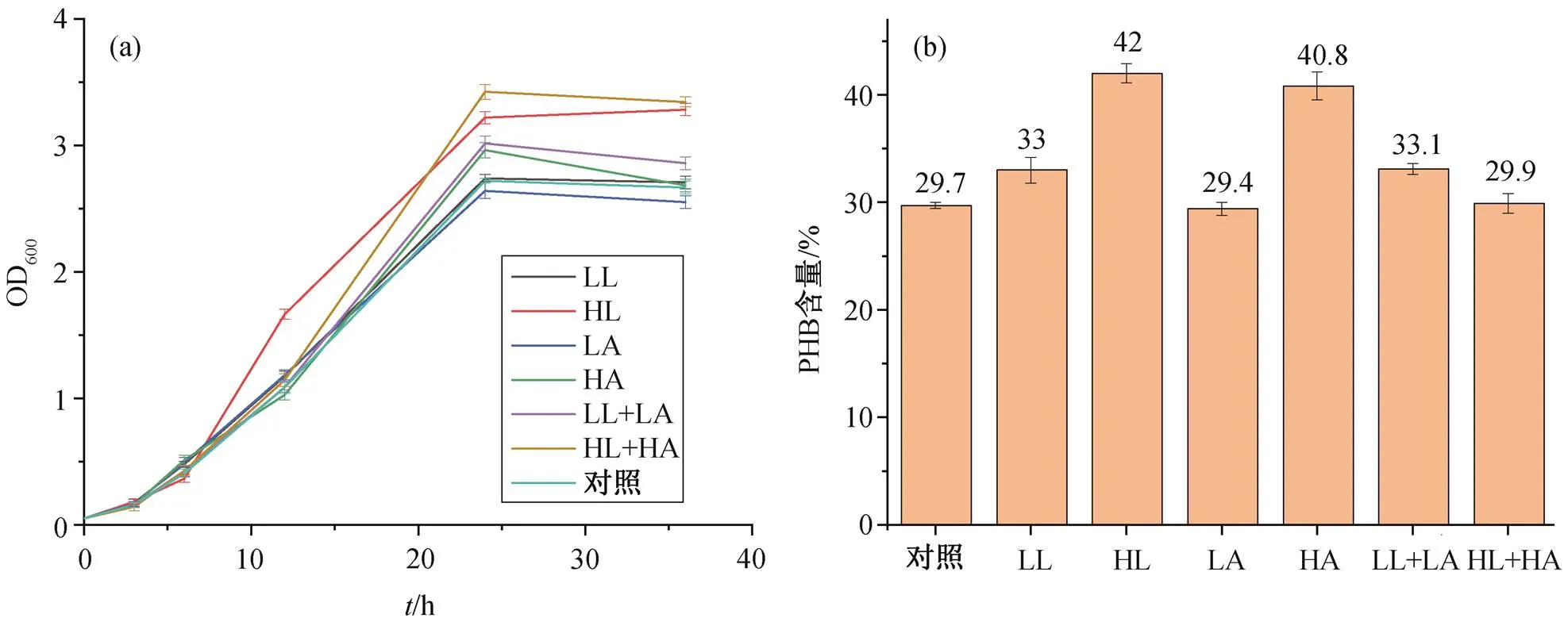
LL: 0.04 g/L乳酸; HL: 1.2 g/L乳酸; LA: 0.02 g/L醋酸; HA: 0.6 g/L醋酸; LL+LA: 0.04 g/L乳酸+0.02 g/L醋酸; HL+HA: 1.2 g/L乳酸+0.4 g/L醋酸; 对照组: 无醋酸及乳酸添加, 下同
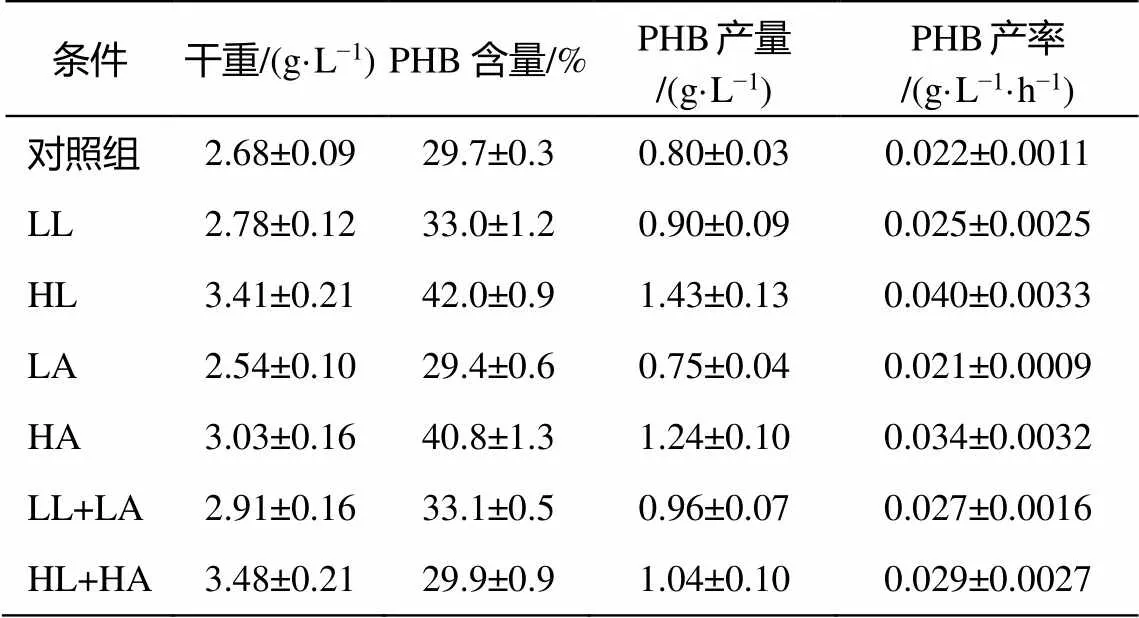
表3 重组菌株BL21_pBAD_phaCAB在不同乳酸及醋酸条件下的PHB终含量、产量及产率
说明: CDW 为细胞干重, 即 1L 菌液所含菌体在充分干燥后的质量; PHB 含量 = PHB 质量(g)/细胞干重(g); PHB 产量 = 细胞干重(g/L)×PHB 含量; PHB 产率= PHB 产量/36 (h)。
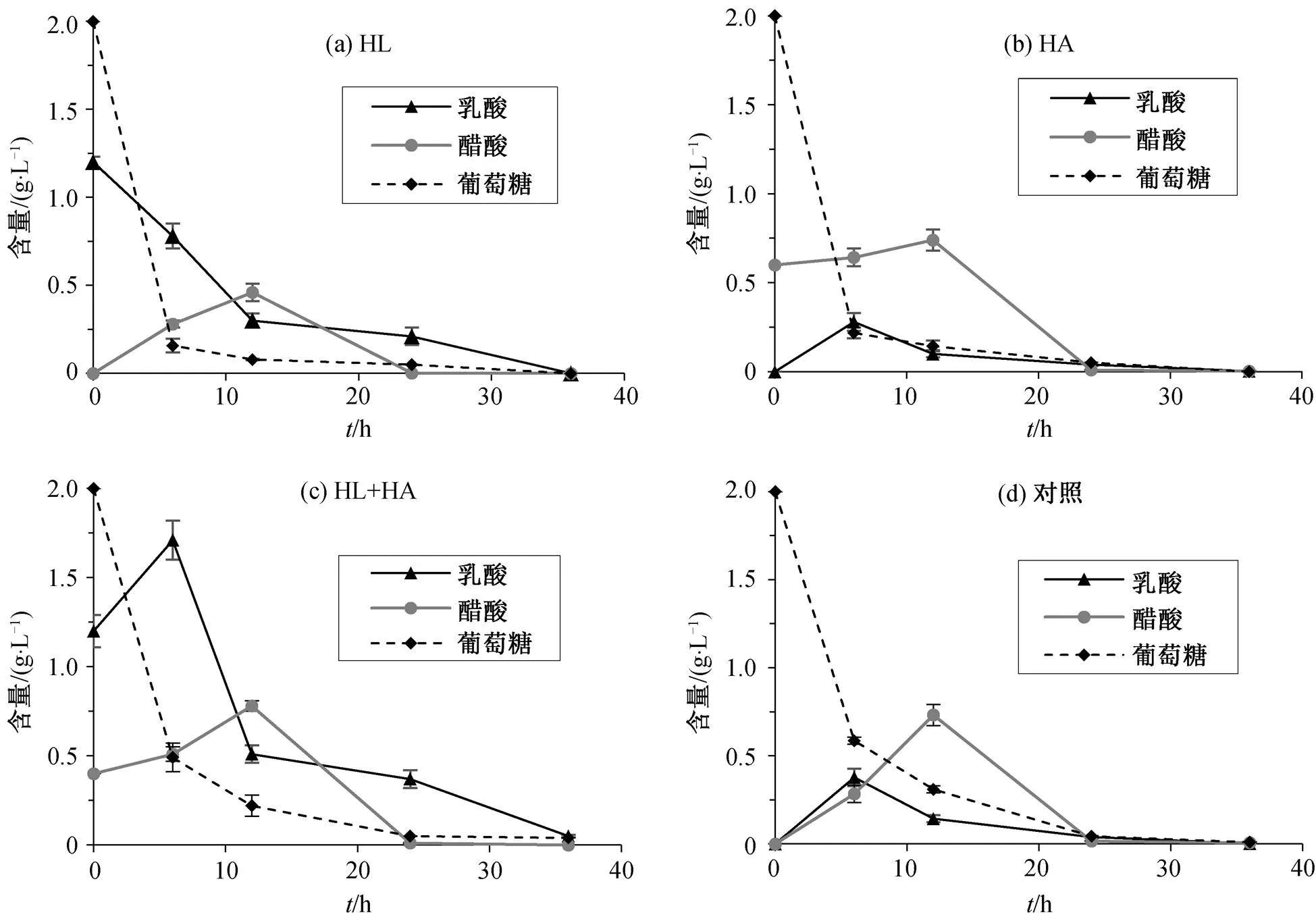
图7 培养基中葡萄糖、乳酸及醋酸含量的变化
3 讨论
通过蛋白质谱及透射电镜结果, 我们确定了重组菌株的功能性。通过对比产 PHB 重组菌株在不同培养基中的生长情况, 我们发现常见的 LB 培养基并不是培养产 PHB 重组大肠杆菌的最佳选择。这是因为 LB 培养基中氮元素过多, 且不直接含葡萄糖, 氮源缺乏型 M9 培养基更符合产 PHB 重组大肠杆菌的生长条件。在添加 2 g/L 葡萄糖的情况下, M9 培养基中培养的产 PHB 重组菌株中的 PHB 含量接近 LB 培养基中的 3 倍, 为细胞干重的 29.7%, 其 PHB 产量为 LB 培养基中的 8倍, 为 0.7g/L, 此时的 PHB 含量受限于低糖的条件。Sathish 等[20]将 20g/L 的葡萄糖添加至 M9 培养基中, 在 37°C 条件下对产 PHB 重组大肠杆菌培养48 小时, 得到细胞干重 62%的 PHB 含量。
通常认为, 醋酸和乳酸是对大肠杆菌生长有害的物质[21], 但本研究发现, 浓度适当的醋酸及乳酸可作为产 PHB 重组菌株的有效碳源。在细胞生长悬浮期及细胞生长对数期的早中期, 细胞快速消耗大量葡萄糖用于自身代谢, 此阶段醋酸总量有明显的积累。这是由于在葡萄糖水平高于细胞生长所需限制水平时, 细胞代谢能够自主分泌醋酸与乳酸[22]。值得注意的是, 在细胞悬浮期, 大多数情况下乳酸处于积累状态, 唯一例外的是在仅添加高浓度乳酸的培养基中, 乳酸含量在监测时间点均处于减少状态, 说明细胞从培养早期便开始消耗乳酸进行自身代谢。结合葡萄糖消耗曲线及生长曲线可以推测, 这是由于细胞在此条件下生在速度极快, 培养基中葡萄糖消耗速度较快, 葡萄糖水平很快跌到细胞生长所需限制水平, 从而抑制乳酸的产生, 而较高的乳酸起始浓度激活了 D-乳酸脱氢酶的逆向催化, 促使细胞利用乳酸合成丙酮酸[23]。此外, 我们发现对于同时添加高浓度乳酸及醋酸的细胞, 其生长速度及最终的稳定 OD 值均高于其他条件, 但其细胞 PHB 含量与对照组相差无几, 说明在此条件下细胞更多地利用碳源进行自身生长及繁殖, 因此细胞中 PHB 含量较低。
本文研究结果表明, 当培养环境中的葡萄糖消耗殆尽时, 细胞可利用培养基中的醋酸及乳酸来维持自身代谢, 直到培养基中乳酸及醋酸均消耗殆尽, 细胞进入生长衰亡期。在添加 1.2g/L 乳酸的培养基中, 产 PHB 重组菌株生长, 能够获得高达 42%的 PHB 含量, 比不添加乳酸的培养基中培养的细胞 PHB 含量高约 41%。然而, 过高浓度的醋酸及乳酸添加会抑制细胞生长, 在乳酸浓度大于 1.2g/L 及醋酸浓度大于 0.6g/L 的生长条件下, 重组菌株的生长速度明显减慢, 终 OD 值明显降低, 且在乳酸浓度 1.2g/L 及醋酸浓度 0.6g/L 以内范围, 有机酸的添加不会对培养基整体 pH 造成明显的改变(培养基 pH 始终在 6.8~7.0 之间)。Sun 等[24]在产PHB 重组大肠杆菌中过表达基因, 使得大肠杆菌可以在含 3g/L 醋酸及 1g/L 葡萄糖的培养基中生长, 得到最终 PHB 产量约为 0.12g/L。但是, 目前仍然没有在大肠杆菌中过表达 D-乳酸脱氢酶产PHB 的报道, 未来可尝试利用此方法提高菌株对乳酸的耐受能力。
4 结论
本研究将来自的 PHB 合成操纵子 phaCAB 基因簇在大肠杆菌中进行重组表达, 获得产 PHB 的重组大肠杆菌。通过用不同的培养基对重组菌株进行培养, 选出更适合产 PHB 重组大肠杆菌的 M9 营养缺乏型培养基。通过向培养基中添加不同浓度的醋酸及乳酸, 探究产 PHB 重组大肠杆菌的生长情况、PHB 含量及产量以及生长过程中对葡萄糖、醋酸及乳酸的消耗情况, 得出以下结论: 1)在低糖环境中, 添加适量醋酸及乳酸能够提高重组菌株产 PHB 能力; 2)细胞倾向于在葡萄糖消耗殆尽后开始利用醋酸进行代谢, 而较高含量的乳酸添加可以在细胞生长初期激活细胞消耗乳酸的能力。
综上所述, 利用比葡萄糖更为廉价的醋酸及乳酸为碳源, 对产 PHB 重组大肠杆菌进行培养是极具经济潜力的手段, 可大大降低 PHB 的生产成本。未来可利用基因工程技术对其进行基因改造, 提高对乳酸及醋酸的耐受力及利用率, 进一步提高其市场化价值。
[1]Madison L, Huisman G W. Metabolic engineering of poly(3-hydroxyalkanoates): from DNA to plastic. Mic-robiology and Molecular Biology Reviews, 1999, 63 (1): 21–53
[2]Al-Haj Ibrahim H, et al. Bio-energy production from rice straw a review. Recent Adv Petrochem Sci, 2018, 5(5): 555671
[3]Vandamme P, Coenye T. Taxonomy of the genus Cupriavidus: a tale of lost and found. International Journal of Systematic & Evolutionary Microbiology, 2004, 54(6): 2285–2289
[4]Napathorn S C, Visetkoop S, Pinyakong O, et al. Poly-hydroxybutyrate (PHB) production using an arabinose- inducible expression system in comparison with cold shock inducible expression system in. Frontiers in Bioengineering and Biotechnology, 2021, 9: 661096
[5]Lee S.moves into the plastic age. Nature Bio-technology, 1997, 15(1): 17–18
[6]Leif J, Carlos M. Pretreatment of lignocellulose: for-mation of inhibitory by-products and strategies for mi-nimizing their effects. Bioresource Technology, 2016, 199: 103–112
[7]Lim H G, Lee J H, Noh M H, et al. Rediscovering acetate metabolism: its potential sources and utiliza-tion for bio-based transformation into value-added chemicals. Journal of Agricultural and Food Chemis-try, 2018, 66(16): 3998–4006
[8]Enjalbert B, Millard P, Dinclaux M, et al. Acetate fluxes inare determined by the ther-modynamic control of the Pta-AckA pathway. Scien-tific Reports, 2017, 7: 42135
[9]John R P, Nampoothiri K M, Pandey A. Fermentative production of lactic acid from biomass: an overview on process developments and future perspectives. Appl Microbiol Biotechnol, 2007, 74(3): 524–534
[10]Nessler S, Bras G L, Bras G L, et al. Crystallization of d-lactate dehydrogenase from Lactobacillus bulga-ricus. Journal of Molecular Biology, 1994, 235(1): 370–371
[11]Wang Qian, Yu Hongmin, Xia Yongzhen, et al. Com-plete PHB mobilization inenhances the stress tolerance: a potential biotechnological app-lication. Microbial Cell Factories, 2009, 8(1): 1–9
[12]Sezonov G, Joseleau-Petit D, d’Ari R.physiology in Luria-Bertani broth. Journal of Bacteriology, 2007, 189(23): 8746–8749
[13]Lin Jihong, Lee M C, Sue Yousheng, et al. Cloning of phaCAB genes from thermophilicinfor poly(3-hydrox-ybutyrate) (PHB) production. Applied Microbiology and Biotechnology, 2017, 101(16): 6419–6430
[14]Lageveen R G, Huisman G W, Preusting H, et al. Formation of polyesters by pseudomonas oleovorans: effect of substrates on formation and composition of poly-(R)-3-hydroxyalkanoates and poly-(R)-3-hydro-xyalkenoates. Applied & Environmental Microbiolo-gy, 1988, 54(12): 2924–2932
[15]Roh H, Lee J S, Choi H I, et al. Improved CO2-derived polyhydroxybutyrate (PHB) production by engineering fast-growing cyanobacteriumUTEX 2973 for potential utilization of flue gas. Bioresource Technology, 2021, 327: 124789
[16]Mai V R. Gas chromatographic determination of poly--hydroxybutyric acid in microbial biomass after hy-drochloric acid propanolysis. Journal of Chromato-graphy A, 1988, 445: 285–289
[17]Palmer J K, Brandes W B. Determination of sucrose, glucose, and fructose by liquid chromatography. Jour-nal of Agricultural & Food Chemistry, 1974, 22(4): 709–712
[18]Özcelik S, Kuley E, Özogul F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. LWT, 2016, 73: 536–542
[19]Zhang Xingchen, Guo Yingying, Liu xu, et al. Engi-neering cell wall synthesis mechanism for enhanced PHB accumulation in. Metabolic Engineering, 2018, 45: 32–42
[20]Sathish A, Glaittli K, Sims R C, et al. Algae biomass based media for poly(3-hydroxybutyrate) (PHB) pro-duction by. Journal of Polymers and the Environment, 2014, 22(2): 272–277
[21]Shi Lihong, Da Yangyang, Zheng Weitao, et al. Pro-duction of polyhydroxyalkanoate from acetate by metabolically engineered. Jour-nal of Bioscience and Bioengineering, 2020, 130(3): 290–294
[22]Neijssel O M, Teixeira de Mattos M J, Tempest D W. Growth yield and energy distribution // Neidhart F C.&, Cellular & Molecular Biology. Washington: ASM Press, 1996: 1683–1693
[23]Kochhar S, Hunziker P E, Leongmorgenthaler P, et al. Primary structure, physicochemical properties, and chemical modification of NAD(+)-dependent D-lactic acid dehydrogenase: evidence for the presence of Arg-235, His-303, Tyr-101, and Trp-19 at or near the active site. Journal of Biological Chemistry, 1992, 267(12): 8499–8513
[24]Sun Shenmei, Ding Yamei, Liu Min, et al. Compari-son of glucose, acetic acid and ethanol as carbon resource for production of poly(3-hydroxybutyrate) and other acetyl-CoA derivatives. Frontiers in Bio-engineering and Biotechnology, 2020, 8: 833
Synthesis of PHB by Recombinant.Using Acetic Acid and Lactic Acid as Carbon Sources
XIAO Meng, JIANG Ying, CUI Yixuan, Sadaf Riaz, Maurycy Daroch†
School of Environment and Energy, Peking University Shenzhen Graduate School, Shenzhen 518055; † Corresponding author, E-mail: m.daroch@pkusz.edu.cn
Two cheap byproducts lactic and acetic acid were directly used as the carbon sources of polyhy-droxybutyrate (PHB)-producing recombinantto investigate the effects of acetic acid and lactic acid on the growth and PHB yield of recombinant. The PHB synthetic operon phaCAB gene cluster constructed on the basis ofgenes was cloned into pBAD vector to obtain PHB-producing recombinant strain, which was expressed inby using arabinose as an inducer. The recombinant strain was cultured using LB and M9 medium respectively, and the growth rate and PHB yield were studied to lead the exploration of optimum medium for PHB-producing recombinant. To explore the effect of lactic acid and acetic acid on the growth and PHB yield of the recombinant strain, 0.04 g/L lactic acid, 1.2 g/L lactic acid, 0.02 g/L acetic acid, 0.6 g/L acetic acid, 0.04 g/L lactic acid + 0.02 g/L acetic acid, 1.2 g/L lactic acid + 0.4 g/L acetic acid were added into M9 medium (containing 2 g/L glucose) as the experimental group where M9 medium (containing 2 g/L glucose) was the control group. The culture media of the 6, 12, 24, and 36 hours were taken respectively to explore the concentration changes in glucose, acetic acid, and lactic acid. The results show that the low nitrogen M9 medium is more suitable for the growth of PHB-producing recombinantin a low sugar culture environment. After the consumption of glucose,.can use acetic acid and lactic acid as carbon sources for metabolism, and by adding a proper amount of acetic acid and lactic acid, the PHB yield of the recombinant strain can get improved. When 1.2 g/L lactic acid is added, an up to 1.43 g/L PHB yield can be achieved which is 78% higher than the control.
PHB; lactic acid; acetic acid;
猜你喜欢
云南化工(2021年11期)2022-01-12 06:06:20
中华养生保健(2020年8期)2021-01-14 01:13:58
心肺血管病杂志(2020年3期)2021-01-14 00:42:38
中国甜菜糖业(2020年1期)2020-12-07 07:54:30
广州化工(2020年8期)2020-05-19 06:23:56
江苏卫生保健(2018年7期)2018-07-31 08:35:34
中国乳业(2016年4期)2016-11-07 09:50:27
腹腔镜外科杂志(2016年12期)2016-06-01 12:10:09
肿瘤影像学(2015年3期)2015-12-09 02:38:50
河北科技大学学报(2015年6期)2015-03-11 16:16:51
