
葱兰夜蛾雌蛾求偶行为节律观察与性信息素鉴定
2023-02-06 07:33胡可炎苑鹏宇钱沉鱼贾彩娟杨文娟刘春燕温秀军
昆虫学报 2023年12期
胡可炎, 苑鹏宇, 钱沉鱼, 贾彩娟, 杨文娟,刘春燕, 温秀军,*, 马 涛,*
(1. 华南农业大学林学与风景园林学院, 广州 510642; 2. 深圳市梧桐山风景区管理处, 深圳 518004;3. 深圳市广信建设工程有限公司, 深圳 518000; 4. 广东省森林资源保育中心, 广州 510130)
葱兰夜蛾Brithyscrini属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae),国外主要分布于印度、缅甸、斯里兰卡和日本等,国内主要分布在上海、江西、汕头、深圳等地(李新巾等, 2005)。该虫以幼虫进行危害,发生范围广,可取食寄主植物花、茎、叶等,造成其叶片糜烂,植株倒伏,影响植物生长,主要危害葱兰Zephyranthescandida、水鬼蕉Hymenocallislittoralis、文殊兰Crinumasiaticum等石蒜科植物(于炜等, 2009)。葱兰夜蛾老熟幼虫主要在寄主植物叶片之间或地表浅土层做蛹室化蛹;成虫主要在白天羽化,羽化后无需补充营养也能交尾、产卵,白天隐藏在寄主植物叶背或草地的杂草间,夜间活动(李新巾等, 2005; 涂小云和陈元生, 2013)。近年来,随着我国城市规模的快速发展,石蒜科植物在公园造景、城市绿地绿化中应用日广,葱兰夜蛾的发生范围和危害程度亦随之增加,故而该虫的危害极大影响了石蒜科植物的观赏价值。
目前防治葱兰夜蛾主要依靠化学防治和人工清理,但是喷洒化学农药容易造成环境污染,同时多次喷雾防治易造成害虫抗药性增强(朱佳虹等, 2014),且葱兰夜蛾的发生地主要集中在街道或者公园,更应选择高效低毒的绿色防控技术进行生态调控。鳞翅目昆虫性信息素因其高效专一、生态环保等优势,作为一种新型绿色防控技术,愈来愈受到广泛关注,不论是监测害虫的发生动态,还是通过大量诱捕或干扰交配直接防控害虫的危害,均可实现害虫的无公害防治,具有广阔的应用前景和生态效益(马涛等, 2018)。目前已有2 000多种害虫的信息素得到鉴定,其中全球商品化的昆虫信息素产品超过了600多个,其中以鳞翅目昆虫性信息素应用最为广泛(马涛等, 2019; 吴帆等, 2023)。鳞翅目昆虫性信息素主要成分为醇、醛、酯、酮、烷烃等化合物或衍生物, 按其化学结构可分为4种类型: TypeⅠ, TypeⅡ, TypeⅢ和Type 0,其中约75%的已知性信息素属于TypeⅠ类,结构为C10-C18直链不饱和脂肪醇、醛、酯类化合物,夜蛾科昆虫性信息素组分结构多属于Type Ⅰ类(董改娟等, 2008; Allisonetal., 2016; Yuvarajetal., 2017)。
关于葱兰夜蛾的研究主要集中于形态学特性和化学防治方法等方面(于炜等, 2009; 涂小云和陈元生, 2013),虽然此前已有日本学者对我国台湾北部地区的葱兰夜蛾性信息素组分进行了分析,但是缺乏相应的触角电位(electroantennogram, EAG)活性测试,所得到的性信息素组分是否与深圳地区的葱兰夜蛾存在地理种群差异,仍需进一步研究(Kouetal., 1991)。因此,本研究首先对深圳地区葱兰夜蛾求偶行为节律进行观察研究,旨在准确获得处女雌蛾,然后对葱兰夜蛾雌蛾的性信息素进行提取、定量及化学鉴定,进而为利用性信息素监测葱兰夜蛾的野外动态及绿色调控提供可靠的技术参考和科学基础。
1 材料与方法
1.1 供试虫源
实验虫源采自深圳湾公园(22.48°N, 114.95°E)。养虫室饲养条件具体如下:温度26~28 ℃,相对湿度70%~80%,光周期12L∶12D,其中光期6:00-18:00,暗期18:00-6:00。每天清理老叶和虫粪,更换新鲜的水鬼蕉叶片,将即将进入预蛹状态的幼虫挑出,放到铺有厚约5 cm湿润沙土的塑料盒中(长×宽×高=29 cm×24 cm×11 cm)。观察到有幼虫化蛹后,每天将沙土中当天产生的蛹移出,集中放置,并做好日期标记。成虫羽化后按性别和日龄分开饲养(从羽化到首个暗期结束为1日龄),每天饲喂浓度为10%蜂蜜水用于补充营养。
1.2 性信息素标准化合物
实验所需的性信息素标准化合物Z11-16∶Ald和E11-16∶Ald,均由中捷四方生物科技股份有限公司提供,纯度≥98%。
1.3 求偶行为观察
求偶观察实验的温度、相对湿度和光暗周期等条件与1.1节的条件一致。通过预实验获知葱兰夜蛾雄蛾无求偶行为,雌蛾求偶都在暗期内进行。因此, 将新羽化的处女雌蛾每头单独饲养。每天18:00进入暗期后,使用红光手电筒辅助观察其求偶行为(手电筒用白色纱布包裹,以减弱光照亮度)。实验设置3组重复,每组10头雌蛾,共观察30头雌蛾。每头虫单独饲养在500 mL透明塑料盒中(长×宽×高=12 cm×8 cm×7.8 cm),每隔0.5 h记录1次求偶情况,直到当天的全部雌蛾求偶结束为止。
1.4 性信息素的提取
选取正在求偶的处女雌蛾,放入-20 ℃冰箱中冷冻约3 min左右,取出轻压雌蛾腹部,然后用眼科手术剪剪下伸出的性信息素腺体,之后用滤纸吸附掉腺体表面的体液,最后将其放入2 mL的色谱进样瓶,按照每腺体10 μL正己烷的规格向2 mL色谱进样瓶中加入适量的重蒸正己烷(图1, 由于葱兰夜蛾雌蛾性信息素腺体体积较大, 加入足量的重蒸正己烷以保证将腺体完全浸没)。在室温下浸泡50 min后,用移液枪将浸泡液,转移到另一个洁净的色谱进样瓶中,使用N2浓缩至5 μL/FE [雌当量(female equivalent, FE)],密封后保存在-20 ℃冰箱中备用。

图1 葱兰夜蛾雌蛾求偶行为Fig. 1 Calling behavior of the female moth of Brithys crini箭头示性信息素腺体的放大。The arrow indicates the magnification of sex pheromone gland.
1.5 性信息素腺体提取物的气相色谱-触角电位联用仪(gas chromatography-electroantennagram detection, GC-EAD)分析
所用仪器为安捷伦公司7820A气相色谱与德国Syntech触角电位联用仪(GC-EAD),FID信号为火焰离子化检测器(flame ionization detector),不分流进样,进样量为1 μL。升温程序如下:初始温度50 ℃,保持1 min,然后以5 ℃/min的速率运行40 min,上升到250 ℃后保持2 min。
选取1日龄葱兰夜蛾雄蛾,用眼科手术剪将其触角剪下,剪取更接近端部的部分并用刀片切除触角端部约0.5 mm,然后用导电胶分别将触角两端包裹住并连接在金属电极上,而后将金属电极插入EAG仪探头中。手动进样1 μL浓缩后的性信息素腺体浸提液。分析样品的色谱柱为DB-5(30 m×250 μm×0.25 μm),出口端接Y型分流器(5181-3397, Agilent Technologies, 美国),使得气相色谱柱分离的气体成分被均匀地分成两部分,一部分进入火焰离子化检测器产生气相色谱的FID信号,另一部分与清洁气体混合后刺激触角产生触角电生理EAD信号,记录的信号经气流控制器(CS-55)连接到直流交流放大器(IDAC-2),再连接到计算机经GC-EAD软件同步记录数据。
1.6 性信息素腺体提取物的气相色谱-质谱联用仪(gas chromatography-mass spectrometry, GC-MS)分析
葱兰夜蛾性信息素腺体提取物鉴定在气相色谱-质谱联用仪7890A-5975C(Agilent Technologies, 美国)进行,色谱柱与GC-EAD所用型号相同,无分流进样,进样量1 μL。升温程序与GC-EAD分析时的设置一致。离子源温度230 ℃,电离方式为EI,电离能量70 eV,载气为He,流速1 mL/min,全扫描采集。
首先使用正己烷反复润洗进样针,吸取1 μL重蒸正己烷进样作为空白,待空白样品分析完后,吸取1 μL性信息素腺体提取物进样分析。通过上述GC-EAD图谱获知引起雄蛾触角反应的峰形和保留时间(retention time, RT),在GC-MS总离子流图中相应的保留时间处得到疑似组分的质谱图,通过仪器配套的Nist14数据库匹配与之相对应的标准化合物质谱图,从而确定性信息素疑似组分的具体化学结构。为了进一步确定性信息素疑似组分的化学结构,还要对上述匹配到的化合物及其同分异构体进行GC-MS分析。配制10 ng/μL的标准化合物溶液,进样和分析方法同上述性信息素腺体提取物一致。通过计算引起雄蛾EAG反应的性信息素疑似组分和相匹配的化合物的Kováts保留指数(Kováts retention index,KI)确定其化学结构。
1.7 性信息素活性组分滴度(含量)测定
在鉴定出葱兰夜蛾雌蛾性信息素腺体提取物活性组分的化学结构后,分别配制0.1, 0.25, 0.5, 0.75和1 ng/μL的该组分标准化合物溶液,按照1.4节中的GC-MS检测参数进行样品分析。获得各浓度标准品的峰面积数据后,制作剂量-峰面积标准曲线并建立两者之间的直线回归方程,用于计算雌蛾性信息素腺体提取物活性组分滴度。提取1日龄处女雌蛾性信息素腺体提取物的活性组分,每个重复5头,共3个重复,提取方法同1.3节。待浸提液浓缩后使用GC-MS分析后可得到活性组分的峰面积,根据剂量-峰面积标准曲线计算出该活性组分滴度。
1.8 雄蛾触角对标准化合物的剂量反应
EAG仪主要构成部分包括直流交流放大器(IDAC-2)、气流控制器(CS-55)、微动操作仪(MP-15)及触角电位探头(PRG-2)(Syntech,德国)。实验前用正己烷配制浓度为0.1, 1, 10和100 ng/μL标准化合物Z11-16∶Ald溶液,低温保存备用。选取1日龄葱兰夜蛾雄蛾,用眼科手术剪将其触角剪下,剪取更接近端部的部分并用刀片切除触角端部约0.5 mm,然后用导电胶分别将触角两端包裹住并连接在金属电极上,而后将金属电极插入EAG探头中。制作规格为0.5 cm×5 cm的滤纸条,用移液枪移取10 μL上述配制好的溶液到不同滤纸条,约15 s后待滤纸条风干放进巴氏玻璃管中。开始测试时,按照空气、正己烷、低浓度Z11-16∶Ald溶液、高浓度Z11-16∶Ald溶液的顺序刺激触角。每种成分测定每根触角3次,重复测定5根触角,取测定平均值。数据由Syntech EAG Pro软件记录并存储。
1.9 数据分析
求偶持续时间为求偶开始至结束的时间间隔,每日龄雌蛾的求偶时长等于当天发生求偶行为的雌蛾求偶持续时间的平均值,求偶率(%)=(求偶雌蛾数量/测试雌蛾总数)×100,根据得到的每组重复的求偶率计算平均值和标准误。KI计算公式如下(Marquesetal., 2000):
其中,n和N分别表示鉴定样品色谱峰前后正烷烃色谱峰具有的碳原子数,tR(n)和tR(N)分别表示鉴定样品前后相应碳原子数的正烷烃色谱峰的色谱保留时间,tR(unknown)表示待鉴定样品色谱峰的色谱保留时间。
所有数据使用Excel 2019进行统计整理和作图,通过SPSS 22.0进行方差分析,差异显著性采用单因素方差分析(one-way ANOVA)并通过最小显著性差异法(least significance difference, LSD)进行多重比较。
2 结果
2.1 葱兰夜蛾雌蛾求偶行为和节律
最早在进入暗期2 h后,能观察到葱兰夜蛾雌蛾的求偶活动。首先,雌蛾微微扇动翅,腹部第8和第9腹节的节间膜上的性信息素腺体表皮开始外翻并膨大,形成两个半球状黄色突起,持续膨胀与收缩,腹部末端的产卵器外伸(图1)。
观察到葱兰夜蛾雌蛾整个求偶过程都在暗期,从1日龄起即开始释放雌激素,最大求偶日龄为3日龄。1日龄雌蛾求偶开始时间在暗期3.0 h之后,此后求偶频次逐渐增加,在暗期5.5 h时达到最大,求偶高峰在暗期5.0-9.0 h;2日龄雌蛾最早求偶时间提前至暗期2.0 h,求偶频次在暗期4.0 h时达到最大,求偶高峰期为暗期3.5-7.0 h求偶频次和求偶率较1日龄有所降低;3日龄雌蛾在暗期2.5 h后开始求偶活动,求偶最高峰在暗期4.5 h时,求偶高峰期为暗期4.0-6.5 h时,求偶活跃程度最低(图2)。1-3日龄雌蛾求偶时长分别为(3.57±0.46), (3.85±0.36) 和(3.63±0.69) h,三者无显著性差异(P>0.05);1-3日龄雌蛾求偶率分别为63.33%±3.33%, 60.00%±11.55%和20.00%±10.00%, 1-2日龄雌蛾求偶率显著高于3日龄雌蛾的(P<0.05)(图3)。
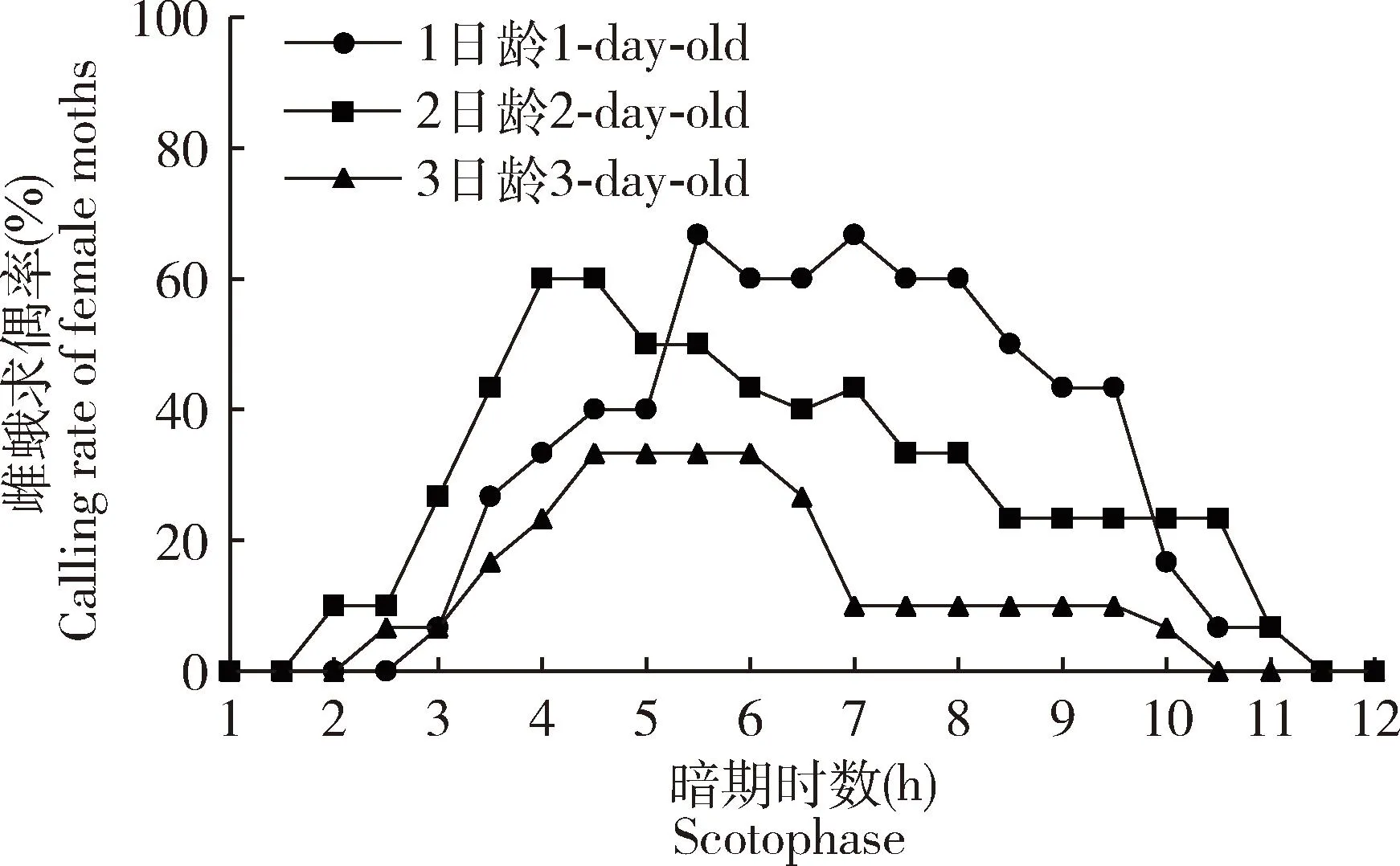
图2 葱兰夜蛾雌蛾求偶时辰节律Fig. 2 Time rhythm of the calling behavior of female moths of Brithys crini
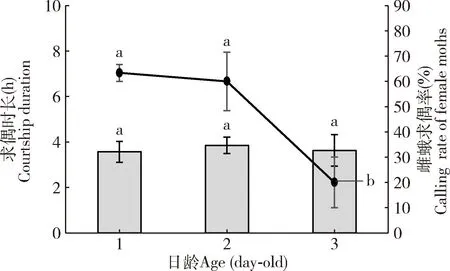
图3 不同日龄葱兰夜蛾雌蛾的求偶时长和求偶率Fig. 3 Courtship duration and calling rates of different day-old female moths of Brithys crini图中数据为平均值±标准误; 同一系列柱上/折线上不同字母表示差异显著(P<0.05, 最小显著性差异检验);图8同。Data in the figure are mean±SE. Different letters above bars /broken line in the same series indicate significant difference (P<0.05, least significant difference test). The same for Fig. 8.
2.2 葱兰夜蛾雌蛾性信息素腺体提取物的GC-EAD分析
根据上述葱兰夜蛾求偶行为节律,1-2日龄雌蛾求偶行为较活跃,求偶高峰期多在进入暗期后的4.0-9.0 h。选取正在求偶的1-2日龄处女雌蛾,从葱兰夜蛾雄蛾触角对处女雌蛾性信息素腺体提取物进行的GC-EAD分析可知,在保留时间为28.056 min处存在能引起葱兰夜蛾雄蛾触角电生理活性反应的组分(图4)。
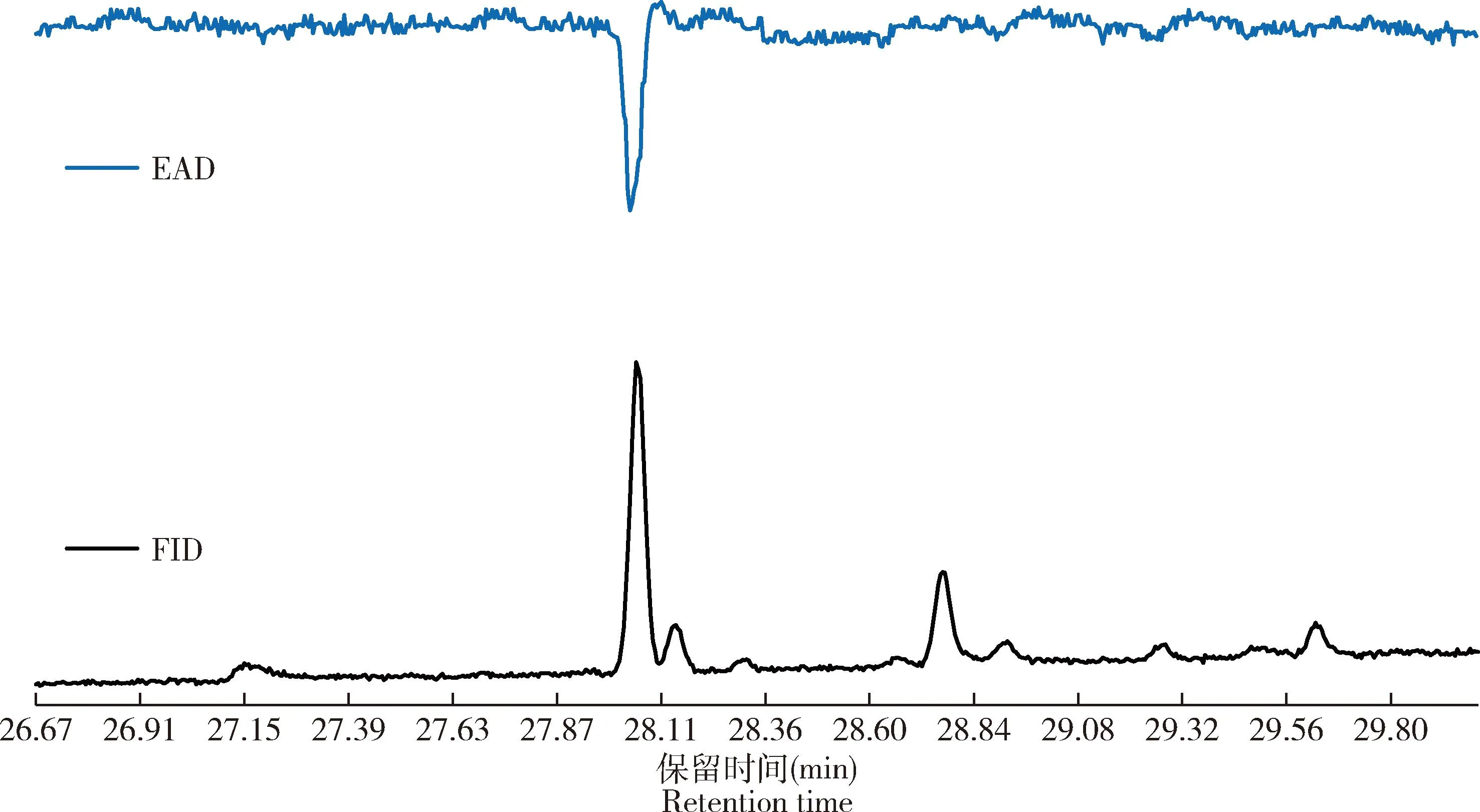
图4 葱兰夜蛾雄蛾触角对雌蛾性信息素腺体提取物的气相色谱-触角电位联用仪(GC-EAD)分析图谱Fig. 4 Gas chromatography-electroantennagram detection (GC-EAD) trace of the antennae of male moths of Brithys crini to the sex pheromone gland extracts from female mothsEAD: 触角电位反应信号Electroantennagram trace; FID: 气相色谱信号Gas chromatography trace.
2.3 葱兰夜蛾雌蛾性信息素腺体提取物的GC-MS分析
按照与GC-EAD测试相同的色谱条件,对葱兰夜蛾处女雌蛾性信息素腺体提取物进行GC-MS分析,得到相应的GC-MS图。结果显示,同样在保留时间28.056 min处存在疑似信息化学物质的色谱峰(图5)。使用Nist 14对该处的质谱图进行数据库检索,结果显示与该疑似组分匹配度最高的化合物为11-十六碳烯醛(11-16∶Ald)。
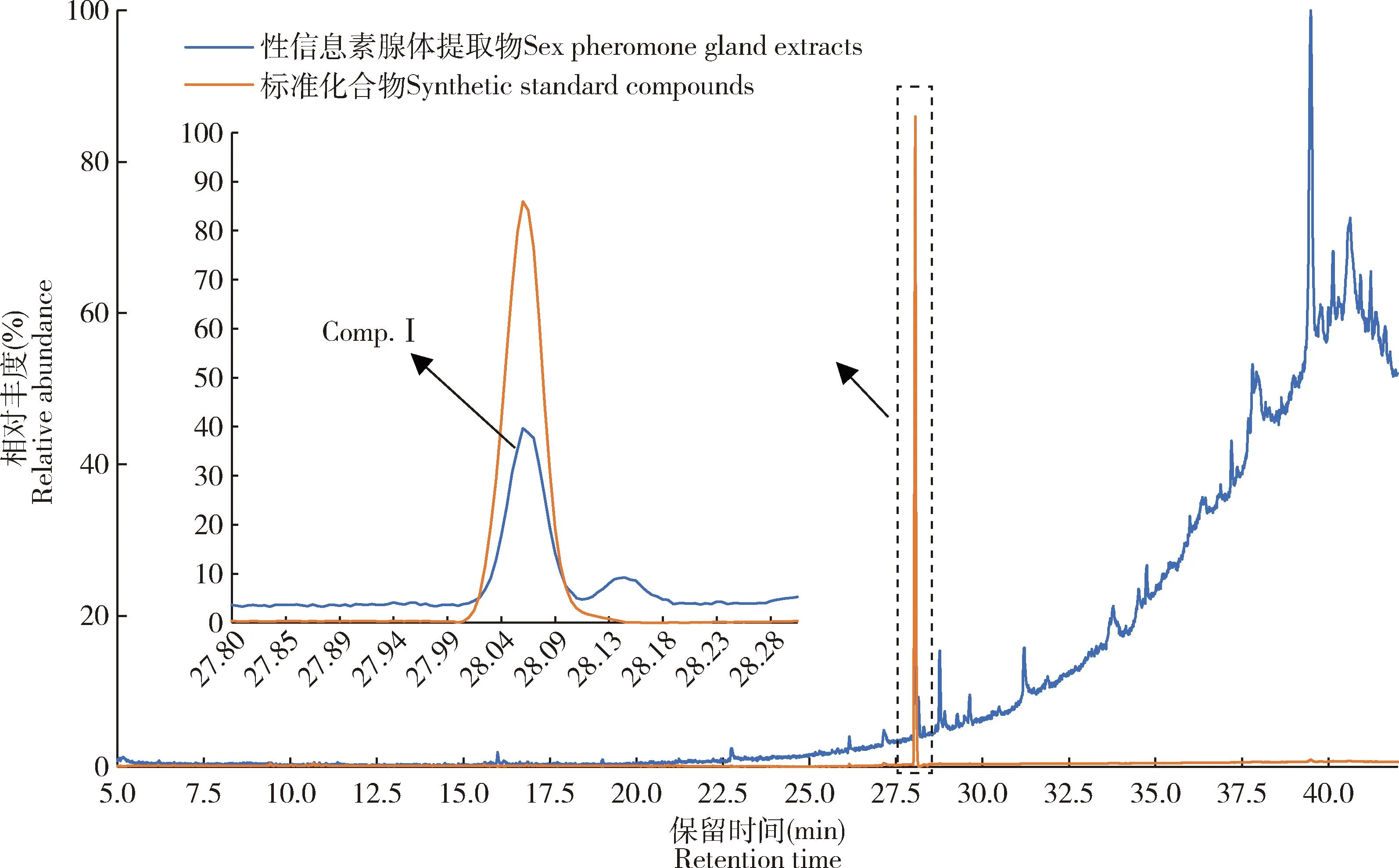
图5 葱兰夜蛾雌蛾性信息素腺体提取物和标准化合物的总离子流图Fig. 5 Total ion chromatogram of the sex pheromone gland extracts from the female moths of Brithys crini and synthetic standard compounds左边箭头示性信息素活性组分峰; 右边箭头示黑色虚线内总离子流图的局部放大。The arrow on the left indicates the peak of sex pheromone active component. The arrow on the right indicates the partial magnification of total ion chromatogram inside the black dotted line.
由于11-16∶Ald存在两种同分异构体Z11-16∶Ald和反-11-十六碳烯醛(E11-16∶Ald),因此同时对这两种化合物进行GC分析。结果显示雌蛾性信息素腺体提取物中的信息素疑似组分Comp. Ⅰ与Z11-16∶Ald和E11-16∶Ald具有相似的质谱图,其分子离子峰m/z238和特征碎片离子峰(m/z55, 69, 81)的相对强度与这两种标准化合物的特征碎片离子峰(m/z)基本保持一致,这进一步证实了Comp. Ⅰ为化合物11-16∶Ald(图6)。但是对比三者的保留时间和Kováts保留指数发现,信息素疑似组分Comp. Ⅰ与Z11-16∶Ald的保留时间和Kováts保留指数完全一致(表1),由此确定上述引起EAD响应的组分为Z11-16∶Ald。
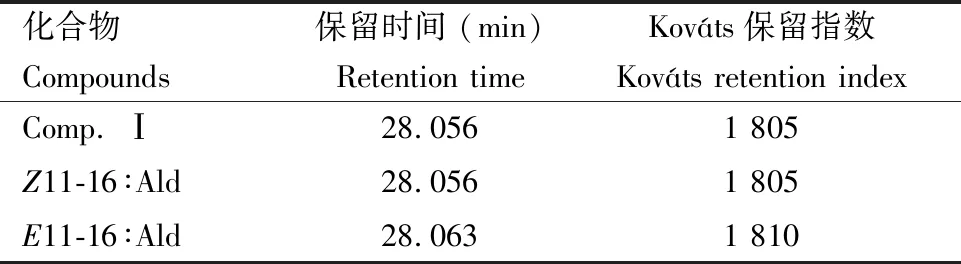
表1 葱兰夜蛾雌蛾性信息素组分Comp. Ⅰ和11-16∶Ald同分异构体在DB-5色谱柱上的保留时间和Kováts保留指数Table 1 Retention time and Kováts retention indices of the sex pheromone component Comp. Ⅰ of the female moths of Brithys crini and 11-16∶Ald isomers on DB-5 column

图6 葱兰夜蛾雌蛾性信息素腺体提取物和标准化合物的质谱图Fig. 6 Mass spectra of the sex pheromone gland extracts from the female moths of Brithys crini and synthetic standard compoundsA: 性信息素腺体提取物Sex pheromone gland extracts; B: Z11-16∶Ald; C: E11-16∶Ald.
2.4 葱兰夜蛾雌蛾性信息素活性组分含量
根据不同浓度标准化合物Z11-16∶Ald的剂量x与对应的峰面积相对丰度y建立剂量-峰面积标准曲线,分析结果表明剂量与峰面积之间的关系呈显著直线回归关系(df=3,P<0.05),标准曲线为y=1 000 006x-83 606,R2= 0.9886(图7)。根据上述标准曲线,计算出1和2日龄雌蛾性信息素腺体提取物中的Z11-16∶Ald平均浓度为(0.6211±0.0022) ng/μL,故而可以推算出单头处女雌蛾Z11-16∶Ald的平均含量为(3.11±0.01) ng。
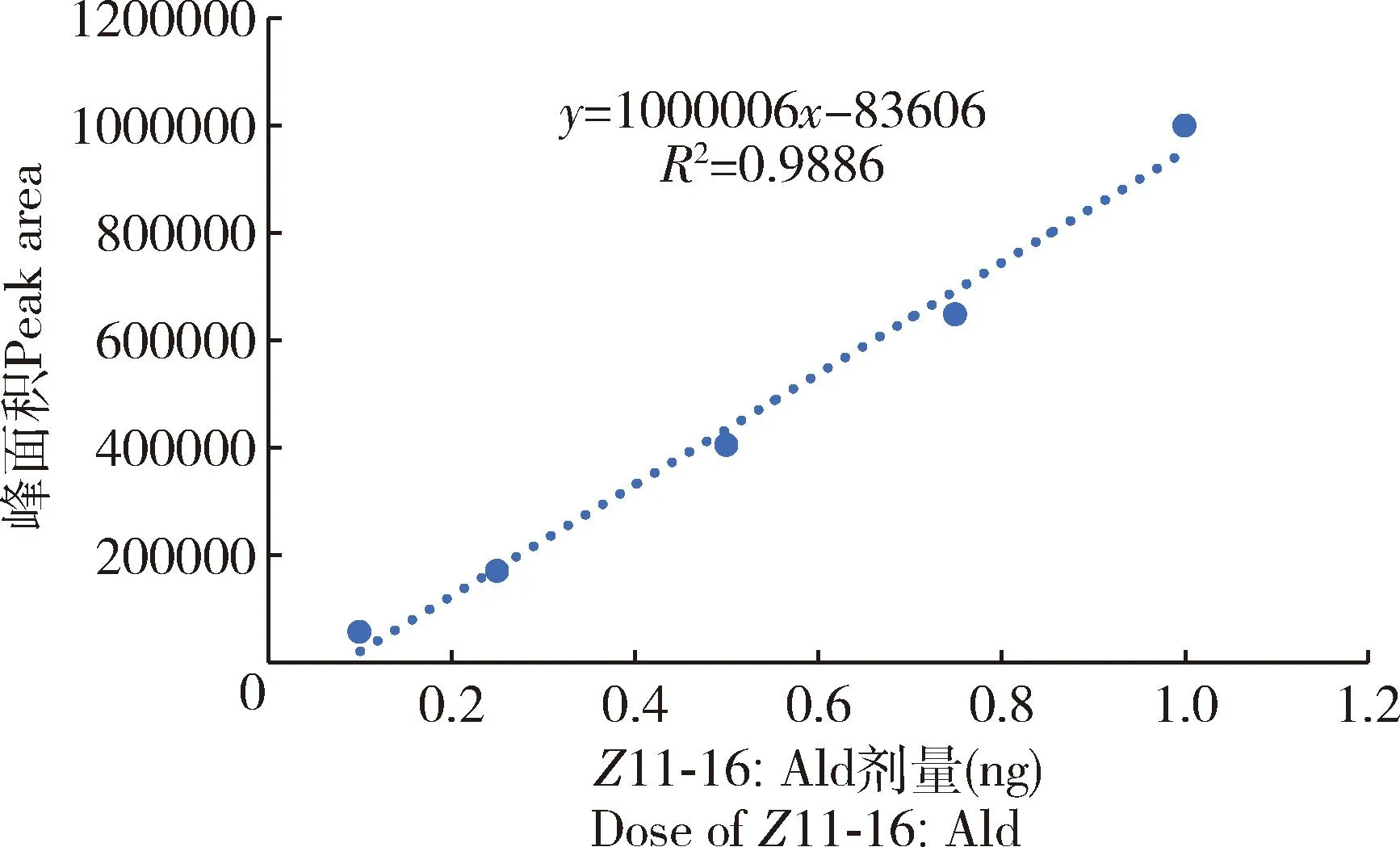
图7 Z11-16∶Ald的剂量-峰面积标准曲线Fig. 7 Standard curve of dose-peak area of Z11-16∶Ald
2.5 葱兰夜蛾雄蛾对标准化合物Z11-16∶Ald的EAG反应
分别取10 μL浓度为0.1, 1, 10和100 ng/μL的标准化合物Z11-16∶Ald溶液进行EAG测试。结果显示,雄蛾对Z11-16∶Ald的EAG反应存在明显的剂量效应:1 ng剂量引起的 EAG 反应与正己烷对照(CK)相近;加大剂量到10 ng后,引起的EAG反应要显著强于CK(P<0.05);随着刺激剂量增加至100 ng,引起的EAG反应最强;继续加大剂量到1 000 ng后,EAG反应较前者有所降低(图8)。
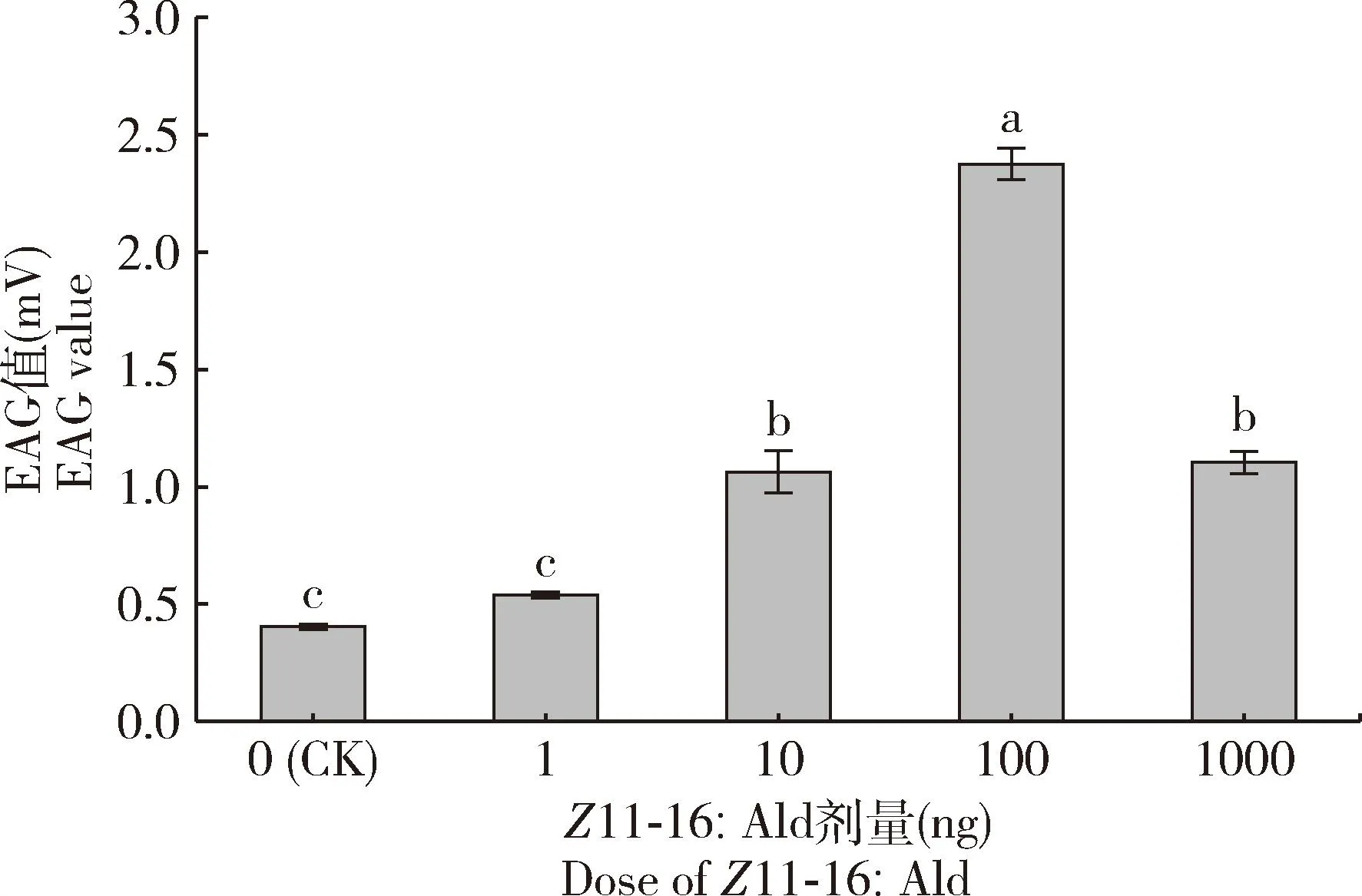
图8 葱兰夜蛾雄蛾对不同剂量Z11-16∶Ald的触角电位(EAG)反应Fig. 8 Electroatennogram (EAG) responses of the male moths of Brithys crini to different doses of Z11-16∶AldCK: 正己烷Hexane.
3 讨论
自然环境中光照强度随白天黑夜稳定而有规律的变化使得昆虫各种行为都具有昼夜节律(Groot, 2014; 钟春兰等, 2022)。蛾类昆虫的求偶节律受到光暗周期的显著影响:一般情况下,暗期越长,求偶持续时间越长 (Delisle and McNeil, 1986; Gerber and Howlader, 1987)。本研究设置的光周期为12L∶12D,旨在获得更长的求偶行为观察时间,也为了更好地开展雌蛾性信息素提取工作。此外,不同昆虫的性发育时间也有所不同,像棉红铃虫Pectinophoragossypiella需要1 d时间(许冬等, 2020),褛裳夜蛾Catocalaremissa雌蛾最早在羽化3 d以后才开始求偶,一周左右表现最为强烈(郑海霞等, 2014)。葱兰夜蛾羽化当天的成虫就已经性成熟,进入暗期后便开始求偶活动,这与木毒蛾Lymantriaxylina(左城等, 2020)、 巨疖蝙蛾Endoclitadavidi(李幸等, 2021)等蛾类相似。推测是因为葱兰夜蛾成虫寿命较短,所以性成熟的时间缩短,且在1日龄时就达到了求偶的最高峰。
此外,葱兰夜蛾随着日龄的增长,求偶开始时间会有所提前,这跟许多的蛾类是一致的,这可能是为了增加交配的机会,主要由于日龄较大的雌蛾释放信息素量的含量也会相应减少(Kamimura and Tatsuki, 1993)。通常,蛾类在羽化后的第1个晚上体内信息素含量较低,在第2和3个晚上急剧增加,然后随着雌蛾日龄的增大,性信息素的释放量会减少(Webster and Cardé, 1982),因此,蛾类昆虫性信息素的释放节律与其求偶节律是紧密相关的,故而可根据求偶行为推算出雌蛾释放信息素的时间段(Mazor and Dunkelblum, 2005; 郭前爽等, 2022)。本研究中,只观察到1-3日龄葱兰夜蛾存在求偶行为,1-2日龄雌蛾求偶行为较为活跃,求偶高峰期在进入暗期后的3.5-9.0 h。据此,选取正在求偶的1-2日龄处女雌蛾开展性信息素浸泡提取工作。
GC-MS技术能将性腺提取液的各种组分进行分离,同时获得各组分的质谱数据,借助化学物质信息库匹配出对应的化合物(Ando and Yamakawa, 2011)。但是绝大多数通过GC-MS技术鉴定的信息素化学结构都存在同分异构体,仅凭质谱信息很难确定信息素组分的顺反构象。将各异构体标准品配制成标准溶液进行EAG反应测试,是确定信息素化学结构的最直观和有效的方法。Kováts保留指数也是一项辅助判断化合物结构的重要指标,其数值只跟化合物本身的性质以及GC-MS所用色谱柱固定相有关(Marquesetal., 2000; 张雪梅等, 2008)。Yang等(2013)通过对天然信息素成分与合成标准品的保留指数比较,确定了桃展足蛾Stathmopodaauriferella的性信息素主要成分。朱俣伟等(2022)通过同样的方法确定了棉大卷叶螟Syleptaderogata性信息素主要组分的化学结构。本研究通过比较活性组分与11-16∶Ald的2种同分异构体在色谱柱上的保留时间和Kováts保留指数,从而确定了葱兰夜蛾性信息素活性组分为Z11-16∶Ald。
Kou等(1991)此前虽然通过GC-MS找出葱兰夜蛾的性信息素组分为Z11-16∶Ald,但是尚未开展性信息素组分的定量、顺反构象及电生理活性分析。因此,本研究通过比较活性组分与11-16∶Ald的2种同分异构体在色谱柱上的保留时间和保留指数,进一步确定了葱兰夜蛾性信息素活性组分的化学结构,并对单一雌蛾的性信息素进行定量。此外,本研究使用不同剂量的Z11-16∶Ald对葱兰夜蛾雄蛾触角进行EAG测试,发现10 ng以上剂量引起的EAG反应显著强于正己烷对照, 100 ng剂量引起的触角反应最强烈,剂量加大后会表现出抑制效果(图8);而实验测得葱兰夜蛾雌蛾性信息素组分的日滴度为3.11 ng。EAG测试结果与雌蛾该组分的日滴度测定结果契合,这为下一步开展野外试验提供了研究基础。
本研究中鉴定的Z11-16∶Ald是一种常见的醛类性信息素,是多种夜蛾科昆虫性信息素的主要组分(董改娟等, 2008; 宋亚茹等, 2017)。结合多数有关昆虫性信息素的报道发现,许多近缘种昆虫含有相同的性信息素主要组分,往往通过次要组分的差异表现出性信息素的特异性(Chenetal., 2018)。同一亚科不同属或同属不同种之间的昆虫性信息素活性化合物种类有可能相同,但是比例存在差异:实夜蛾亚科同属的Heliothispunctiger和Heliothisphloxiphaga的性信息素组分为Z11-16∶Ald和顺-11-十六碳烯醇(Z11-16∶OH),但二者比例不同,前者两种性信息素化合物配比为100∶75,后者为100∶7.2(Rothschildetal., 1982; Rainaetal., 1986);谷实夜蛾Heliothiszea和点实夜蛾Heliothispeligera性信息素的主要成分是Z11-16∶Ald,次要成分是顺-9-十四碳烯醛(Z9-14∶Ald), 但比例各有不同, 分别为100∶3.9和100∶14.6(Klunetal., 1980; Dunkelblum and Kehat, 1989)。同理可以推测,葱兰夜蛾性信息素除Z11-16∶Ald这一主要组分外,是否存在影响葱兰夜蛾化学通讯交流的次要组分,需要接下来进一步可以在性信息素提取上优化方法,选择更高效和高富集的技术,比如固相微萃取技术、顶空-固相微萃取和直接接触吸附萃取等;同时可使用具有更宽容的峰容量和色谱分辨率、更高灵敏度的全二维气相色谱飞行时间质谱联用仪(comprehensive two-dimensional gas chromatography/time-of-flight mass spectrometry, GC×GC/TOFMS),以适应葱兰夜蛾性信息素腺体提取物有效成分浓度较低的情形,或是探索次要组分在葱兰夜蛾化学通讯交流中的作用。
同时,虽然大部分昆虫性信息素通常是由两种或两种以上的化合物组成,但是,也有部分昆虫的信息素组分被鉴定为多元组分,实际应用过程中,单个组分即可具有较好的生物活性,像金纹细蛾Phyllonorycterringoniella的信息素组分为顺-10-十四碳烯乙酸酯(Z10-14∶Ac)和反4,顺10-十四碳烯乙酸酯(E4,Z10-14∶Ac),实际1 mgE4,Z10-14∶Ac就具有显著的诱蛾效果(Duetal., 2013);也有昆虫的信息素组分目前报道仅为1种活性组分,像苹果蠹蛾Cydiapomonella和杨小舟蛾Micromelalophasiversi(Liuetal., 2021),这与本研究中葱兰夜蛾的1种性信息素活性组分一致。
总之,葱兰夜蛾性信息素组分的鉴定,可有利于开发基于性信息素的高效引诱剂,不仅可以监测葱兰夜蛾的发生动态,还可以通过大量诱捕或者迷向干扰降低葱兰夜蛾的交配率,进而为葱兰夜蛾的绿色防控技术提供可靠的技术参考和理论依据。
猜你喜欢
新医学(2023年5期)2023-05-27
华人时刊(2020年19期)2021-01-14
今日农业(2020年17期)2020-12-15
今日农业(2020年13期)2020-12-15
广东医科大学学报(2020年6期)2020-02-06
中国医科大学学报(2020年4期)2020-01-08
今日农业(2019年13期)2019-08-12
能源(2018年5期)2018-06-15
文学少年(有声彩绘)(2017年1期)2017-02-14
安徽医药(2014年4期)2014-03-20
