
溴虫氟苯双酰胺对草地贪夜蛾的适合度及解毒酶活性的影响
2023-02-06 07:41:56姬雪娇王文杰杨庆杰翟艳慧芮昌辉
昆虫学报 2023年12期
姬雪娇, 崔 丽, 王文杰, 杨庆杰, 翟艳慧, 芮昌辉
(中国农业科学院植物保护研究所, 北京 100193)
草地贪夜蛾Spodopterafrugiperda又称秋粘虫,原产于美洲热带和亚热带地区,是一种世界性的迁飞性害虫。自2016年1月在非洲首次报道以来,目前已入侵超过130个国家 (Goergenetal., 2016; Rwomushana, 2022)。由于草地贪夜蛾适应力强(11~30 ℃均可发育、无滞育现象)、食性杂(偏好禾本科,在其他科也可见,如菊科、豆科)、繁殖力强(平均1 500粒/雌,最高可达2 000粒/雌)、迁飞远(每晚定向迁飞100 km)等特点,使其能迅速在迁入地定殖、扩散为害,因此其严重威胁我国玉米的生产安全(郭井菲等, 2019)。化学防治是控制草地贪夜蛾危害的主要手段。但由于长期的药剂选择和其自身的强大适应性,草地贪夜蛾已对40多种杀虫剂产生了不同程度的抗性,既包括有机磷、氨基甲酸酯类、拟除虫菊酯类等传统杀虫剂,也包括双酰胺类和多杀霉素等新型杀虫剂(王芹芹等, 2019; 吴益东等, 2019);另外室内生物测定和靶标突变分析均表明,入侵中国的草地贪夜蛾已对有机磷类、氨基甲酸酯类和拟除虫菊酯类杀虫剂产生了抗性,影响了其防治效果(Zhangetal., 2021; Wangetal., 2023)。
溴虫氟苯双酰胺(broflanilide)是由三井化学和巴斯夫共同开发的作用于γ-氨基丁酸受体(γ-aminobutyric acid receptors, GABARs)的新型间二酰胺类药剂,并进一步被IRAC归类为30组(γ-氨基丁酸门控氯离子通道变构调节剂)。与现有的杀虫剂无交互抗性。对鳞翅目、鞘翅目等昆虫有很高的活性(Katsutaetal., 2019)。于2020年12月28日在我国正式登记。作为一种新型的杀虫剂,溴虫氟苯双酰胺对草地贪夜蛾的毒力和田间药效均表现突出。10%溴虫氟苯双酰胺SC对草地贪夜蛾3龄和5龄幼虫均有较强的活性,72 h的校正死亡率均高于90%,田间药效试验表明,不同用量的10%溴虫氟苯双酰胺SC处理后1和7 d防效均可达96.33%以上,显著高于对照药剂5.7%甲维盐ME和200 g/L氯虫苯甲酰胺SC(王风良等, 2022)。王敏等(2022)研究也发现,溴虫氟苯双酰胺室内毒力显著高于硫虫酰胺和四唑虫酰胺,仅次于甲维盐;在田间用药后7 d,100 g/L溴虫氟苯双酰胺SC对玉米草地贪夜蛾的防效与对照药剂5.7%甲维盐DF和10%硫虫酰胺SC相当,均在93%以上,显著高于18%四唑虫酰胺SC的防效。溴虫氟苯双酰胺在草地贪夜蛾防治上表现出优异的速效性和持久性,是一种良好的田间应急防控替代药剂。目前,害虫对溴虫氟苯双酰胺的抗性研究还处于起步阶段,仅有1例关于小菜蛾Plutellaxylostella的报道。小菜蛾对溴虫氟苯双酰胺产生耐药性的风险较低,10代内敏感性几乎没有变化(Sunetal., 2022)。鉴于溴虫氟苯双酰胺对草地贪夜蛾的防治优越性和抗性情况还未见报道,本研究通过室内连续10代选育后,测定草地贪夜蛾对溴虫氟苯双酰胺的抗性、解毒酶活性以及适合度变化,旨在明确草地贪夜蛾对溴虫氟苯双酰胺的早期抗性风险和生化机理,以期为该杀虫剂在田间合理使用和草地贪夜蛾的综合防治提供理论依据。
1 材料与方法
1.1 供试昆虫
草地贪夜蛾为室内繁育种群(来自海南种群、华宁种群和玉溪种群的成虫交配所得的后代),一部分后代作为未汰选种群UNSEL(不接触药剂进行饲养),一部分后代作为汰选种群Bro-SEL(连续接触药剂进行汰选)。初孵幼虫通过人工饲料群体饲养至3龄幼虫后,再转移到在25 mL连体杯中进行单头饲养,直至化蛹,将蛹挑入9 cm培养皿中,待其开始羽化后转移至成虫笼中,进行交配。每日补充新鲜蜂蜜水并收集产卵纸。饲养条件为温度(25±2) ℃,相对湿度60%~70%和光周期14L∶10D。草地贪夜蛾人工饲料配方为: 75 g豆饼粉、75 g玉米面、 30 g啤酒酵母、210 g番茄酱、14 g琼脂和900 mL开水。
1.2 供试药剂与试剂
98.0%溴虫氟苯双酰胺原药,日本东京化成工业株式会社生产;酶液提取试剂[乙二胺四乙酸(ethylenediaminetetraacetic acid, EDTA)、二硫代苏糖醇(dithiothreitol, DTT)、苯基硫脲(phenylthiourea, PTU)、苯甲基磺酰氟(phenylmethanesulfonyl fluoride, PMSF)]均购自北京广达恒益;酶活性测定试剂[牛血清蛋白,考马斯亮蓝G-250,α/β-醋酸萘酯(α/β-napthyl acetate, α/β-NA),固蓝B盐,十二烷基硫酸钠(sodium dodecyl sulfate, SDS),毒扁豆碱,1-氯-2, 4-二硝基苯(1-chloro-2,4-dinitrobenzene, CDNB),还原性谷胱甘肽(glutathione, GSH)]均购自北京广达恒益;CYP450活性测定试剂盒购自上海恒远生物;增效醚(piperonyl butoxide, PBO)(95%),磷酸三苯酯(triphenyl phosphate, TPP)(95%)和马来酸二乙酯(diethyl maleate, DEM)(95%)购自上海阿拉丁试剂。
1.3 抗性选育
溴虫氟苯双酰胺汰选种群Bro-SEL建立:取1 mL浓度为0.25 mg/L溴虫氟苯双酰胺药液均匀涂抹在31.8 cm2人工饲料上(9 cm的培养皿),待药液自然风干后,将200~300头左右的初孵幼虫挑入其中。饲养4 d时统计死亡率,并将存活的幼虫转移至无毒饲料中饲养。每代汰选600~1 200头左右(3~4个重复)。选择压力在70%~80%死亡率之间。以0.25 mg/L的浓度连续汰选10代。取汰选后1代种群[Bro-SELC(G1)]和10代种群Bro-SEL 3龄幼虫用于毒力和解毒酶活性测定。
1.4 毒力测定方法
参考饲料表面涂毒法进行生物测定(Okumaetal., 2018)。配好的人工饲料和水按照1∶1(m/m)加热混匀,分装到孔直径16 mm、深17 mm的24孔盒内,每孔大约0.6 mL,冷却后加盖备用。使用0.05% (w/v) Triton X-100的水溶液将1%的母液(丙酮溶解)稀释5~7个浓度梯度。0.05% Triton X-100的水溶液作为空白对照。将配制好的溶液以每孔30 μL的体积添加到24孔盒饲料表面。室温下干燥后,用镊子往每孔加入1头3龄幼虫并用盖子封住。每个浓度处理24头幼虫。药剂处理后恢复条件同饲养条件。48 h时统计死亡率,计算致死中浓度(median lethal concentration, LC50)。以毛笔尖触碰幼虫后不能正常转曲为判断死亡的标准。
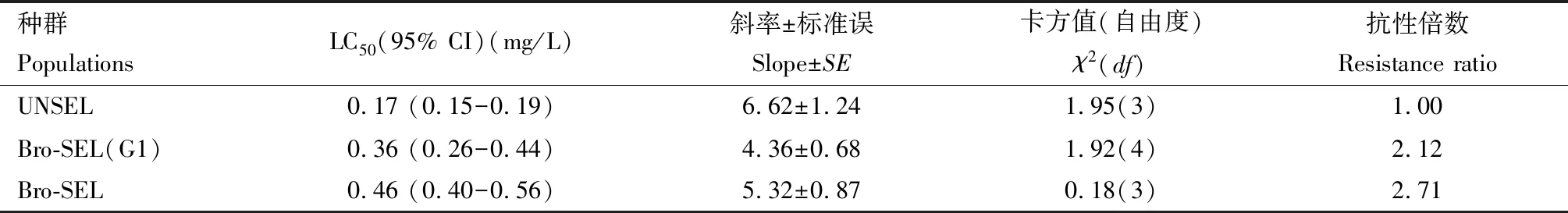
表1 溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫的毒力Table 1 Toxicity of broflanilide to the 3rd instar larvae of Spodoptera frugiperda
1.5 种群生命表建立
分别收集UNSEL和Bro-SEL种群成虫第2-3 天产的卵块,每个种群各收集10块。待孵化后,每块孵化的卵块中挑选10头幼虫。每头幼虫放于一个装有1 g左右饲料体积为25 mL塑料连体杯中,每个种群100头幼虫。每24 h观察并记录幼虫的发育和死亡情况,直至化蛹。饲料根据取食量和新鲜程度适时更换。化蛹第2天,用万分之一天平称重后再辨别雌雄,之后转移至一个3 cm培养皿中。待成虫出现后,将同一天羽化的雌雄成虫配对放入一个500 mL塑料量杯中(内有产卵纸和10%蜂蜜水), 并用棉纱布封口。每天更换产卵纸和棉纱布并适时补充蜂蜜水,记录每天的产卵量,直至成虫死亡。实验条件同前面的饲养条件。
1.6 解毒酶活性测定
1.6.1酶液制备:收集UNSEL和Bro-SEL种群的3龄中期幼虫,每5头加入1 mL 0.1 mol/L 磷酸缓冲盐液PBS[pH 7.8,包含1 mmol/L EDTA、1 mmol/L DTT、1 mmol/L PTU、1 mmol/L PMSF 和10%(w/w)甘油],冰上研磨匀浆,4 ℃下12 000 r/ min离心15 min,取上清液作为酶源,进行细胞色素P450单加氧酶(cytochrome P450 monooxygenase, CYP450)和谷胱甘肽S-转移酶(glutathione S-transferase, GST)活性测定。将该酶源用pH 7.0,0.04 mol/L PBS缓冲液稀释20倍用于测定羧酸酯酶(carboxylesterase, CarE)活性。共3次生物学重复。
1.6.2CYP450活性测定:采用双抗体夹心法测定CYP450活性。按照制造商说明书的要求,在有纯化的昆虫CYP450抗体包被的微孔板中加入样品、酶标试剂,形成抗体-抗原-酶标抗体复合物,经过温育和彻底洗涤后加底物3,3′,5,5′-四甲基联苯胺(TMB)避光显色,之后加终止液混匀,颜色由蓝变黄。用酶标仪测定450 nm下的吸光度。每个样品重复测定3次。
1.6.3CarE活性测定:参照van Asperen (1962)测定CarE活性,略有改动。酶反应体系包括600 μL反应混合液(含0.3 mmol/L α-NA, 1×10-4mol/L 毒扁豆碱和pH 7.0的0.1 mol/L PBS缓冲液)和120 μL的稀释酶液。然后在30 ℃金属浴中振动30 min。之后添加120 μL的显色剂(质量分数1%固蓝B盐∶5% SDS=5∶2, v/v, 现配现用),混匀后室温静置30 min,用酶标仪测定600 nm 下的吸光度。空白对照以pH 7.0,0.1 mol/L PBS缓冲液代替酶液。每个样品重复测定3次。将底物α-NA换为β-NA,酶活性测定方法不变。
1.6.4GST活性测定:参照Habig等(1974)方法以CDNB为底物测定GST活性。酶反应体系共1.5 mL,包含1.2 mL 66 mmol/L PBS缓冲液(pH 7.0,含2 mmol/L EDTA), 0.15 mL 50 mmol/L GSH, 0.05 mL 30 mmol/L CDNB和0.1 mL的酶液,空白对照以pH 7.0, 66 mmol/L PBS缓冲液代替酶液。立即在27 ℃, 340 nm下连续测定5 min内吸光度变化。每个样品重复测定3次。
1.6.5总蛋白含量测定:参照Bradford (1976)的考马斯亮蓝G-250法测定总蛋白含量。
1.7 增效剂对溴虫氟双酰胺增效作用的测定
用0.05% Triton X-100的水溶液将PBO, TPP和DEM分别配制成100 mg/L的溶液,然后用此溶液将1%溴虫氟苯双酰胺母液稀释成5~7个浓度梯度。以100 mg/L增效剂水溶液作为空白对照。其他条件同毒力测定。计算增效比(synergistic ratio)(单独使用溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫的LC50值/溴虫氟苯双酰胺加增效剂对草地贪夜蛾3龄幼虫的LC50值)。
1.8 数据分析
毒力回归方程和LC50采用统计软件DPS(7.05)生物测定模板进行计算;解毒酶活性利用统计DPS的LSD法进行显著性差异分析;利用 TWOSEX-MSChart 软件,基于年龄阶段、两性生命表理论对原始生命表数据进行分析(Chi and Liu, 1985; Chi, 1988),计算出每个种群不同发育阶段的历期、单雌产卵量、内禀增长率r、周限增长率λ、净增殖率R0、平均世代周期T;根据paired bootstrap test 法计算UNSEL和Bro-SEL 种群生命表参数和种群参数的平均值和标准误及其两者的显著性差异,进行100 000次评估。相对适合度R=Bro-SEL种群的R0/UNSEL种群的R0。绘图使用Origin 软件。生命表主要参数所用计算公式如下所述:
特定年龄存活率lx:
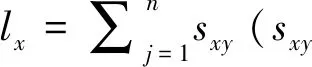
特定年龄繁殖力mx:

净增殖率R0:
内禀增长率r:
周限增长率λ:
λ=er;
平均世代周期T:
特定年龄-龄期生命期望值exj:

特定年龄-龄期繁殖力vxj:
2 结果
2.1 溴虫氟苯双酰胺对草地贪夜蛾UNSEL和Bro-SEL种群的毒力比较
经1和10代汰选后,溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫的毒力变化见表1。结果显示:草地贪夜蛾初孵幼虫以0.25 mg/L溴虫氟苯双酰胺汰选10代后,与未汰选种群UNSEL相比, 溴虫氟苯双酰胺处理48 h时对汰选10代种群Bro-SEL 3龄幼虫的LC50值由0.17 mg/L增加到0.46 mg/L;与未汰选种群UNSEL相比, Bro-SEL的抗性倍数为2.71,但汰选1代种群Bro-SEL (G1)的抗性倍数仅为2.12。这表明,经过持续的较高强度的选择压力,草地贪夜蛾对溴虫氟苯双酰胺的抗性倍数变化很少,抗性仅增加1.71倍,抗性水平仍处于敏感阶段。
2.2 草地贪夜蛾UNSEL和Bro-SEL种群生命表参数和适合度比较
2.2.1草地贪夜蛾UNSEL和Bro-SEL种群发育与繁殖比较:草地贪夜蛾经溴虫氟苯双酰胺10代汰选后,Bro-SEL种群的发育历期与繁殖力发生了显著的变化(表2)。与未汰选种群UNSEL相比, 汰选种群Bro-SEL的2龄幼虫历期, 3龄幼虫历期, 6龄幼虫历期, 成虫产卵前期(adult preoviposition period, APOP)和总产卵前期(total preovipositior period, TPOP)均极显著延长(P<0.01),蛹历期显著延长(P<0.05)。同时,Bro-SEL种群的单雌产卵量[(681.85±97.52)粒]也显著低于UNSEL种群的[(1 237.51±77.25)粒](P<0.01)。

表2 草地贪夜蛾UNSEL和Bro-SEL种群的发育历期和繁殖力Table 2 Developmental duration and fecundity of the UNSEL and Bro-SEL populations of Spodoptera frugiperda
2.2.2草地贪夜蛾UNSEL和Bro-SEL种群蛹重比较:草地贪夜蛾经溴虫氟苯双酰胺10代汰选后, Bro-SEL 种群平均蛹重较UNSEL种群没有明显改变(图1)。UNSEL和Bro-SEL种群的蛹的重量主要分布于200~250 mg范围,其中在230~240 mg范围内占比最高,分别为22.45%和21.52%。Bro-SEL种群的平均蛹重[(226.78±2.53) mg]略高于UNSEL种群的平均蛹重[(221.92±1.73) mg],但两者差异不显著(P>0.05)。但是,Bro-SEL种群部分蛹重可高达260~280 mg,总占比8.86%。说明长期接触溴虫氟苯双酰胺有可能会提高部分个体的蛹的重量。

图1 草地贪夜蛾UNSEL和Bro-SEL种群的蛹重分布图Fig. 1 Distribution of the pupal weight of the UNSEL andBro-SEL populations of Spodoptera frugiperda
2.2.3草地贪夜蛾UNSEL和Bro-SEL种群参数比较:草地贪夜蛾经溴虫氟苯双酰胺10代汰选后,Bro-SEL的种群参数较UNSEL种群也发生明显的变化(表3)。Bro-SEL种群的平均世代周期[(35.13±0.64) d]极显著高于UNSEL种群的[(30.61±0.28) d](P<0.01)。然而,Bro-SEL种群的内禀增长率[(0.15±0.01) d-1]、周限增长率[(1.16±0.01) d-1]和净增殖率(184.26±39.91)与UNSEL种群的[分别为(0.20±0.01) d-1, (1.23±0.01) d-1和(531.86±69.7)]相比均减小,且差异极显著(P<0.01)。Bro-SEL种群的相对适合度为0.35。说明Bro-SEL种群存在适合度代价。

表3 草地贪夜蛾UNSEL和Bro-SEL种群的种群参数Table 3 Population parameters of the UNSEL and Bro-SEL populations of Spodoptera frugiperda
2.2.4草地贪夜蛾UNSEL和Bro-SEL种群存活率与繁殖力的比较:与UNSEL相比,Bro-SEL种群的sxj曲线的重叠程度更高, 说明长期接触溴虫氟苯双酰胺会影响草地贪夜蛾个体间的发育速率。同时,Bro-SEL种群从2龄幼虫开始的各个阶段的存活率均低于UNSEL种群,并且Bro-SEL种群在6龄幼虫和蛹期的发育时间均高于UNSEL种群(图2: A, B)。

图2 草地贪夜蛾UNSEL (A)和Bro-SEL(B)种群的特定年龄-龄期存活率(sxj)Fig. 2 Age-stage-specific survival rates (sxj) of the UNSEL (A) and Bro-SEL (B) populations of Spodoptera frugiperdaEgg: 卵; L1-L6: 分别为1-6龄幼虫1st-6th instar larvae, respectively; Pupa: 蛹; Female: 雌成虫Female adult; Male: 雄成虫Male adult. 图4和5同。The same for Figs.4 and 5.
特定年龄存活率(lx)是指从卵存活到x年龄的可能性,忽略了龄期,描述了草地贪夜蛾随年龄变化的存活率,也是sxj的简化曲线。相比于UNSEL存活曲线在成虫前期坡度较平缓,Bro-SEL存活曲线在成虫前期坡度较陡(图3: A),说明Bro-SEL个体在幼虫阶段存活率低。在第37天时Bro-SEL成虫存活率开始高于UNSEL(图3: A)。特定年龄繁殖力mx表示草地贪夜蛾在x年龄的平均产卵量。Bro-SEL和UNSEL种群的mx曲线整体上表现先增后降的趋势,但Bro-SEL种群的mx曲线出现波动(图3: B),这表明药剂处理影响了草地贪夜蛾的产卵期,不仅导致平均产卵量降低,还导致出现了间断性产卵。种群特定年龄净繁殖率(lxmx)表示草地贪夜蛾存活个体在x年龄时的繁殖值,即净产卵量。Bro-SEL种群的净产卵量是明显低于UNSEL种群的(图3: C)。雌成虫特定年龄繁殖力(fx9)表示草地贪夜蛾雌成虫在x年龄的产卵量。fx9曲线和mx曲线具有相同的变化趋势(图3: D)。UNSEL雌成虫第26天开始产卵,第28天达到产卵高峰,雌成虫特定年龄繁殖力为216.53;Bro-SEL雌成虫第29天开始产卵,第35天达到产卵高峰,雌成虫特定年龄繁殖力为99.12(图3: D)。
在卵期,UNSEL和Bro-SEL种群的繁殖力贡献值均与各自的周限增长率相同,分别为1.23和1.16 d-1(表3)。雌成虫出现后,UNSEL种群在第27天的特定年龄-龄期繁殖值最大,为775.2(图4: A),而Bro-SEL种群在第31天达到最大特定年龄-龄期繁殖值,为395.4(图4: B)。

图4 草地贪夜蛾UNSEL (A)和Bro-SEL(B)种群的特定年龄-龄期繁殖值(vxj)Fig. 4 Age-stage-specific reproductive values (vxj) of the UNSEL (A) and Bro-SEL (B) populations of Spodoptera frugiperda
UNSEL和Bro-SEL种群的特定年龄-龄期生命期望值随年龄的增长而逐渐降低(图5: A, B)。在进入卵、1龄幼虫、5龄幼虫和蛹期,Bro-SEL种群的特定年龄-龄期生命期望值(分别为30.41, 27.41, 26.19和19.15 d)明显低于UNSEL种群的(分别为35.44, 32.44, 26.99和19.33 d);而在进入2龄幼虫、3龄幼虫、4龄幼虫、6龄幼虫、雌成虫和雄成虫期,Bro-SEL种群的特定年龄-龄期生命期望值(分别为31.89, 30.19, 29.19, 24.19, 15.30和10.38 d)则明显高于UNSEL种群的(31.70, 29.99, 28.99, 23.99, 12.34和9.15 d),尤其在雌成虫阶段,Bro-SEL种群的特定年龄-龄期生命期望值延长达2.96 d(图5: A, B)。

图5 草地贪夜蛾UNSEL (A)和Bro-SEL (B)种群的特定年龄-龄期生命期望值(exj)Fig. 5 Age-stage-specific life expectancy (exj) of the UNSEL(A) and Bro-SEL (B) populations of Spodoptera frugiperda
2.3 草地贪夜蛾UNSEL和Bro-SEL种群体内解毒酶活性比较
经溴虫氟苯双酰胺10代汰选后,草地贪夜蛾Bro-SEL种群3龄幼虫体内解毒酶活性发生了改变(图6)。与未汰选种群UNSEL(对照)相比,Bro-SEL种群体内CarE活性显著升高,其中以α-NA和β-NA为底物时的活性分别为对照的1.62和1.61倍,且与对照差异显著(P<0.05)(图6: C, D);而CYP450和GST活性与对照相比差异不显著(P>0.05)(图6: A, B)。

图6 草地贪夜蛾UNSEL和Bro-SEL种群3龄幼虫体内3种解毒酶活力比较Fig. 6 Comparison of the detoxification enzyme activities in the 3rd instar larvae of the UNSEL and Bro-SEL populations of Spodoptera frugiperdaA: 细胞色素P450单加氧酶(CYP450)活性Cytochrome P450 monooxygenase (CYP450) activity; B: 谷胱甘肽S-转移酶(GST)活性Glutathione S-transferase (GST) activity; C: 羧酸酯酶(CarE)活性, 以α-NA为底物Carboxylesterase (CarE) activity using α-NA as the substrate; D: CarE活性, 以β-NA为底物CarE activity using β-NA as the substrate. 图中数据为平均值±标准误(N=3); 柱上不同字母示差异显著(P<0.05, 最小显著差异法)。Data in the figure are mean±SE(N=3). Different letters above bars indicate significant difference(P<0.05, least significant difference test).
2.4 增效剂对溴虫氟苯双酰胺的增效作用
3种增效剂(PBO,TPP和DEM)对溴虫氟苯双酰胺有一定的增效作用(表4)。PBO和TPP与溴虫氟苯双酰胺混用后,能显著地增加对草地贪夜蛾3龄幼虫的毒力,增效比分别为1.31和1.42。而DEM同溴虫氟苯双酰胺混用后,也能增加对草地贪夜蛾 3 龄幼虫的毒力, 增效比为1.21; 但增效不显著。

表4 在草地贪夜蛾3龄幼虫的生测中增效剂对溴虫氟苯双酰胺的增效作用Table 4 Synergistic effects of synergists on boflanilide in bioassay against the 3rd instar larvae of Spodoptera frugiperda
3 讨论与结论
通过饲料表面涂毒法,对室内草地贪夜蛾初孵幼虫进行持续10代的定向强选择,草地贪夜蛾对溴虫氟苯双酰胺的抗性升高缓慢。汰选10代后,与未汰选种群UNSEL相比,汰选种群Bro-SEL抗性倍数仅增加1.71倍(表1)。表明草地贪夜蛾长期接触溴虫氟苯双酰胺,其敏感性变化较小,抗性水平仍处于敏感阶段。这与Sun等(2022)的研究结果是一致的。小菜蛾经过10代的持续选择,对溴虫氟苯双酰胺的抗性倍数仅为1.4倍。推测可能是由于溴虫氟苯双酰胺属于新上市的杀虫剂,在田间使用范围少,所收集到的种群本身含有抗性遗传基因少,从而影响其抗性发展。鉴于其对草地贪夜蛾优异的防控效果和抗性发展缓慢,因此,溴虫氟苯双酰胺可作为田间应急防控的替代药剂。但在田间长期使用该药剂防治草地贪夜蛾时,为延缓抗药性的发生和最大限度地发挥其防治效果,需要长期跟踪监测其抗药性发展状况,并注意与不同作用机理的药剂进行轮换使用。
昆虫抵抗杀虫剂的重要途经之一是体内的解毒酶将杀虫剂降解成无毒的物质排除体外。齐浩亮等(2017)发现,亚致死剂量的溴虫氟苯双酰胺能诱导小菜蛾体内的CarE活性升高,但对多功能氧化酶和GST活性没有影响;Zhan等(2021)研究发现溴虫氟苯双酰胺处理后的草地贪夜蛾体内的多功能氧化酶、CarE和GST均能做出积极的响应。本研究发现增效剂(PBO, TPP和DEM)与溴虫氟苯双酰胺一起使用能增加溴虫氟苯双酰胺对草地贪夜蛾的毒力,表明多功能氧化酶、CarE和GST均能参与草地贪夜蛾体内溴虫氟苯双酰胺的解毒代谢(表4)。同时,草地贪夜蛾经溴虫氟苯双酰胺10代汰选后,体内的CarE活性显著升高,其中以α-NA和β-NA为底物时的活性分别为对照的1.62和1.61倍(图6: C, D),但CYP450和GST活性无明显变化(图6: A, B)。说明,在草地贪夜蛾对溴虫氟苯双酰胺的抗性发展中,CarE可能发挥着更加积极的作用。
长期接触杀虫剂时,昆虫能通过一些行为、生理或遗传机制来降低杀虫剂的毒害,例如:通过组成型过表达解毒酶或在目标位点诱导突变,但同时可能也给它们的种群带来适合度代价,在发育和生殖方面产生负面影响((Kliot and Ghanim, 2012; Belinato and Martins, 2016)。因此,大多数杀虫剂抗性机制都与适合度代价有关,因为与敏感种群相比,这些变化往往对抗性昆虫的整体适合度产生不利影响。Sun等(2022)发现,小菜蛾经10代汰选后,幼虫期明显延长,繁殖力没有显著变化,相对适合度为0.93。本研究发现,草地贪夜蛾经溴虫氟苯双酰胺10代汰选后,其发育历期延长,幼虫存活率和繁殖力降低,且其种群参数(内禀增长率、周限增长率和净增殖率)也降低,相对适合度为0.35,表现出适合度代价(表2, 3)。但同时也观察到一些有利影响,如大蛹的比例相对提高和雌成虫寿命显著延长。这表明可能存在某种生理机制来补偿相关的适合度代价(图1)。另外,还发现雌成虫生命期望值显著延长2.96 d,这有利于增加汰选种群雌成虫的产卵天数来补偿产卵力下降(图5)。
综上所述,在连续10代汰选下,草地贪夜蛾对溴虫氟苯双酰胺的代谢抗性发展缓慢,敏感性几乎没有变化。长期的药剂汰选,草地贪夜蛾种群的发育与繁殖受到了一定程度的影响,表现出适合度代价。这些研究结果说明溴虫氟苯双酰胺在草地贪夜蛾的防治中将是一种重要的替代药剂。尽管如此,为确保溴虫氟苯双酰胺的优异防效,在田间使用时需要注意与不同作用机理的药剂轮换使用。此外,尽管多功能氧化酶、CarE和GST均参与草地贪夜蛾体内溴虫氟苯双酰胺的解毒代谢,但在抗性发展中,CarE可能起着更重要的作用。有关草地贪夜蛾对溴虫氟苯双酰胺的代谢抗性需要进一步研究。
猜你喜欢
空间科学学报(2021年1期)2021-05-22 01:36:34
今日农业(2020年17期)2020-12-15 12:34:28
今日农业(2020年13期)2020-12-15 09:08:51
世界农药(2019年3期)2019-09-10 07:04:16
世界农药(2019年3期)2019-09-10 07:04:14
今日农业(2019年13期)2019-08-12 07:59:08
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
中国果菜(2016年9期)2016-03-01 01:28:44
中国蔬菜(2015年9期)2015-12-21 13:04:40
火炸药学报(2014年3期)2014-03-20 13:17:38
