
共生菌Rickettsia响应宿主烟粉虱成虫日龄的免疫基因表达调控作用
2023-02-06 07:41:56赵冬晓张志春牛洪涛郭慧芳
昆虫学报 2023年12期
赵冬晓, 张志春, 牛洪涛, 郭慧芳
(江苏省农业科学院植物保护研究所, 南京 210014)
昆虫是地球上种类最多的生物类群,为自然生态系统的重要组成部分。极高的物种多样性得益于昆虫长期进化中形成的独特防御机制。当遭受病原物侵染时,昆虫可以通过先天免疫系统(innate immune system)的细胞免疫或体液免疫来吞噬病原物或激活胞内信号通路产生溶菌酶、抗菌肽、凝集素等物质抵抗病原物(刘小民和袁明龙, 2018)。此外,昆虫体内普遍存在丰富的共生菌,它们对宿主具有重要作用,包括提高宿主适合度、提升对病原菌的防御能力等(Himleretal., 2011; King, 2020)。研究表明,共生菌能通过调控宿主免疫、分泌毒素、竞争宿主资源等方式抑制病原菌侵染,被誉为昆虫的“外源免疫”(King, 2020)。因此,天然免疫系统提供的内源防御(intrinsic defense)与共生菌提供的外源防御(extrinsic defense)形成了昆虫抵抗病原物的双重防线(Hooperetal., 2012)。
研究发现,共生菌对宿主昆虫的天然免疫系统具有一定的调控作用,且调控方式在不同昆虫-共生菌互作体系中差异较大。有些共生菌通过激活昆虫天然免疫来提高宿主对外源微生物的抵抗能力。例如,Wolbachia会导致宿主埃及伊蚊Aedesaegypti体内NADPH氧化酶和活性氧(reactive oxygen species, ROS)簇的高表达而激活免疫应答中的Toll信号通路,产生抗菌肽(antimicrobial peptide)和防御素(defensis),抑制登革热病毒(dengue virus, DENV)对埃及伊蚊的侵染(Xietal., 2008; Kambrisetal., 2009; Moreiraetal., 2009)。然而,有些具有保护功能的共生菌表现为削弱昆虫自身的免疫功能。因为宿主将抵抗病原物的任务“外包”给这类保护性共生菌来承担,进而节省了宿主在免疫上的能量投入(King and Bonsall, 2017)。例如,携带具有抗寄生蜂、病原菌等功能的共生菌Hamiltonelladefense和Serratiasymbiotica能分担豌豆蚜Acyrthosiphonpisum先天免疫功能,因为蚜虫基因组缺少部分免疫相关基因和肽聚糖识别蛋白(peptidoglycan recognition protein, PGRP)基因等一些天然免疫系统中的关键因子(Gerardoetal., 2010)。另外,从宿主的整个生命过程看,免疫功能的状态与机体衰老呈现平行关系(Yousefzadehetal., 2021),但共生菌对宿主免疫的调控作用是否会随宿主生命周期而发生波动却鲜有报道。
溶菌酶与防御素是昆虫先天免疫过程中的广谱的抗菌效应分子(刘小民和袁明龙, 2018)。昆虫在受到病原微生物侵染时,溶菌酶通过水解细菌细胞壁内的肽聚糖来杀死入侵的病原体,防御素在血淋巴及消化道中产生抗菌肽,在昆虫的天然免疫中扮演重要角色(Beck and Habicht, 1996)。已有研究鉴定出烟粉虱Bemisiatabaci的溶菌酶基因和防御素相关基因,它们在烟粉虱抵抗细菌、真菌侵染中起重要作用(Wangetal., 2013; 于洁等, 2016)。
烟粉虱是世界性害虫,传播危害性极大的番茄黄化曲叶病毒(tomato yellow leaf curl virus, TYLCV)等植物病毒(Wangetal., 2017; 褚栋和张友军, 2018)。烟粉虱体内携带多种共生菌,它们能为宿主补充植物汁液中缺少的营养,也能提高宿主适合度及抗逆能力等,对烟粉虱种群扩张与传播植物病毒等起到了促进作用(Himleretal., 2011; Guoetal., 2014; Luanetal., 2018; Zhaoetal., 2018; Wangetal., 2020)。连续多年的调查结果表明,共生菌Rickettsia在我国烟粉虱自然种群中快速扩散,且其感染量在共生菌群落中占绝对优势(Zhaoetal., 2021, 2022, 2023)。研究发现,Rickettsia对宿主烟粉虱具有保护功能,能够延缓杀虫真菌、环境致病菌等对烟粉虱的致死作用(Hendryetal., 2014; Zhaoetal., 2023),并有效防御丽蚜小蜂Encarsiaformosa的寄生(刘媛等, 2019)。然而,共生菌Rickettsia会对烟粉虱先天免疫产生怎样的影响尚未见报道。基于Rickettsia对提高烟粉虱适合度、抵御病原物等生态功能,以及溶菌酶、防御素等相关基因在害虫免疫系统中的关键地位,本研究通过比较感染和不感染Rickettsia烟粉虱不同日龄成虫体内溶菌酶与防御素相关免疫基因表达量,跟踪了烟粉虱不同日龄成虫体内Rickettsia相对含量与免疫基因表达量的动态,并分析了二者的相关性。研究结果将明确共生菌Rickettsia对宿主烟粉虱免疫基因的调控作用,并探明烟粉虱成虫日龄对Rickettsia调控宿主免疫基因的影响,有助于深入认识基于共生菌与宿主先天免疫协同的烟粉虱免疫防御机制,并为进一步解析共生菌与宿主协同进化的生理与生态机理提供了依据。
1 材料与方法
1.1 烟粉虱品系的建立
烟粉虱于2019年3月采集于南京田间茄子,采回后在人工气候室内棉花苗上饲养[温度(27±1) ℃,相对湿度60%±5%,光周期14L∶10D]。将新羽化烟粉虱雌雄成虫放在自制的微型笼里(直径6 cm,高8 cm),并夹在棉花叶片上进行单对饲养,产卵后7 d,PCR检测母代共生菌感染类型,待子代羽化为成虫,抽5~10头检测共生菌,检查是否与母代相符(Zhaoetal., 2018)。以上步骤重复进行,并把相同感染类型放在同一养虫笼饲养,建立100%感染共生菌Rickettsia的烟粉虱品系(R+)、 100%不感染Rickettsia的烟粉虱品系(R-)和45.83%感染Rickettsia的烟粉虱混合品系(mixture)。
1.2 样品收集与cDNA合成
收集R+品系、R-品系以及mixture混合品系烟粉虱成虫(对其发育期不作控制Rickettsia感染率为收集样品实测感染率),每个样品50~100头烟粉虱成虫,每个品系设置3~5个生物学重复。另外,收集初羽化24 h内、3-7日龄以及大于15日龄R+烟粉虱成虫,每个样品50~100头烟粉虱成虫,每个日龄设置5个生物学重复。所有样品用动物组织RNA提取试剂盒(Promega,美国)提取总RNA,用反转录试剂盒(Promega,美国)合成cDNA后置于-20 ℃保存。
1.3 qRT-PCR检测
qRT-PCR检测以烟粉虱Btβ-actin为内参基因,以1.2节合成的cDNA为模板,扩增R+品系、R-品系以及感染率为45.83%的二者混合品系mixture烟粉虱成虫中免疫关键因子溶菌酶基因BtLyz1和防御素基因Btdef。鉴于昆虫日龄增长带来的机体衰老是影响机体免疫水平的重要因素,以1.2节不同日龄的R+品系烟粉虱成虫cDNA为模板扩增细胞凋亡基因BtCaspase,以精确反映出不同日龄烟粉虱成虫的衰老程度(Bulletal., 2012)。在此基础上,对1.2节不同日龄的R+品系烟粉虱成虫cDNA进行共生菌Rickttsia16S rRNA基因、BtLyz1和Btdef的扩增。反应体系(20 μL): DNA模板2 μL, SYBR Premix 10 μL, 上下游引物(10 μmol/L)各0.4 μL, ddH2O 7.2 μL。反应使用SYBR®Premix Ex TagTM(Yeason,上海),在Roche LightCycler®480II实时荧光定量PCR仪进行扩增。反应程序: 95 ℃预变性30 s; 95 ℃变性5 s, 60 ℃退火 31 s, 40 个循环; 95 ℃ 15 s, 60 ℃ 1 min, 95 ℃ 15 s。定量PCR的引物见表1。每个样品5个生物学重复,3个技术重复,采用2-ΔΔCt法计算各样本目的基因的相对表达量。

表1 引物信息Table 1 Primer information
1.4 数据分析
感染和不感染共生菌Rickettsia烟粉虱成虫中BtLyz1和Btdef表达量比较采用单因素方差分析。感染共生菌Rickettsia烟粉虱不同日龄成虫中BtLyz1,Btdef,BtCaspase和Rickettsia16S rRNA基因表达量比较采用单因素方差分析。为进一步揭示Rickettsia对烟粉虱免疫基因的调控作用及规律, 利用Pearson法对R+烟粉虱不同日龄成虫Rickettsia16S rRNA基因表达量与BtLyz1和Btdef表达量进行相关性分析。
2 结果
2.1 Rickettsia提高烟粉虱免疫基因表达量
qRT-PCR检测结果显示,感染Rickettsia的烟粉虱(R+) 成虫中BtLyz1相对表达量显著高于不感染Rickettsia的烟粉虱(R-)以及混合品系(mixture)烟粉虱成虫中的(F2,6=390.288,P<0.001)(图1: A)。Btdef在R+品系成虫的相对表达量最高,其次是mixture品系成虫中的,在R-品系成虫中的相对表达量最低(F2,6=7.039,P=0.027)(图1: B)。

图1 烟粉虱成虫中溶菌酶基因BtLyz1(A)和防御素基因Btdef(B)的相对表达量Fig. 1 Relative expression levels of lysozyme gene BtLyz1 (A) and defensin gene Btdef (B) in adult Bemisia tabaciR+: 100%感染Rickettsia的品系100% Rickettsia-infected line; R-: 100%不感染Rickettsia的品系 100% Rickettsia-free line; Mixture: 感染率为45.83%的R+/R-混合品系R+/R- mixed line with the infection rate of 45.83%. 图中数据为平均值±标准差;柱上不同小写字母表示经单因素方差分析在P<0.05水平差异显著。Data in the figure are mean±SD. Different small letters above bars indicate significant difference at P<0.05 level by one-way ANOVA. 图2同。The same for Fig. 2.
2.2 不同日龄烟粉虱成虫免疫基因表达量和Rickettsia相对含量
结果显示,烟粉虱初羽化24 h内、3-7日龄以及大于15日龄成虫中BtCaspase的相对表达量具有显著差异,其中从初羽化24 h内到3-7日龄再到大于15日龄,烟粉虱成虫的BtCaspase相对表达量呈现显著上升趋势(F2,12=52.156,P<0.001)(图2)。该结果在分子水平上证明了烟粉虱的生理状态随日龄增加而逐渐衰退,且本研究对烟粉虱成虫生命周期阶段的划分具有较高的合理性。

图2 感染Rickettsia烟粉虱不同日龄成虫中细胞凋亡基因BtCaspase的相对表达量Fig. 2 Relative expression levels of apoptotic gene BtCaspasein different day-old adults of Bemisia tabaciinfected with Rickettsia0: 初羽化24 h内Newly emerged within 24 h.
在此基础上,分别测定了初羽化24 h内、3-7日龄以及大于15日龄烟粉虱成虫体内Rickettsia16S rRNA基因及BtLyz1和Btdef的相对表达量。结果显示, 3个时期的烟粉虱成虫体内Rickettsia16S rRNA基因、BtLyz1和Btdef相对表达量差异显著,且波动规律相似(图3)。其中3-7日龄烟粉虱成虫的Rickettsia16S rRNA基因相对表达量显著高于初羽化24 h内以及大于15日龄的(F2,12=18.758,P<0.001)(图3)。与之类似,3-7日龄烟粉虱成虫的BtLyz1与Btdef相对表达量显著高于初羽化24 h内以及大于15日龄成虫的(BtLyz1:F2,12=8.259,P=0.006;Btdef:F2,12=5.414,P=0.021)(图3)。
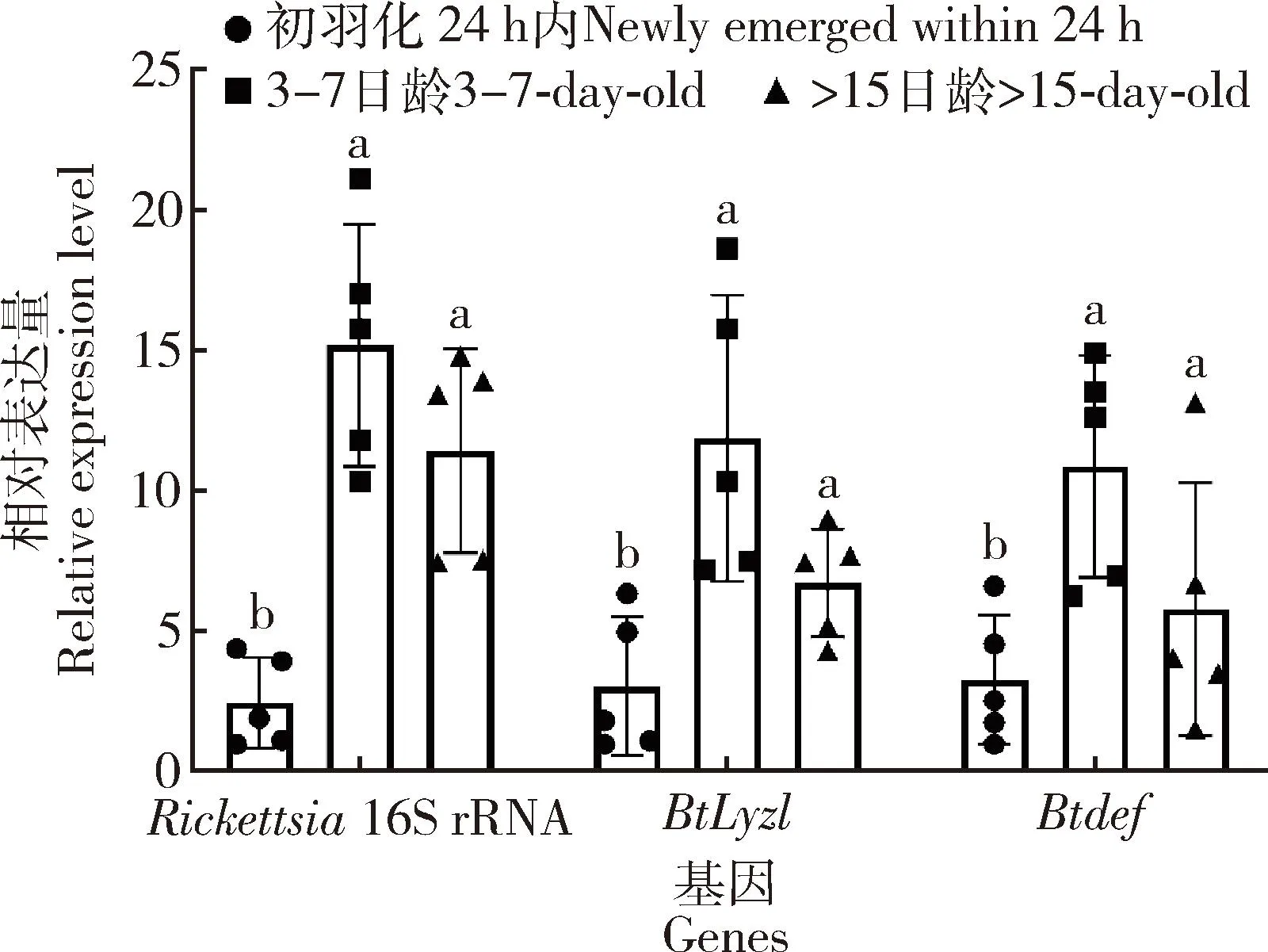
图3 感染Rickettsia不同日龄烟粉虱成虫中Rickettsia16S rRNA基因、BtLyz1和Btdef的相对表达量Fig. 3 Relative expression levels of Rickettsia 16S rRNAgene, BtLyz1 and Btdef in different day-old adults of Bemisia tabaci infected with Rickettsia图中数据为平均值±标准差;柱上不同字母表示经单因素方差分析在P<0.001 水平差异显著。Data in the figure are mean±SD. Different cetters above bars indicate significant difference at P<0.001 level by one-way ANOVA.
2.3 不同日龄烟粉虱成虫体内Rickettsia 16S rRNA基因表达量和免疫基因表达量的相关性
结果显示,在初羽化24 h内和3-7日龄烟粉虱成虫中Rickettsia16S rRNA基因相对表达量与BtLyz1相对表达量呈现显著正相关(初羽化24 h内:P=0.001; 3-7日龄:P=0.0113)(图4: A, B),且与Btdef相对表达量同样呈显著正相关(初羽化24 h内:P=0.0379; 3-7日龄:P=0.0229)(图4: D, E)。但是在处于衰老期(>15日龄)的烟粉虱成虫中,Rickettsia16S rRNA基因相对表达量与BtLyz1和Btdef相对表达量均不具有相关性(BtLyz1:P=0.9097;Btdef:P=0.9972)(图4: C, F)。
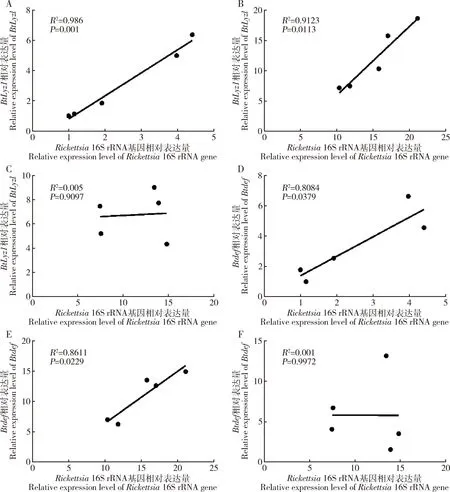
图4 感染Rickettsia烟粉虱不同日龄成虫中Rickettsia 16S rRNA 基因相对表达量与BtLyz1(A-C)和Btdef(D-F)相对表达量的相关性Fig. 4 Correlations between the relative expression level of Rickettsia 16S rRNA gene and relative expression levels of BtLyz1 (A-C) and Btdef (D-F) in different day-old adults of Bemisia tabaci infected with RickettsiaA, D: 初羽化24 h内Newly emerged within 24 h; B, E: 3-7日龄3-7-day-old; C, F: >15日龄>15-day-old.
3 讨论
与宿主共生的细菌群落是影响宿主健康的重要因素。共生菌调节宿主免疫反应是影响宿主健康的途径之一(Hooperetal., 2012)。本研究中,共生菌Rickettsia能够提高宿主烟粉虱免疫关键因子溶菌酶、防御素基因的表达量(图1)。在细胞凋亡基因低表达的初羽化24 h内和3-7日龄烟粉虱成虫中,Rickettsia菌量(Rickettsia16S rRNA基因表达量)的上升引起免疫基因表达量上升,而在细胞凋亡基因高表达的衰老(大于15日龄)烟粉虱成虫体内Rickettsia菌量与宿主免疫基因表达不再相关(图4)。这表明,宿主衰老等关键生理过程是维持共生菌与宿主免疫互作关系的重要基础。
进化和生态背景下,生物体免疫功能的变化被研究者定义为“生态免疫(ecological immunology)”(Rolff and Siva-Jothy, 2003)。随着“生态免疫”概念的提出,昆虫与其体内共生菌等微生物协同进化背景下的天然免疫功能变化越来越受到关注(Kwongetal., 2017)。研究表明,共生菌通过调控宿主免疫水平而保护宿主抵抗病原物。另外,共生菌对宿主免疫的调控有利于维持宿主共生菌的高密度,进而增强共生菌对宿主的生态功能(Nicholsetal., 2021; Goldsteinetal., 2023)。最新研究发现,共生菌Hamiltonella和Regiella抑制宿主豌豆蚜A.pisum酚氧化酶和血细胞色素等免疫基因表达是它们能在宿主体内高密度存在的重要原因(Kaechetal., 2021; Nicholsetal., 2021)。本研究发现,共生菌Rickettsia引起宿主烟粉虱体内溶菌酶与防御素相关免疫基因表达量显著提高。由此推测,Rickettsia的存在对其他微生物的入侵与增殖具有抑制作用,这为解析Rickettsia在烟粉虱共生菌群落中的主导地位以及抑制病原真菌在烟粉虱体内的增殖速度等生态现象提供了一定的分子依据(Zhaoetal., 2021, 2022, 2023)。
衰老是生命发生、发展过程中机体从组织结构到生理功能的丧失和退化的过程(Nishiuraetal., 2022)。在整个生命过程中,机体衰老和免疫功能的状态呈现平行关系,即在机体衰老的过程中,免疫细胞受到内源性DNA损伤导致免疫功能下降即免疫衰老(Yousefzadehetal., 2021)。本研究中,以细胞凋亡基因表达量上升为标志的烟粉虱机体衰老(图2),引起了其免疫基因表达量的显著下降以及共生菌Rickettsia量的显著下降(图3)。在衰老期,低量的Rickettsia与低表达量的溶菌酶、防御素基因之间不再有相关性,表明这个阶段中Rickettsia对宿主免疫不再具有调控功能。前期研究表明,伴随宿主衰老的细胞凋亡水平上升是导致宿主体内共生菌损失的重要生理级联反应(Roberty and Plumier, 2022);而共生菌对宿主产生生态功能往往是建立在一定量基础上,共生菌量低于阈值是对宿主功能强度的直接影响因素(Nicholsetal., 2023)。由此推测,宿主机体衰老和免疫功能下降以及共生菌无法从宿主获取大量增殖所需能量的双重作用,导致了共生菌对衰老宿主免疫调控作用的变化与扰动。
本研究结果表明,宿主体内共生菌量以及宿主生理状态对确定共生体或宿主免疫系统是否为宿主提供最佳保护策略至关重要。由此推测,共生菌Rickettsia与烟粉虱免疫基因表达的动态互作,为衰老水平较低的中、低日龄烟粉虱成虫提供防御病原物等保护功能,而对高日龄的衰老烟粉虱成虫保护功能弱,但这一推测有待后续研究进一步证实。未来对共生菌与宿主昆虫互作的研究要考虑宿主衰老等影响共生菌量的关键生理因素,多角度、多维度探索共生菌与宿主互作的动态规律,以更科学、更全面地理解共生菌与宿主的协同进化机制。
猜你喜欢
昆虫学报(2022年11期)2022-12-28 02:44:18
成都大学学报(自然科学版)(2021年1期)2021-05-22 01:31:18
植物保护(2018年5期)2018-12-05 10:36:16
湖北植保(2017年4期)2017-08-31 11:09:49
小溪流(画刊)(2017年8期)2017-08-09 19:49:55
小溪流(画刊)(2017年6期)2017-07-10 18:11:40
阅读与作文(小学低年级版)(2015年6期)2015-05-30 10:57:28
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:21:01
中国兽药杂志(2012年4期)2012-11-06 07:26:12
郑州大学学报(理学版)(2012年4期)2012-03-25 13:59:08
