
辣椒锌指蛋白DnaJ-Like基因家族鉴定及对高温胁迫的表达响应
2023-02-02 06:46:46段敏杰李怡斐杨小苗王春萍黄启中黄任中张世才
生物技术通报 2023年1期
段敏杰 李怡斐 杨小苗 王春萍 黄启中 黄任中 张世才
(1.重庆市农业科学院蔬菜花卉研究所,重庆 401329;2.重庆市农业科学院生物技术研究所,重庆 401329)
DnaJ 蛋白是一类分子量为41 kD 的热激蛋白(Hsp40),最早在大肠杆菌(Escherichia coli)中发现,又称为J-蛋白[1]。J-蛋白通常包含4 个功能结构域:N 末端J-结构域、G/F 结构域、锌指结构域(CxxCxGxG)及羧基末端区[2-3]。其中J-结构域具有极其保守的组氨酸/脯氨酸/天冬氨酸(His/Pro/Asp,HPD)三肽,是J-蛋白最核心特征[4]。J-蛋白通常作为伴侣蛋白,与热激蛋白HSP70 结合,参与蛋白质折叠、展开、组装和降解,并维持蛋白质稳定[5]。然而越来越多的研究发现DanJ 蛋白的一些活性并不依赖于完整的J-结构域,如当HPD 三肽发生突变甚至缺失的情况下,DanJ 蛋白仍能保持其活性[6-7],这类蛋白被划分至J-Like 蛋白。依据拟南芥(Arabidopsis thaliana)的分类方式[8],该类蛋白可分为三类:具有类似J-结构域的DNAJD 蛋白,具有类似锌指结构域和C 末端结构域的DNAJF 蛋白,具有位于羧基端重复DnaJ CxxCxGxG 或GRXLike(GRL)CxxCx7CxxC 锌指结构域的DnaJ-like 锌指蛋白(DNAJE)[8-12],DNAJE 蛋白是目前研究最多、最为复杂的一类J-Like 蛋白。
已有研究表明,DNAJE 蛋白在植物光合复合体组装中发挥重要作用,而光合复合体组装对环境干旱、温度、光照等的改变非常敏感,每一种变化都会引起植物氧化还原反应的失衡,进而影响植物抗逆性[13]。最早在玉米(Zea maysL.)中鉴定到一个DNAJE基因BSD2[14],之后在莱茵衣藻(Chlamydomonas reinhardtii)中鉴定到同源BSD2基因[15],该基因能够通过转录后调控rbcl 蛋白,从而促进叶绿体功能的正常发挥。同时,在拟南芥[16-20]、百脉根(Lotus corniculatusL.)[20]等中也鉴定到CYO1、SCO2、ANGULATA7等DNAJE基因,其均与叶绿体发育相关,在维持氧化还原反应和光合作用平衡中发挥作用。Fristedt 等[21]和Lu等[22]研究也发现,DNAJE 蛋白在光系统Ⅱ的维护和光系统Ⅰ积累中发挥特殊作用。目前,所有分离鉴定已知功能的DNAJE 蛋白中,仅THRUMIN1 属于GRL CxxCx7CxxC 型,其他均为DnaJ CxxCxGxG 型锌指蛋白[23]。
植物生长发育过程中会遭受诸多不利环境因子胁迫,如高温、低温、干旱、盐等,导致正常生长发育受阻,从而影响产量和品质[24]。辣椒(Capsicum annuum)为一年生或多年生茄科(Solanaceae)作物,是全球种植面积最大的蔬菜作物和消费量最大的辛辣调味品[25]。辣椒喜温不耐热,最适生长温度为22-28℃,温度超过32℃,会产生不可逆热害症状,导致其生长发育不良,造成减产,严重制约辣椒产业发展[24]。目前在辣椒中还未见DNAJE基因家族的相关报道,辣椒全基因组测序数据[26]的公布为辣椒DNAJE基因家族的鉴定和分析提供了条件。
本研究对辣椒DNAJE基因家族进行鉴定,系统分析基因结构特点、进化关系、启动子顺式作用元件、共线性关系及不同组织器官的表达模式等,并研究其在高温胁迫下的生理生化和基因表达特征,为进一步探究辣椒DNAJE基因响应高温胁迫机制奠定理论基础。
1 材料与方法
1.1 材料
试验材料为课题组耐热育种骨干亲本862。挑选籽粒饱满一致的辣椒种子,播种于10 cm ×10 cm营养钵中,温室育苗。待幼苗长至5-6 片真叶时转入光照培养箱适应性生长,昼夜温度/光周期设定为26℃/12 h 和22℃/12 h,光照强度4 000 lx,每日定时浇水,控制空气相对湿度为70%。培养3 d 后调整培养箱昼夜温度至40℃/30℃,进行高温胁迫处理,对照组为26℃/22℃,光周期、光照强度、空气相对湿度保持不变。分别于处理0、1、3、5、7、10 d 时取叶片经液氮速冻后,-80℃保存备用。每次取样随机选择10 株,重复3 次。
1.2 方法
1.2.1 辣椒DNAJE基因家族鉴定和特征分析 从国家基因库核酸序列归档系统(CNSA)(https://db.cngb.org/cnsa/)和茄科基因组数据库(https://www.sgn.cornell.edu/) 分别下载辣椒Zunla 1 和番茄(Solanum lycopersicumL.)全基因组数据,构建本地蛋白数据库;从Pfam 数据库(https://pfam.xfam.org/)获得锌指(zinc finger)结构域DnaJ_CxxCxGxG HMM 模型文件PF00684,通过本地Hmmsearch 命令初步筛选获得DNAJE基因家族成员;参考已发表文献[8,10]从拟南芥数据库(https://www.arabidopsis.org/)获得33 个拟南芥DNAJE基因家族成员蛋白序列,与辣椒和番茄蛋白序列进行本地Blastp 比对,收集E-value<1e-5 输出基因,剔除重复序列后,与HMM 检索结果合并;将合并的候选序列上传至Pfam、NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)及SMART(http://smart.emblheidelberg.de/),基于DNAJE 结构域进一步比对筛选。利用ExPASy(https://web.expasy.org/compute_pi/)计算辣椒DNAJE 蛋白分子量和等电点,利用WoLF PSORT(https://wolfpsort.hgc.jp/)在线分析工具进行DNAJE 蛋白的亚细胞定位预测;利用在线软件SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测分析DNAJE 蛋白二级结构。
1.2.2 辣椒DNAJE基因家族染色体定位和比较进化关系分析 基于获取的辣椒基因组注释信息文件,利用TBtools[27]可视化工具构建辣椒染色体定位图;利用MEGA 5.0 软件的邻近法(neighbour-joining,NJ)构建辣椒、番茄和拟南芥DNAJE 进化树,设置Bootstrap 值为1 000,置换模型为P-distance,其他为默认参数;利用在线软件iTOL(http://itol.embl.de/)对进化树进行美化。
1.2.3 辣椒DNAJE基因家族保守基序、基因结构、启动子作用元件分析 利用MEME(https://memesuite.org/meme/tools/meme) 在线软件预测分析DNAJE 蛋白保守motif,motif 最大发现数设定为5个,其他参数默认。利用TBtools 软件分析DNAJE基因结构,利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线软件分析基因启动子顺式作用元件,利用MEGA 5.0 构建辣椒DNAJE基因家族系统进化树,方法参数同上,利用TBtools 软件对上述结果进行可视化。
1.2.4 辣椒和拟南芥DNAJE基因家族共线性及RNA-seq 分析 利用TBtools 软件中Fasta stats 和Table Row 功能构建辣椒和拟南芥染色体骨架、DNAJE基因位置,再利用其One step MCscanX(Super Fast)和Advanced circos 插件对辣椒和拟南芥进行共线性分析;从NCBI 数据库(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE45037) 下载叶、花、茎、根及不同时期果实的RNA-seq 数据,提取DNAJE基因家族成员TPM 值并绘制基因表达热图。
1.2.5 辣椒高温胁迫响应相关基因RT-qPCR 分析 利用FastPure Universal Plant Total RNA Isolation Kit(Vazyme)提取样品总RNA,通过琼脂糖凝胶电泳分析RNA 质量,NanoDrop 2000 超微量分光光度计检测RNA 浓度,利用Hiscript Ⅲ 1st strand cDNA Synthesis Kit(Vazyme)试剂反转录合成cDNA;利用Primer 5.0 设计特异性引物,并在NCBI Primer-BLAST(https://www.ncbi.nlm.nih.gov/tools/primerblast/)进行特异性验证,以UBI3为内参基因(表1)。参照ChamQ universal SYBR qPCR Master Mix(Vazyme)试剂操作说明书进行RT-qPCR 分析。采用2-△△Ct算法计算基因相对表达量,使用EXCEL、Origin 2021、SPSS 18.0 软件进行数据分析及作图。

表1 实时荧光定量PCR 引物Table 1 Primers used for quantitative real-time PCR
1.2.6 高温胁迫下辣椒幼苗的生理生化响应 超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)活性及丙二醛(malondialdehyde,MDA)、过氧化氢(hydrogen peroxide,H2O2)含量测定参照试剂盒说明书(苏州科铭生物技术有限公司)进行操作,采用Thermo MULTISKAN GO 型酶标仪分别测定不同波长下吸光度,计算酶活性或含量,利用EXCEL、Origin 2021 进行数据分析及作图。
2 结果
2.1 辣椒DNAJE基因家族鉴定与分析
本研究在Zunla 1 和番茄基因组中分别鉴定获得28 个和26 个DNAJE基因家族成员,依据其在染色体上的位置将辣椒DNAJE基因命名为CaDNAJE1-CaDNAJE28。对所有CaDNAJE 蛋白理化性质分析结果(附表1)显示,CaDNAJE基因序列开放阅读框(ORF)长度范围为297-1 410 bp,变化范围较大。CaDNAJE 蛋白氨基酸序列长度在99(CaDNAJE23)-470(CaDNAJE18)个之间,分子量变化范围为10.05(CaDNAJE23)-52.26(CaDNAJE18)kD,理论等电点(pI)为4.75(CaDNAJE7)-9.64(CaDNAJE20)。亚细胞定位预测发现,CaDNAJE 蛋白主要定位在叶绿体、细胞核及细胞质。二级结构分析表明(附表2),28 个CaDNAJE 蛋白二级结构完整,以无规卷曲(random coil)和α-螺旋(α-helix)为主。
2.2 辣椒DNAJE基因家族染色体定位和进化关系分析
28 个CaDNAJE基因不均匀分布在辣椒除7 号和8 号染色体之外的10 条染色体上,另外有3 个基因未锚定在染色体上(图1)。其中,1 号和9 号染色体上分布最多,均有5 个;3 号和11 号染色体上各包含3 个;2 号、6 号和10 号染色体上各有2 个;而4 号、5 号和12 号染色体上只有1 个成员。
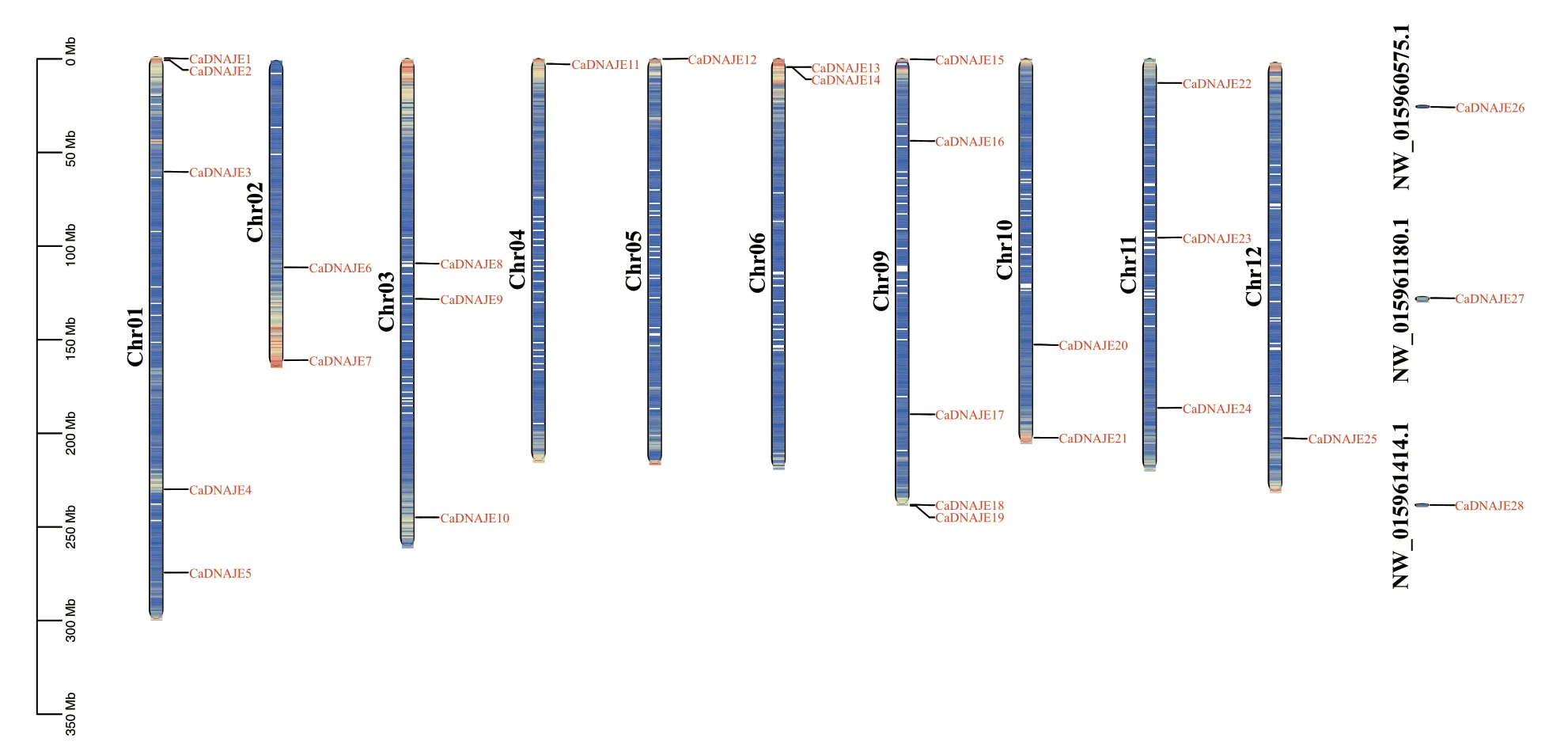
图1 辣椒DNAJE 基因家族成员染色体定位Fig.1 Chromosomal location of DNAJE gene family members in pepper(Capsicum annuum)
为探讨辣椒、番茄和拟南芥DNAJE基因家族成员之间的进化关系,构建了28 个辣椒、26 个番茄和33 个拟南芥DNAJE 蛋白的系统进化树(图2)。进化结果分为2 个亚族,Group Ⅰ和 Group Ⅱ。Group Ⅰ 有16 个辣椒成员、15 个番茄成员和20 个拟南芥成员,属于DNAJ CxxCxGxG 型DNAJE 锌指蛋白。Group Ⅱ 中有12 个辣椒成员、11 个番茄成员和13 个拟南芥成员,属于GRL CxxCx7CxxC 型DNAJE 锌指蛋白。
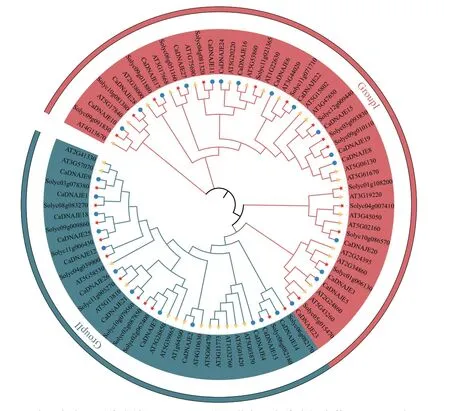
图2 辣椒、拟南芥和番茄DNAJE 基因家族成员进化树Fig.2 Phylogenetic tree of DNAJE gene family members in pepper,Arabidopsis and tomato(Solanum lycopersicum)
2.3 辣椒DNAJE基因家族成员保守基序、顺式作用元件及基因结构分析
基于MEME 在线分析软件,在CaDNAJE 蛋白中分析了5 个保守motif(图3-B),基序长度介于15-45 个氨基酸之间(表2)。系统进化树分析结果显示( 图3-A),Group Ⅰ 中CaDNAJE13 和CaDNAJE25 分别缺失motif 2 和motif 5 基序,其他10 个成员均包含motif 1-motif 5 保守基序;Group Ⅱ所有16 个成员均缺失motif 2、motif 3 和motif 5,只包含motif 1 和motif 4 保守基序。
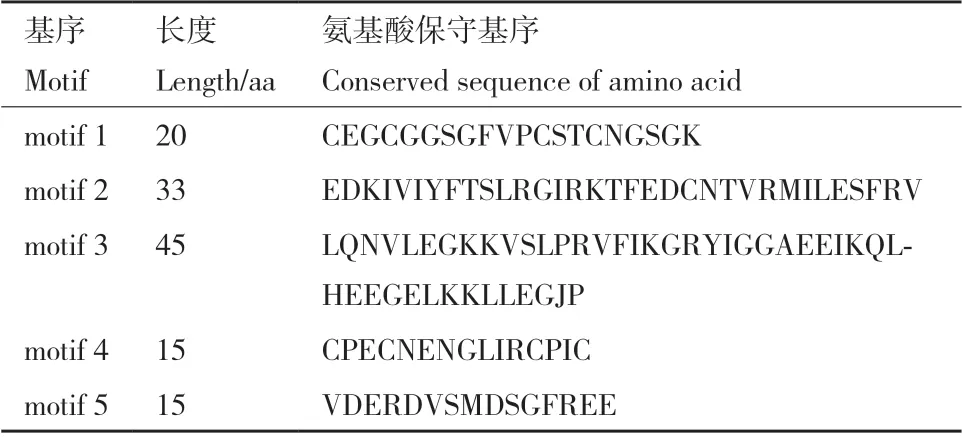
表2 辣椒DNAJE 蛋白氨基酸保守基序Table 2 Conserved motif of the DNAJE proteins in pepper
基因结构分析表明(图3-C),同一亚族内成员具有相似的基因结构,但不同亚家族间基因结构和内含子数目存在明显差异。Group Ⅰ 中除CaDNAJE2和CaDNAJE26外,均只包含1 个外显子,大部分成员无内含子;Group Ⅱ 中成员具有0-11 个内含子,其中具有4 个以上内含子的占比为56%,CaDNAJE19和CaDNAJE16内含子数目最多,CaDNAJE23和CaDNAJE20只包含1 个内含子,CaDNAJE27无内含子。
利用在线工具对该家族基因启动子顺式作用元件进行预测,结果如图3-D 所示。多个CaDNAJE基因启动子区富集了光响应(GATA-motif、G-box)、MeJA 响应(TGACG-motif)、脱落酸响应(ABRE)、水杨酸响应(TCA-element)、MYB 结合位点参与干旱诱导(MBS)、低温响应(LTR)、防御与胁迫响应(TC-rich repeat)、昼夜节律调节(circadian)等顺式作用元件。其中,TGACG-motif 和ABRE 较多,而MSA-like 和TGA-element 分别仅在CaDNAJE15和CaDNAJE20中被检测到。
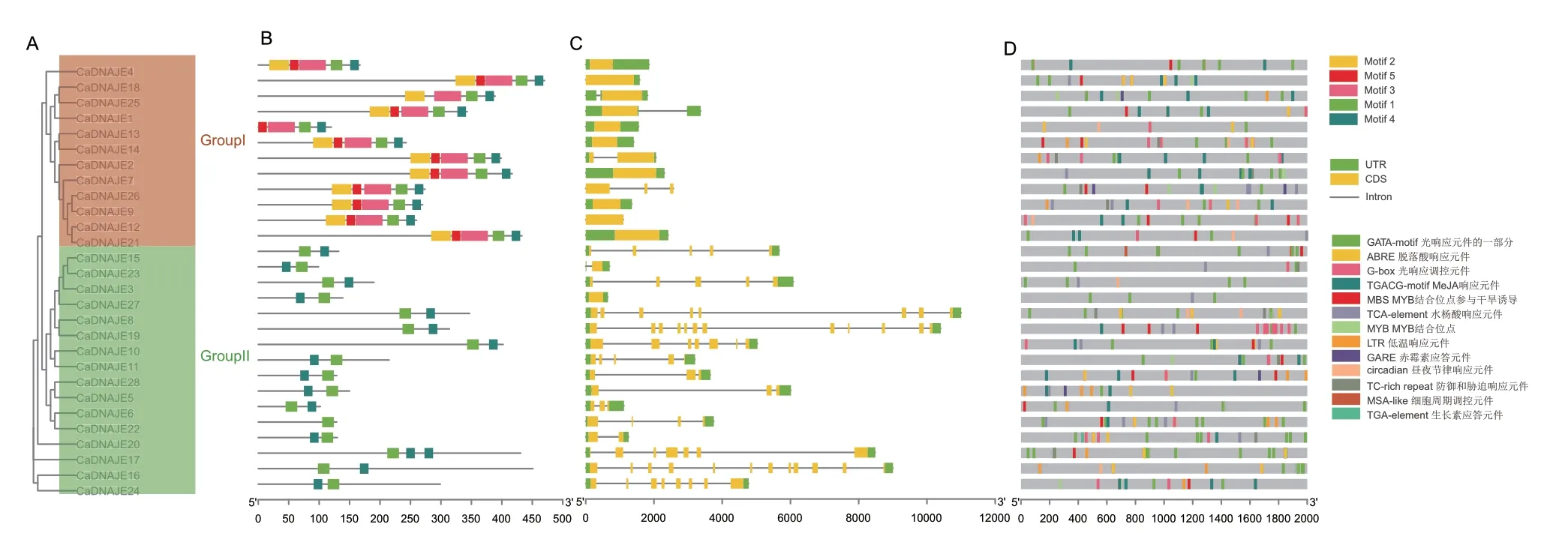
图3 辣椒DNAJE 基因家族保守基序、基因结构和顺式作用元件分析Fig.3 Analysis of conserved motifs,gene structure and cis-acting elements of DNAJE gene family in pepper
2.4 辣椒与拟南芥DNAJE基因家族成员共线性分析
共线性分析结果(图4)显示,辣椒和拟南芥之间有10 对DNAJE共线性基因,辣椒中9 个对应拟南芥中10 个DNAJE基因。分别为CaDNAJE1/AT4G10630、CaDNAJE4/AT5G06470、CaDNAJE4/AT3G11773、CaDNAJE7/AT3G28850、CaDNAJE8/AT5G61670、CaDNAJE10/AT4G13670、CaDNAJE11/AT1G75690、CaDNAJE13/AT5G58530、CaDNAJE18/AT5G03870、CaDNAJE21/AT2G41330 直系同源基因对。其中,CaDNAJE4基因在拟南芥中有2 个共线性基因,表明该基因发生了扩张。此外,在辣椒中仅有CaDNAJE12/CaDNAJE21之间存在共线性关系,表明该基因家族种内共线性基因较少。
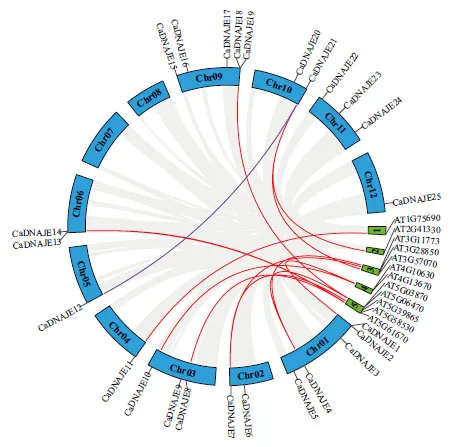
图4 辣椒和拟南芥DNAJE 基因家族共线性分析Fig.4 Collinearity analysis of DNAJE gene family between pepper and Arabidopsis
2.5 辣椒DNAJE基因家族表达模式分析
利用GEO Datasets 中的转录组数据,对辣椒DNAJE基因家族组织特异性表达模式进行分析,结果如图5。辣椒DNAJE基因在不同组织及果实中有明显表达差异,28 个基因可以分为4 类。CaDNAJE1、CaDNAJE4、CaDNAJE6等A 类基因在辣椒各组织中表达量都很低甚至不表达,多数为Group Ⅱ 基因;B 类4 个基因中仅CaDNAJE26在叶和花芽中表达量相对较高;C 类CaDNAJE 5、CaDNAJE 27、CaDNAJE 23等基因在花和幼果中高表达;D 类基因多数为Group Ⅰ 基因,除CaDNAJE2、CaDNAJE10、CaDNAJE20在0-1 cm 幼果或叶和花芽中几乎不表达外,其他基因在各组织中均高表达。

图5 DNAJE 基因在辣椒不同组织及果实发育过程中的表达分析Fig.5 Expression profile analysis of pepper DNAJE genes in different tissues and fruit development
2.6 辣椒高温胁迫响应基因的表达分析
通过分析NCBI SRA 数据库中辣椒高温胁迫转录组数据(SRP187794),对表达显著差异的12 个CaDNAJE基因和3 个Hsf/HSP 耐热相关基因进行高温胁迫RT-qPCR 分析(图6)。其中,CaHSP22、CaHSP16.4和CaHsfB5基因受高温胁迫表达水平极显著升高,随胁迫时间延长,CaHSP22、CaHSP16.4表达呈下降趋势;辣椒DNAJE基因中除CaDNAJE24负向调控辣椒响应高温胁迫,表达水平下调外,其他11 个基因均呈现不同程度的上调表达,但在不同胁迫时期的表达模式有所差异。CaDNAJE2、CaDNAJE3、CaDNAJE13和CaDNAJE21这4 个基因在1 d 时表达水平最高,之后呈下降趋势;CaDNAJE11基因在1、7 和10 d 时表达水平均显著高于对照,且在7 d 时表达水平最高;CaDNAJE15基因在胁迫1、3、5 d 时表达水平基本持平,之后降低;CaDNAJE17、CaDNAJE19、CaDNAJE23和CaDNAJE27这4 个基因表达基本呈现先升后降再升趋势,均在5 d 时达到最高。CaDNAJE基因在高温胁迫下的高表达,表明其作为应激蛋白可能在辣椒响应高温胁迫中起调节作用,以维持辣椒正常的生长发育。
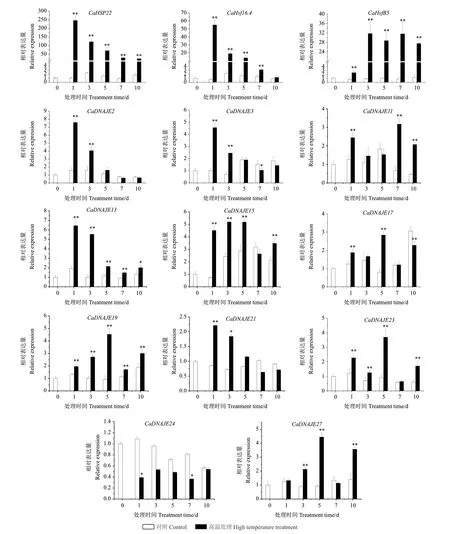
图6 辣椒高温胁迫相关基因表达分析Fig.6 Expression analysis of genes related to high temperature stress in pepper
2.7 高温胁迫下辣椒幼苗的生理生化响应
当植物遭遇高温等逆境胁迫时,会造成体内H2O2积累,威胁植物细胞膜系统,导致膜脂过氧化产物MDA 含量升高。同时,植物自身SOD、POD、CAT 等抗氧化保护酶系统启动,参与植物对外界逆境胁迫的响应。由图7可知,随着高温胁迫时间延长,H2O2含量急剧升高,5 d 达到最高的5.36 μmol/g,之后开始下降;MDA 含量呈上升趋势,10 d 时比处理前增幅437.01%;SOD 酶活性呈先升后降趋势,在3 d 时活性达到顶峰的188.0 U/g,相比对照,3-10 d 酶活性升高幅度为24.7%-66.1%;POD 酶活性整体呈上升趋势,在10 d 时达到最高的3 819.88 U/g,相比对照,3 d 时活性提升幅度最大,为75.5%;CAT 酶活性前期变化不大,但在10 d 时酶活性明显升高,达到276.91 nmol/(min·g),为对照(40.25 nmol/(min·g))的6.9 倍。

图7 辣椒幼苗对高温胁迫的生理生化响应Fig.7 Physiological and biochemical responses of pepper seedlings to high temperature stress
3 讨论
近年来,随着辣椒全基因组数据的公布[26-28]和生物信息学方法的不断完善,研究人员已对辣椒DnaJ[29]、B-BOX[30]、MYB[31]、AP2/ERF[32]等多个基因家族进行了鉴定分析,但辣椒DNAJE基因家族系统鉴定和分析还未见报道。本研究基于Zunla 1全基因组数据,对辣椒DNAJE基因家族进行鉴定,从基因结构特征、系统进化、顺式作用元件、共线性关系、组织特异性表达及高温胁迫响应等多方面进行了分析和研究。
3.1 植物DNAJE基因功能分析
HSP70 是植物主要伴侣蛋白家族之一,在蛋白质量控制中起非常重要的作用。DnaJ 蛋白(HSP40)是HSP70 的共同伴侣,他能识别未折叠的底物并将其传递给HSP70,刺激ATP 酶活性,诱导伴侣构象改变,进而稳定其与底物的相互作用[33]。DNAJE 蛋白拥有类似于DnaJ 蛋白中参与结合底物的锌指结构域[34],而缺乏典型的J-结构域,能够不依赖HSP70和J-结构域,直接与底物结合发挥作用。已有研究证实,ORANGE 蛋白作为DNAJE 蛋白家族一员,通过转录后调控植物烯合成酶PSY 以及通过抑制细胞核中转录因子TCP14 活性调控色素合成和叶绿体发育[35-37]。本研究在辣椒中鉴定出28 个DNAJE基因,进化分析显示,辣椒、番茄和拟南芥DNAJE 蛋白被划分至2 个亚族,每个亚族中辣椒、番茄和拟南芥成员分布情况相似,说明其具有较近的同源关系。启动子顺式作用元件对基因转录及调控具有非常重要的作用[38]。分析发现,DNAJE基因启动子区域包含大量光响应、激素响应和胁迫应答元件。结果表明,DNAJE基因可能不仅参与光系统调控,也参与了激素响应和胁迫应答。诸多研究已经证实了这点,Amiya 等[13]报道了一个莱茵衣藻DNAJE 类囊体相关蛋白ZnJ6,其能与氧化还原酶、光合蛋白等相互作用,防止因外界环境胁迫使蛋白发生错误折叠和聚集,从而维持氧化还原和光合平衡,提升抗逆性。Hartings 等[39]在拟南芥中分离了HCF222基因,具有二硫化物还原酶活性,参与细胞色素b6f 组装,进而影响PSII 和PSI 光合复合物的合成;Ham 等[40]通过对烟草(Nicotiana tabacum)DNAJE基因Tsip1的研究发现,其能够与Tsi1转录因子互作,激活下游水杨酸反应基因,进而参与烟草抗逆胁迫应答反应。通过共线性分析发现,辣椒和拟南芥具有10 对直系同源基因,且CaDANJE1/AT4G10630、CaDNAJE8/AT5G61670、CaDNAJE10/AT4G13670、CaDNAJE11/AT1G75690、CaDNAJE13/AT5G58530 这5 对不仅进化关系最近,且序列相似度很高。其中,AT5G61670、AT4G13670 和AT1G75690 已分别被鉴定为拟南芥OR[41-42]、pTAC5[43]和LQY1[44]基因,在质体发育、逆境应答及光系统建成等方面发挥重要作用,因此推测辣椒近源基因也具有类似功能。
3.2 辣椒幼苗响应高温胁迫相关基因表达分析
已有研究表明,热激转录因子Hsf 和热激蛋白HSP 是参与植物应激反应的主要通路[45],CaHsfB5[46]、CaHSP22[47]和CaHSP16.4[48]等基因在辣椒响应高温胁迫中显著上调表达,进而提升辣椒耐热性,这与本研究结果一致。DNAJE 蛋白与典型DanJ 蛋白具有相似的功能,其表达也能够显著提高植物对非生物胁迫的耐受力,从而降低细胞损伤[49]。Lee 等[50]在苜蓿(Medicago sativaL.)中鉴定了一个DNAJE基因MsDJLP,该基因的表达受低温(4℃)和高温(42℃)诱导,通过在烟草中异位过表达MsDJLP基因增强了转基因烟草对低温和高温的耐受力。So 等[51]通过对大豆(Glycine maxL.)DNAJE基因GmDjp1的研究发现,其受多种非生物胁迫诱导,在大肠杆菌中异源表达GmDjp1基因,能够提高大肠杆菌的耐受性,表明GmDjp1可能在细胞热休克应激过程中发挥关键作用。本研究对NCBI SRA 数据库中辣椒高温胁迫转录组数据进行分析,从2 个亚族中挑选出12 个在高温胁迫下表达量具有显著差异的CaDNAJE基因进行RT-qPCR 分析,结果表明,CaDNAJE24在高温胁迫中表达量降低,呈负调控,其他基因表达水平均显著提高,但其在不同胁迫时间的表达量存在差异,这反映出DnaJ-Like基因以不同调控方式参与了植物响应高温胁迫进程。
4 结论
在辣椒全基因组中共鉴定出28 个DNAJE家族基因,进化分析结果分为2 个亚族,同一亚族具有相似的蛋白保守基序、基因结构以及组织特异性表达模式。高温胁迫导致辣椒生理生化改变,并诱导DNAJE基因高表达,表明该家族基因参与了辣椒响应非生物胁迫的生物过程。
本文所有附表数据请到本刊官网下载(http://biotech.aiijournal.com/CN/1002-5464/home.shtml)。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
科学与财富(2021年3期)2021-03-08 10:56:02
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13 01:45:42
温州大学学报(自然科学版)(2019年2期)2019-06-04 11:52:00
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
