
沙门氏菌烈性噬菌体JN-S202001的分离、鉴定及其应用于即食鸭脖的抑菌效果
2023-01-31 09:22赵远洋吴国平
江西农业大学学报 2022年6期
赵远洋,舒 梅,吴国平,毕 妍,潘 红,钟 婵
(江西农业大学 食品科学与工程学院,江西 南昌 330045)
【研究意义】近年来食源性病原菌引起的感染事件逐年增加,是全球公共卫生关注的焦点问题[1]。全球每年约15 亿腹泻病人,其中70%是由污染食源性致病菌食品引起。我国仅2021 年5 月就有食物中毒事件28 起,共963 例,其中微生物性食物中毒事件9 起,共593 例,分别占32.14%和61.58%[2]。沙门氏菌(Salmonella)在自然界中血清型众多,人和温血动物的肠道是其主要栖息环境,并可通过其排泄物污染环境甚至进入食品链,造成食源性疾病的传播[3]。15~20 CFU 低剂量沙门氏菌(Salmonella)就可能导致腹痛、腹泻、呕吐等症状,严重者可造成死亡。大多数由沙门氏菌引起的感染归因于食用了被污染的禽畜肉、鸡蛋、牛奶、生菜等食品[4]。王赞等[5]抽检2018 年6-10 月河北省部分地区市售猪肉馅及冷鲜鸡食品,共检测出沙门氏菌阳性样品144 份,阳性检出率高达63%。2019 年10 月一起食用肠炎沙门氏菌污染糕点引起596 个病例[6]。随着抗生素的滥用,大量抗生素通过禽畜粪便和食物链释放到环境中,导致抗生素耐药细菌及其耐药基因的选择和传播[7]。因此亟须开发防控食源性致病菌的新型抗菌剂。【前人研究进展】噬菌体(phage)是一类专门感染细菌的病毒,1915 年和1917 年由TWORT 和d’Herelle 发现,由于其具有自我复制、特异性高、环境中含量丰富等优点,可被用于治疗细菌感染[8-9]。此外,口服噬菌体鸡尾酒对健康人体受试者无副作用,未观察到荨麻疹或过敏症状,与口服安慰剂的对照组相比,试验组的血液理化指标和肠道菌群结构无显著差异,显示噬菌体对人体具有良好的安全性[10]。有研究表明,烈性噬菌体可以有效降低食品中受污染的沙门氏菌含量[11-12]。Atterbury 等[13]从屠宰场、家禽养殖场废水中分离3 株沙门氏菌噬菌体,其应用可在24 h 内使盲肠内溶物中肠炎沙门氏菌降低约2.19~4.2 Log10CFU/g。Duc 等[14]从鸡皮、鸡胃中分离得到的5 株噬菌体组成鸡尾酒,处理鼠伤寒沙门氏菌模拟污染肠炎鸡肉样品,8oC 和25 ℃条件下放置,沙门氏菌分别降低1.86 Log10CFU/块和2.21 Log10CFU/块。烈性噬菌体也可用于控制多重抗生素耐药细菌的生长。Zhao 等[15]研究表明,利用4 种噬菌体处理胡萝卜种植土壤中耐四环素大肠杆菌和耐氯霉素铜绿假单胞菌,显著降低了该系统中病原菌和相应抗生素抗性基因的丰度。【本研究切入点】即食性卤制鸭脖具有独特的风味和口感,深受消费者喜爱,但由于其加工、流通环节多,且不再加热而直接食用,增加致病菌污染引起的食品安全风险[16-17]。沈托等[18]在2011—2019 年调查发现,熟肉制品中沙门氏菌的污染率为3.91%,路边摊点食品沙门氏菌阳性检出率更高(7.84%)。因此,开发噬菌体抑菌剂有望用于食品中沙门氏菌污染防控,对提高食品卫生安全具有理论参考价值和实践应用前景。【拟解决的关键问题】利用本研究室从食品中分离的沙门氏菌作为宿主,从生活污水中分离纯化沙门氏菌烈性噬菌体,对噬菌体进行生物学鉴定、全基因组测序,并初步测定其用于即食鸭脖污染沙门氏菌的抑菌效果,以期为新型天然噬菌体抑菌剂的研制开发提供依据。
1 材料与方法
1.1 材料与试剂
沙门氏菌菌株:本研究室2018—2019年分离来自南昌市农贸市场的鸡肉和猪肉等食品,经理化和分子生物学鉴定,但未鉴定血清型;沙门氏菌(ATCC BAA-708)及大肠杆菌O157:H7(9490)菌株由美国麻省大学食品科学系Robert E.Levin教授馈赠;沙门氏菌(BNCC185946、BNCC336875)购自北纳生物有限公司;以上菌株均保存于本研究室-80oC冰箱。污水样品为江西农业大学菜市场附近的生活污水池塘。即食卤制鸭脖为市售新鲜食品。
LB 营养肉汤培养基(北京陆桥技术有限责任公司);PEG 8000、3%磷钨酸负染色液(北京索莱宝科技有限公司);DNase I,RNase A(日本TaKaRa 公司);柱式DNA 核酸提取试剂盒(哈尔滨元亨生物药业有限公司);镁盐(saline magnesium,SM)缓冲液。
1.2 仪器与设备
HFsafe-1200LC生物安全柜(上海力申科学仪器有限公司);UV 6100A紫外可见分光光度计(上海元析仪器有限公司);pHS-3E pH 计(上海佑科仪器仪表有限公司);TS-2102C 恒温摇床(上海天呈实验仪器制造有限司);SHP-160 型生化培养箱(上海三发科学仪器有限公司);GL-21M 高速冷冻离心机(湖南湘仪仪器开发有限公司)。
1.3 试验方法
1.3.1 细菌培养 参考何学文等[19]方法,将活化后的沙门氏菌于37 ℃、180 r/min条件下培养过夜后,以3%的比例转接至10 mL LB培养液中,37 ℃,180 r/min培养至指数期(OD600nm≈0.5),取1 mL菌液,4 ℃,12 000 r/min离心10 min,沉淀重悬于1 mL灭菌生理盐水备用。
1.3.2 噬菌体的分离、纯化 参考文献[11]的分离方法,2020 年11 月采自江西农业大学菜市场附近生活污水500 mL,加氯化钙至终浓度1 mmol/L,充分混匀后室温静置2 h。4 ℃,3 000 r/min 离心10 min,上清经0.22 μm 滤膜过滤。取滤液50 mL,加等体积2×LB 液体培养基,再加指数期沙门氏菌3 mL,37 ℃,180 r/min 培养过夜。将培养液4 ℃,5 000 r/min,离心10 min。上清液经0.22 μm滤膜过滤后,滤液即为噬菌体原液。
将噬菌体原液用SM 缓冲液按10倍梯度稀释至适当浓度。取稀释液100 μL与等体积的指数期沙门氏菌混匀,静置15 min 后与7 mL 的LB 半固体培养基混匀倒入LB 固体培养基上,冷却后37 ℃倒置培养直至出现噬菌斑。
用无菌枪头挑取大而透明的噬菌斑于1 mL SM 缓冲液中,37 ℃,200 r/min 震荡过夜。次日将SM 缓冲液4 ℃,12 000 r/min 离心10 min,取上清液100 μL 与等体积指数期沙门氏菌混匀后加入5 mL LB 液体培养基中,37 ℃,200 r/min培养至液体澄清。培养液经高速离心,取上清液经0.22 μm滤膜过滤。滤液用SM 缓冲液10 倍梯度稀释制双层平板,37 ℃倒置培养过夜后挑取单个噬菌斑。重复上述操作3~5 次,直至噬菌斑的形态大小基本一致。双层平板法测定噬菌体效价,每个梯度重复3次。
1.3.3 噬菌体最佳感染复数(multiplicity of infection,MOI)参考Wang 等[20]的方法,调整沙门氏菌浓度至1×108CFU/mL。用SM缓冲液调节噬菌体效价,使最终噬菌体的感染复数为100~0.000 001。分别取噬菌体液和宿主菌液各500 μL混匀静置15 min后加入LB液体培养基5 mL,37 ℃,200 r/min培养4 h。双层平板法计算噬菌体效价,每组3个平行,效价最高的为最佳感染复数。
1.3.4 噬菌体一步生长曲线 参考高明明等[21]的方法,取噬菌体液与指数期沙门氏菌各1 mL 按最佳MOI(MOI=0.01)混匀,静置10 min 后12 000 r/min 离心2 min,沉淀用LB 液体培养基洗涤2 次后重悬于37 ℃的10 mL LB 液体培养基中。37 ℃,200 r/min 培养185 min,在0,2,5 min 时分别取样,5~15 min 时每隔5 min取样1次,15~45 min时每隔10 min取样1次,45~185 min时每隔20 min取样1次。样品离心过滤后上清液用双层平板法计算噬菌体效价,每组重复3次。
1.3.5 噬菌体裂解谱 采用成斑效率(efficiency of plating,EOP)法测定噬菌体裂解谱。EOP值表示噬菌体感染不同细菌后产生子代噬菌体的能力。EOP值越高则噬菌体对该细菌的裂解能力越强:EOP≥0.5为高裂解能力,0.1≤EOP<0.5为中等裂解能力,0.001≤EOP<0.1为低裂解能力,EOP≤0.001认为无裂解能力[22]。
1.3.6 噬菌体pH 稳定性和热稳定性 参考侯忠余等[23]的方法,将效价约109PFU/mL 的噬菌体液分装至12 个1.5 mL 无菌离心管中,每管300 μL,于30~80 ℃的水浴锅中水浴1 h,随后以双层平板法测定其效价,每组实验重复3次。用HCl和NaOH 调节SM 缓冲液pH 至2~13,分别取100 μL 噬菌体液加入900 μL各pH值的SM缓冲液中,37 °C水浴1 h,随后以双层平板法测定其效价,每组实验重复3次。
1.3.7 噬菌体电镜观察 参考Li 等[24]的方法,富集后的噬菌体悬液加入DNase Ι 和RNase A 至终浓度为1 μg/mL,37 ℃水浴1 h。然后加入终浓度为1 mol/L 的氯化钠,混匀后冰浴2 h,4 ℃,12 000 r/min 离心15 min,取上清液加入终浓度为10%的PEG 8 000,冰浴搅拌过夜,使噬菌体充分沉淀。次日4 ℃,15 000 r/min离心30 min,弃上清液,沉淀用2 mL SM缓冲液重悬,双层平板法测定效价。浓缩后的噬菌体液用无明胶的SM 缓冲液稀释至109PFU/mL。铜网在噬菌体液中浸泡5 min 后取出置于滤纸上干燥2 min,再将铜网放入3%磷钨酸中浸泡2 min,取出干燥5 min,在透射电镜下观察。
1.3.8 噬菌体的核酸提取和基因组测序 根据柱式DNA核酸提取试剂盒说明书提取核酸。将噬菌体核酸样品交由上海翰宇生物科技有限公司进行全基因组测序。用BLAST-N(NCBI)分析核酸序列。使用RAST(https://rast.nmpdr.org/)预测并注释开放阅读框架(ORF)。噬菌体基因组图谱由CGviewServer(http://cgview.ca/)生成。利用抗生素耐药性基因数据库(http://arpcard.mcmaster.ca)和毒力因子数据库(http://www.mgc.ac.cn/VFs/)进行抗生素耐药性和毒力因子相关基因的筛选。使用MEGA Version 7.0 软件以2 000次bootstrap重复的UPGMA方法构建系统发育树[24]。
1.3.9 噬菌体对即食性鸭脖中沙门氏菌的抑制作用 参考Li等[24]的方法,将100 μL噬菌体悬液(终浓度为108PFU/g)接种到10 g 沙门氏菌阴性的市售新鲜即食卤制鸭脖,以SM 缓冲液处理作为对照组。然后分别加入终浓度为102CFU/g 和104CFU/g 的沙门氏菌。样品在室温下静置10 min 晾干表面水分后,在4 ℃条件下保存48 h。每24 h采集样品1次,稀释至适当倍数后取100 μL 涂布于SS琼脂培养基上,37 ℃倒置过夜培养后计数沙门氏菌落总数。
1.3.10 数据计算方法 噬菌体效价与成斑效率EOP计算公式如下:

2 结果与分析
2.1 噬菌体分离纯化及透射电镜观察
以沙门氏菌2089b 为宿主菌,从生活废水中分离到一株烈性沙门氏菌噬菌体,命名为JN-S202001。双层平板上形成的噬菌斑形态大小基本一致,斑块透明且不带晕圈(图1A)。噬菌体效价为9.0×109PFU/mL。透射电镜观察结果表明,JN-S202001具有直径约73 nm呈立体对称结构的头部,长约233 nm不收缩且柔韧的尾部,符合有尾噬菌体目长尾噬菌体科特征(图1B)。

图1 JN-S202001双层平板噬菌斑(A)和透射电镜下形态(B)Fig.1 Plaques in a double-layer agar plate(A)and transmission electron micrographs(B)of JN-S202001
2.2 噬菌体最佳感染复数(MOI)及一步生长曲线测定
不同MOI 下噬菌体的效价见表1。MOI 由100 降至0.01 时,噬菌体效价不断升高,在MOI=0.01 时达到最大值7.5×1010PFU/mL。表明JN-S202001最佳MOI为0.01。

表1 JN-S202001最佳感染复数测定Tab.1 Determination of the optimal MOI of JN-S202001
一步生长曲线结果显示,JN-S202001 潜伏期较短,为5 min,爆发期持续时间较长,共80 min,在85 min 左右达到稳定期,裂解量为裂解末期噬菌体效价(1.6×109PFU/mL)与感染初期宿主菌量(2.4×107CFU/mL)之比,约67 PFU/cell(图2)。

图2 JN-S202001的一步生长曲线Fig.2 One-step growth curve of JN-S202001
2.3 噬菌体热稳定性和pH稳定性测定
噬菌体在30~50 ℃均可保持较高效价(>108PFU/mL),60 ℃时下降约2.0 log10PFU/mL。70 ℃时只有少量噬菌体存活(1.9 log10PFU/mL),温度达到80 ℃时噬菌体全部失活(图3A)。在pH 2~11 内噬菌体效价较为稳定,维持在108PFU/mL 左右,当pH=12 时噬菌体的效价略有下降,在pH=1 和pH=13 的环境中噬菌体全部失活(图3B)。
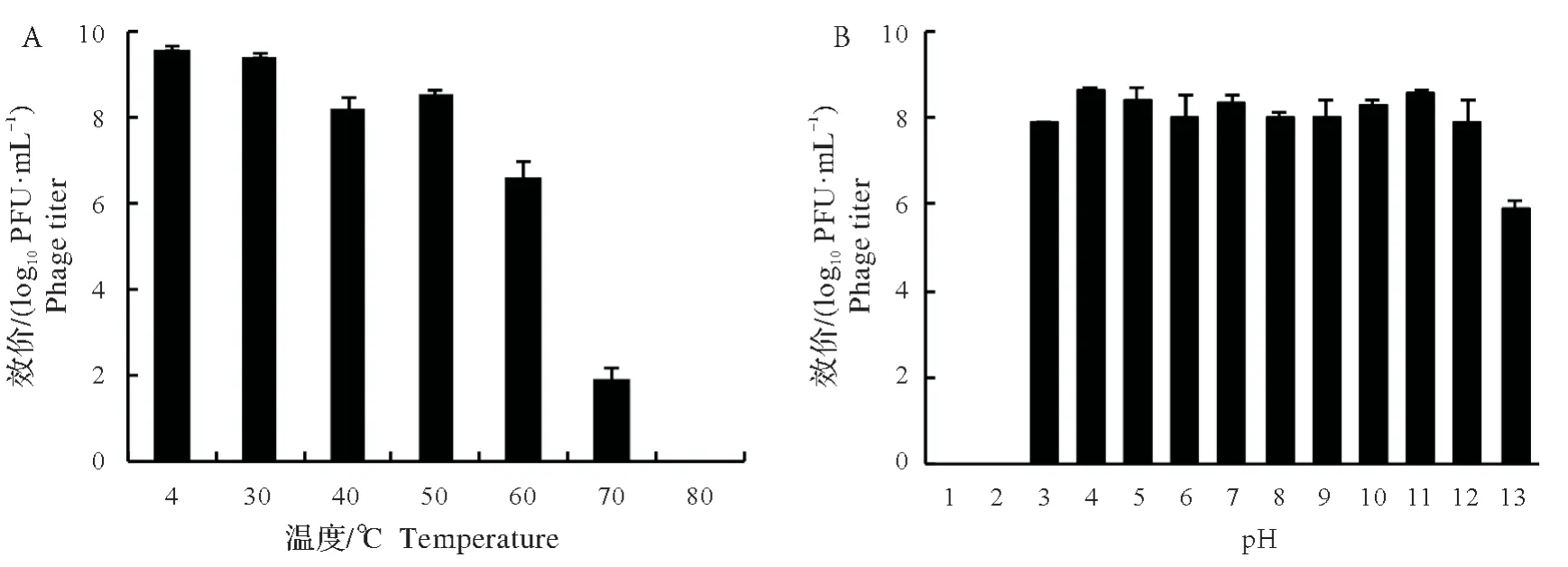
图3 JN-S202001的热稳定性(A)和pH稳定性(B)Fig.3 Thermal(A)and pH(B)stability of JN-S202001
2.4 噬菌体裂解谱测定
JN-S202001 对42 株沙门氏菌中的9 株具有裂解能力,裂解比例为21.4%。在可裂解的9 株沙门氏菌中,7 株为多重抗生素耐药菌株,4株对8种抗生素具有耐药性,3株对4种抗生素具有耐药性(表2)。该噬菌体对实验室保存的一株大肠杆菌O157:H7 和金黄色葡萄球菌无裂解能力。结果表明,JN-S202001 能够裂解受试沙门氏菌株,包括多重抗生素耐药菌株。

表2 JN-S202001裂解谱测定Tab.2 Determination of host ranges of JN-S202001
2.5 JN-S202001的基因组序列及系统发育树分析
JN-S202001 的核酸可以被DNase I 消化,但不被RNaseA 消化,表明其基因组是DNA 分子。全基因组序列分析表明,JN-S202001 的基因组由111 256 bp 的双链DNA 组成,G+C 含量约为40%(GenBank 登录号:OL502173)。编码184个ORF(正链138个,负链46个),包括24个tRNA 基因和47个与已报道的功能基因有显著同源性的编码蛋白基因。氨基酸序列同源性分析表明,JN-S202001 的基因组没有编码毒力和抗生素耐药性的基因,且未检测到与噬菌体溶原性相关的基因。BLAST-N 分析表明,JN-S202001基因组与长尾噬菌体科Demerecviridae亚科噬菌体沙门氏菌噬菌体SP76同源性最高,达99.07%。
利用JN-S202001的裂解酶氨基酸序列构建系统发育进化树(图5),结果表明,JN-S202001与沙门氏菌噬菌体100268_sal2(NCBI登录号:NC_031902.1)同源性最高。
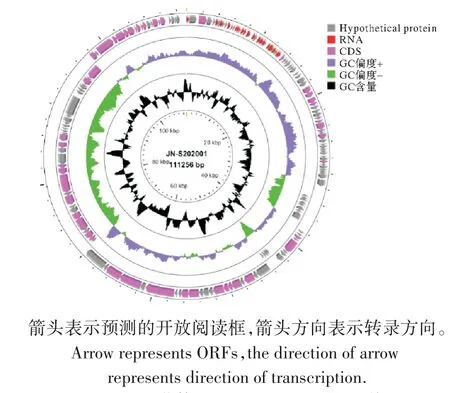
图4 噬菌体JN-S202001基因组图谱Fig.4 Genomic map of phage JN-S202001

图5 JN-S202001裂解酶蛋白发育进化树Fig.5 The phylogenetic tree of JN-S202001 Lysin
2.6 JN-S202001对即食鸭脖污染的沙门氏菌抑制效果
未经JN-S202001 处理的鸭脖污染终浓度为2.0 Log10CFU/g 的沙门氏菌,4 ℃储存48 h 后沙门氏菌浓度降至1.3 Log10CFU/g;而JN-S202001 处理组在4 ℃储存48 h 后未检测到沙门氏菌(P<0.01,t(4)=87.64)(图6A)。将终浓度4.1 Log10CFU/g的沙门氏菌接种于鸭脖,对照组沙门氏菌浓度在4 ℃储存48 h后降至3.3 Log10CFU/g;JN-S202001 处理组在储存0 h 时沙门氏菌降至3.3 Log10CFU/g,储存48 h 后沙门氏菌降至2.0 Log10CFU/g,与对照组相比差异极显著(P<0.01,t(4)=35.03)(图6B)。
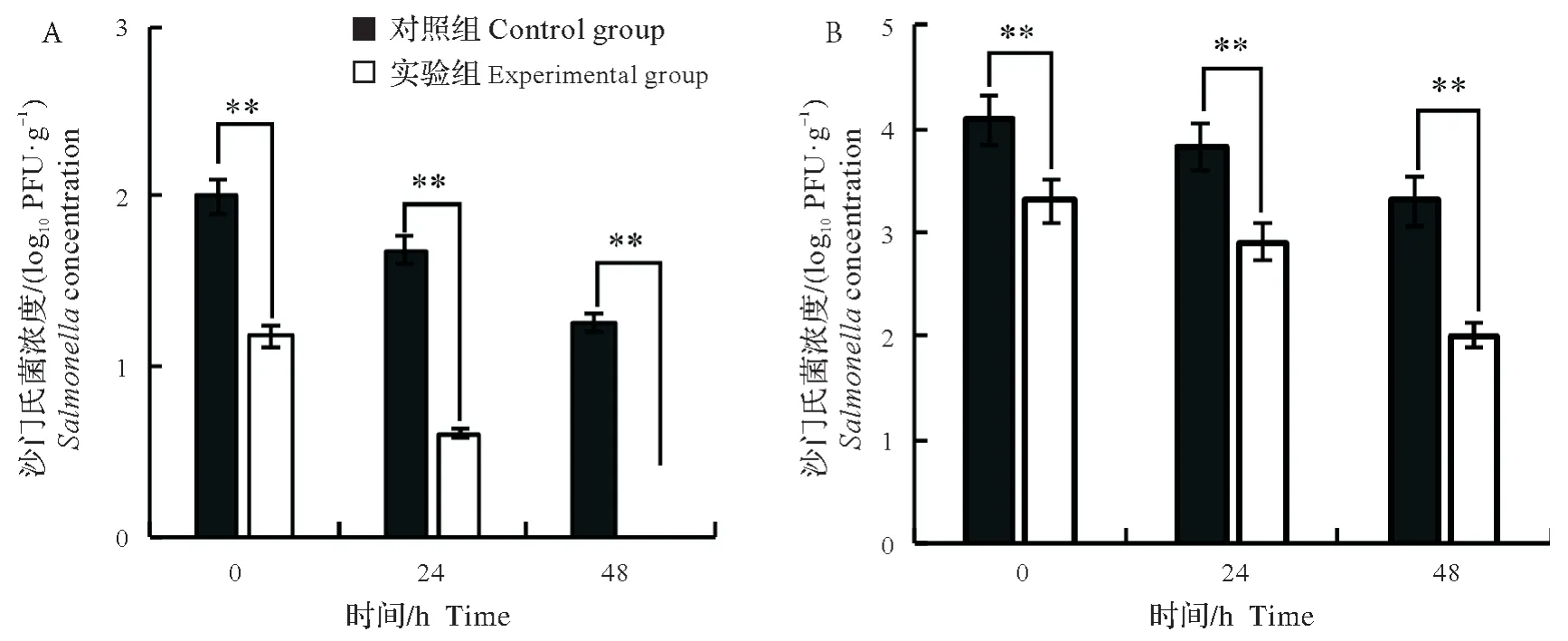
图6 JN-S202001对4 ℃保存48 h的即食鸭脖污染的沙门氏菌抑菌效果Fig.6 Antibacterial effect of JN-S202001 in ready-to-eat duck neck at 4 °C for 48 h
3 结论与讨论
本研究从生活废水中成功分离和鉴定了一株沙门氏菌的特异性噬菌体JN-S202001。根据其形态特征和基因组序列,推测JN-S202001 属于有尾噬菌体目长尾噬菌体科。一步生长曲线结果表明,JNS202001的潜伏期和裂解期分别为5 min和80 min,裂解量约67 PFU/细胞,与李萌[25]和聂若男等[26]分离的噬菌体裂解性相似。JN-S202001 对7 株多重抗生素耐药沙门氏菌均表现出裂解活性,表明噬菌体可用于抑制多重抗生素耐药细菌。噬菌体成功感染宿主需要首先吸附到宿主的特异性受体上,革兰氏阴性菌噬菌体常以脂多糖、外膜蛋白、鞭毛、O-抗原等作为受体[27]。沙门氏菌O-抗原具有多样性,是划分沙门氏菌血清型的重要依据,JN-S202001对受试菌株中的沙门氏菌具有感染性,表明该噬菌体可能以某种O-抗原作为受体,具体吸附机制有待后续进一步研究。
JN-S202001 在较高的温度(<60 ℃)和较宽pH(2~11)内保持活性稳定。李萌[25]以鼠伤寒沙门氏菌ATCC14028 为宿主分离出一株烈性噬菌体,在温度为40~50 ℃,pH 4~10 内均可保持较高裂解能力(>109PFU/mL)。包细明[28]分离的噬菌体BpS 在pH 5~10 时效价比较稳定,保持在109PFU/mL 以上,但其温度适应范围较窄,在温度低于50 ℃时效价比较稳定,存活率高于88%,温度高于60 ℃时噬菌体存活率仅20%左右。聂若男等[26]从污水中分离的一株烈性短尾沙门氏菌噬菌体T139,在30~50 ℃和pH 4~12条件下效价相对稳定。JN-S202001对温度和pH具有较宽的适应范围,在酸性条件下(pH 2~4)仍能保持较高的裂解能力,显示具有良好的应用潜力。
全基因组测序和注释表明,JN-S202001的基因组全长111 256 bp,编码184个开放阅读框,不含毒力基因和耐药基因。预测的蛋白质编码序列(CDS)与已报道的全基因组基因的同源性表明,JN-S202001与有尾噬菌体目Demerecviridae 亚科噬菌体的沙门氏菌噬菌体SP76相似性最高。基因组注释表明,JNS202001 具有编码宿主裂解蛋白、DNA 复制/修饰/调节和包装蛋白、尾部结构蛋白及一些附加功能蛋白的所有基本结构和功能基因。噬菌体侵染革兰氏阴性菌的机制通常为宿主内膜(IM)被穿孔素通透形成跨膜孔,使裂解酶从细胞质中逸出至细胞壁肽聚糖层,从而降解肽聚糖发挥裂解功能[29]。JN-S202001编码的holin(ORF8)、endolysin(ORF9),推测与裂解活性有关,其裂解特性与已报道的革兰氏阴性菌噬菌体裂解机制一致。此外,endolysin(ORF9)系统进化树(图5)分析表明,JN-S202001 与沙门氏菌噬菌体100268_sal2同源性最高。
模拟沙门氏菌污染即食鸭脖,与未经JN-S202001 处理对照组相比,样品在4 ℃保存48 h 后,JNS202001 极显著(P<0.01)降低了沙门氏菌含量(>2.0 Log10CFU/g)(图6)。Phothaworn 等[30]将分离的鼠伤寒沙门氏菌噬菌体ST-W77 和肠炎沙门氏菌噬菌体SE-W109 混合添加到污染沙门氏菌的牛奶或鸡肉中,结果表明能够将样品的抑菌效果延长一周。Abhisingha 等[31]研究了噬菌体鸡尾酒在低温(4 °C和-20 °C)贮藏的鸡肉中对沙门氏菌的抑制效果,结果表明在4 ℃下贮藏72 h后,噬菌体鸡尾酒处理组的沙门氏菌数量发生显著降低(0.4~1.0 Log10CFU/cm2,P<0.05);在-20°C条件下,0~24 h内沙门氏菌数量显著降低0.4~0.7 Log10CFU/cm2(P<0.05)。综上,噬菌体JN-S202001 有望作为一种潜在的生物抑菌剂用于食品中沙门氏菌污染的防控。
致谢:江西省科学技术支撑项目(20212BBF63027,20212BAB215018)和江西省现代农业产业技术体系建设专项资金资助(JXARS-12)同时对本研究给予了资助,谨致谢意!
猜你喜欢
中国食品学报(2022年6期)2022-07-19
心理学探新(2022年1期)2022-06-07
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
中国饲料(2021年17期)2021-11-02
食品安全导刊(2021年20期)2021-08-30
科学24小时(2020年4期)2020-05-14
猪业科学(2018年5期)2018-07-17
中国当代医药(2015年8期)2015-03-01
中成药(2014年9期)2014-02-28
