
敲除OsNramp5基因创制低镉优质粳稻新材料的应用评价
2023-01-18 12:32裴峰王广达高鹏冯志明胡珂鸣陈宗祥陈红旗崔傲左示敏
中国水稻科学 2023年1期
裴峰 王广达 高鹏 冯志明 胡珂鸣 陈宗祥 陈红旗 崔傲 左示敏 ,
(1扬州大学 江苏省作物基因组学和分子育种重点实验室/植物功能基因组学教育部重点实验室,江苏 扬州 225009;2扬州大学 江苏省粮食作物现代产业技术协同创新中心/江苏省作物遗传生理重点实验室,江苏 扬州 225009;3中国水稻研究所 水稻生物学国家重点实验室,杭州 311400;4镇江禾下土农业科技有限公司,江苏 镇江 212008;5扬州大学 教育部农业与农产品安全国际合作联合实验室,江苏 扬州 225009;*通信联系人,email:smzuo@yzu.edu.cn)
水稻是我国三大主粮之一,全国有60%以上人口以稻米为食。民以食为天,保证稻米供给安全对于保证国家粮食安全至关重要[1]。2011年以来,我国人均稻谷占有量达152 kg[2],温饱问题已基本解决,在此基础上,人们对健康、安全和高品质稻米的需求日益提高。降低稻米中有害物质的积累,并提高其营养成分,成为水稻育种的一个重要新方向。
稻米的安全品质与农田土壤息息相关,由于不当的农艺技术,许多有机和无机污染物进入了农田土壤,其中最为严重的当属重金属污染[3]。农业部对全国30万亩基本农田保护区进行抽样调查,结果显示其中重金属超标率12.1%,产出粮食中重金属含量超标率超过了10%[4]。重金属污染元素包括镉、砷、汞、镍、铅等,其中镉污染的发生率最高,为25%,远超其他土壤重金属污染率[5]。镉是一种有色重金属,与铅锌矿伴生,会随着金属矿石的冶炼而进入到环境中,特别是20世纪的粗放式发展,导致许多农田中出现了镉超标情况[6]。近年来,南方多地先后出现的“镉大米”事件也充分显示了农田镉污染问题的严重性。镉在人体内的代谢半衰期可达到10~35年,会对骨骼、泌尿系统、生殖系统等产生破坏,更甚者会导致畸形和癌症[7-10],因此,长期食用镉超标大米会对身体造成不可逆的严重影响。我国对稻米中的镉含量有明确标准,即大米镉含量要求低于0.2 mg/kg,但国际食品法典委员会对大米中镉的限量为0.4 mg/kg[11-12]。由于农田面源污染治理是一个长期而又艰巨的任务,因此,开发镉低积累型水稻品种具有十分重要的意义。
自然抗性相关巨噬细胞蛋白(natural resistanceassociated macrophage protein, Nramp)是生物体中进化保守的一个大家族,作为质子偶联的金属离子转运体,可以转运锰、锌、铜、铁、镉、镍、钴、和铝离子[13-15]。OsNramp5基因于2012年被初次报道,其在水稻根部负责锰、铁、镉的吸收以及这些元素的转运,亚细胞定位显示OsNramp5蛋白定位于根尖成熟区的内皮层和外皮层的质膜上[16]。敲除该基因可致水稻丧失对镉吸收的大部分能力,同时对锰和铁的吸收也受到一定影响[16-18]。OsNramp5-KO敲除植株的根、地上部和稻米中镉含量都明显低于对照,尤其是稻米中的镉含量下降极为明显[16,19]。为此,有研究认为可用OsNramp5-RNAi植株中籽粒中镉的极低积累而地上部仍有一定量的镉富集特性,作为镉污染农田的土壤修复植物材料[17,20]。在对该基因的相关操作中,发现不同品种的转基因材料农艺表型间不尽相同,如在中花11中敲除该基因可导致幼苗长势变弱,最终产量仅有原材料的11%,在越光中敲除该基因则不影响长势、产量和口感[16,19]。Wang等[21]敲除了黄华占中OsNramp5基因,发现对产量和稻米品质的影响会随着基因缺失程度的增加而增加。Tang等[22]在华占和隆科638S背景中对该基因进行了敲除,得到了不影响生长量、产量和籽粒品质的低镉水稻,同时将它们与隆两优华占进行杂交,得到了性状优良的杂交低镉水稻,说明该基因在低镉水稻育种中具有重要价值。
除了重金属污染导致的稻米品质安全问题外,全球还有约1/3人口存在“隐性饥饿”问题,即由于饮食结构等因素的影响,造成这部分人群对钙、铁、锌、硒等人体四大健康元素的摄入严重不足,最终不利于人体健康。比较而言,钙、铁、锌、硒的缺乏在中国更为普遍[23-24]。比起钙和铁在奶制品及常见的蔬菜中广泛存在,锌和硒的主要食物来源显得不够普遍。中国营养学会建议锌和硒摄入量标准为每人每天15 mg和50 μg[25]。锌元素主要存在于海产品和动物内脏之中,这也决定了我国多数人的日常膳食中对锌的摄入不足。大米中锌的含量很低,精米中锌平均含量仅12 mg/kg,通过大米摄入的锌仅为3.6~6.0 mg/d[26]。对硒元素,Gao等[27]对我国苏州地区居民的每日膳食硒摄入量进行了调查分析,发现当地人均硒摄入量为43.9 μg/d,明显低于WHO和FAO的人均推荐摄入量[28];膳食中的日常硒主要摄入来源是猪肉和谷物,分别占24.7%和22.6%[27]。我国存在一条东北-西南走向的低硒带,纬度跨越大,缺硒地区覆盖了国土面积的72%,暗示我国大多数人对硒的摄入不足[29]。由于稻米是我国绝大多数人的日常主食,增强稻米中的锌和硒含量将是提升我国居民对这两种元素摄入量的可行途径之一。施用锌肥和硒肥或者选择天然富硒土壤是目前富锌、富硒稻米生产的主要策略[30-31]。其中,有研究还发现施用这两种元素的肥料有利于减少水稻对镉的吸收,降低籽粒中镉的含量[32-36]。
南粳46是江苏省近年主要推广的优良食味高产水稻品种,在江苏省内外不同级别的优良食味稻米品比中多次获得金奖,已成为江苏优良食味水稻的“标杆性”品种[37-38]。本研究通过CRISPR/Cas9技术,在南粳46背景下对OsNramp5基因进行敲除,创建了无外源转基因成分的nramp5-KO敲除系,进而对敲除系的主要农艺、产量、品质、抗逆性以及在镉、锌、硒等元素不同处理下籽粒中相关元素积累量进行了全面分析。结果可为健康、安全的高端大米生产提供新材料与种植参考。
1 材料与方法
1.1 试验材料与肥料
供试水稻品种为南粳46及其OsNramp5敲除系nramp5-KO。试验使用的硒肥由苏州硒谷科技有限公司提供,产品编号SETEK-BF-003,其成分为氨基酸≥100 g/L、Cu+Fe+Zn+Mn+B≥20 g/L,Se≥1000 mg/L;叶面喷施的锰肥为MnSO4,锌肥为ZnSO4·7H2O。
1.2 载体构建
利用Miao等[39]描述的CRISPR载体(引自南京农业大学万建民教授团队)和方法构建OsNramp5的基因编辑载体,具体为:使用CRISPR-P网站(http://cbi.hzau.edu.cn/cgi-bin/CRISPR)设计sgRNA,在该网站target genome里选择Oryza sa tiva(RAPDB),Locus Tag中输入OsNramp5的基因登录号(Os07g0257200),在结果中选择评分较高的靶点。通过比较,最终选用的18 bp基因特异性序列为TTCTTCCTGTACGAGAGC,在该序列前后加上接头,构成OsNramp5-sgRNA-F引物:AGATGATCC GTGGCATTCTTCCTGTACGAGAGCGTTTTAGA GCTATGC;将其反向互补获得OsNramp5-sgRNA-R引物:GCATAGCTCTAAAACGCTCTC GTACAGGAAGAAGCCACGGATCATCT。将引物OsNramp5-sgRNA-F和OsNramp5-sgRNA-R直接进行PCR退火反应(在10 μL体系中各加入1 pmol/L的引物,94℃10 min,0.1℃/s退火至15℃,15℃保持10 min),构建到sgRNA中间载体pOs-sgRNA上,接着将其亚克隆到含Cas9表达组件的目标载体pC1300-Cas9上,测序正确后扩大繁殖提取质粒,转入农杆菌感受态细胞EHA105中。
1.3 敲除系的获得与筛选
采用农杆菌介导的植物遗传转化方法转化水稻南粳46的愈伤组织,使用潮霉素筛选,获得T0代转基因植株,采用CTAB法提取T0代植株叶片DNA。以日本晴OsNramp5基因序列为参考序列,利用NCBI(https://www.ncbi.nlm.nih.gov)在目标PAM区两侧适当位置设计引物,以确保可扩增出敲除位点所在的第9外显子,最终设计出引物OsNramp5-CX-F/OsNramp5-CX-R(GGCATCAGT CAGAGGAATCAAGG/GGCACATACGCTGGAC AGTAC)对南粳46野生型和T0代转基因植株的叶片DNA分别进行扩增测序,将T0代转基因植株的扩增测序结果分别与南粳46野生型的测序结果进行比对,分析目标PAM区是否存在序列变异。保留发生突变并引起氨基酸序列改变的T0代突变株进行后续实验。在T1代群体中,采用Cas9蛋白编码基因特异引物Cas9-F/Cas9-R(GTCGCCTACCA CGAGAAGTA/GTGAGGTCCTGGTGGTGCTC)和潮霉素筛选标记基因特异引物HPT-F/HPT-R(ACG GTGTCGTCCATCACAGTTTCC/TTCCGGAAGTG CTTGACATTGGGGA)进行外源转基因成分的检测,筛选出无外源转基因成分的突变植株。为了便于筛选纯合突变体植株,我们针对各T0突变株中目标基因的突变信息,分别开发了dCAPS(derived Cleaved Amplification Polymorphism Sequence)和/或InDel标记进行不同突变信息的特异检测[40]。对野生型和敲除系基因突变区段进行序列分析,结合Primier Premier 5.0 软件,在碱基变异处双侧设计一对PCR引物;对于InDel变异,获得的一对PCR引物即为相应变异的InDel标记;对于dCAP标记,还将进一步利用网站dCAPS Finder 2.0,在变异位点处的前引物中引入1~2个错配碱基,使得该错配碱基与变异位点处的特定碱基间组建为一个限制性内切酶的识别序列,扩增产物即可用该限制性内切酶进行切割,而不具有该切点的则切割不开,从而实现对特定变异位点的分子标记检测。
1.4 田间试验设计
试验分为盆栽和田间试验两部分,均于扬州大学文汇路校区校内试验田进行。
1.4.1 盆栽试验
盆栽试验于2021年5月3日浸种、5月8日播种,6月11日栽秧于盆钵中。盆栽采用长方形长条盆,每盆装入土壤15 kg。盆栽试验中设计三种外源镉浓度,即C0为对照,C1为每1 kg土添加1.2 mg外源镉(1.2 mg/kg),C2为每1 kg土添加2.0 mg外源镉(2.0 mg/kg)。在栽秧前15 d将配置好的CdCl2溶液加入到土壤中,加水浸润土壤并混匀,7 d后再次搅拌。栽秧后将盆栽移入大田中,正常水肥管理。试验设置3次重复,完全随机排列。其中C0盆栽苗设置更多重复,以便用于其他试验处理。喷施锰肥试验采用10 mmol/L MnSO4对C0盆栽中水稻进行叶片喷施,具体方法为:在分蘖末期的晴朗午后,喷至叶面有水珠滴落,3 d后重复喷施一次。高低温逆境胁迫试验始于8月31日,将进入孕穗期的盆栽苗转入玻璃温室,其中低温处理下的最高温度为25℃,最低温度为20℃;高温处理下的最高温度为38℃,最低温度为28℃(图1),湿度维持80%,处理时间为7 d。在处理期间将抽出的穗上挂上标签进行标记,处理结束后,将盆栽搬回大田原位置,最终收获时调查这些穗的结实率。纹枯病菌接种鉴定采用离体茎秆接种法进行,纹枯病菌菌株为本实验室一直保存使用的YN-7[41]。待各材料生长至孕穗期时,从基部剪取长势一致的健康茎秆,在倒2叶叶鞘基部茎节以下1 cm处剪断,剪去茎秆上叶片后在温室中浸水培养24 h,以消除应激反应。将培养携带纹枯病菌丝的木皮插入至倒2叶鞘内,距离叶枕1 cm;接种后将接种茎秆基部插入浸润了营养液的花泥并将花泥放置于盛有营养液的周转箱中;将周转箱转移至专门的纹枯病菌离体茎秆保湿装置中,并放置在人工气候室中;气候室设置30℃/14 h光照、24℃/10 h黑暗,相对湿度75%~95%;1周后测量病斑扩展情况。
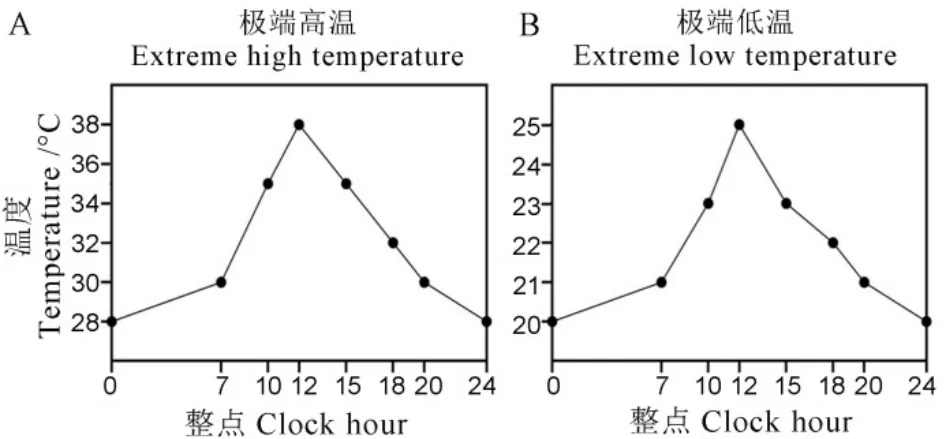
图1 极端温度条件的温度变化曲线Fig. 1. Temperature curves for extreme temperature conditions.
1.4.2 田间试验
田间试验于5月13日浸种、5月17日播种,6月16日栽秧,单株栽秧,行株距为30 cm×11 cm。每个小区8行,每行25株。基肥、返青肥和穗肥分别为300 kg/hm2复合肥(N∶P∶K为26∶10∶12)、112.5 kg/hm2尿素和225 kg/hm2复合肥(N∶P∶K为26∶10∶12),分蘖肥施用尿素,分设3种施用量,即187.5 kg/hm2(高肥)、为112.5 kg/hm2(常规)和37.5 kg/hm2(低肥)。试验采用随机区组试验设计进行。
在穗肥常规用量小区,分别在小区两侧各8行进行锌肥和硒肥喷施试验,其中锌肥使用量为1.5 kg/hm2、质量浓度为1∶1000,硒肥使用量为200 mL/hm2、体积浓度1∶100。于抽穗期的下午5点,采用喷壶进行叶面人工喷施,3 d后重复喷施一次。
1.5 测量内容与方法
1.5.1 农艺性状的测定
按照常规方法测定株高和有效分蘖个数,测定时避开小区边缘,连续测量10株,测定3次。收获上述水稻的所有穗,每株选3个主穗测量穗粒数和实粒数,计算结实率。测量完毕后人工脱粒,晒干(含水量12%左右),用风选机去除空瘪粒,使用自动数粒仪测量千粒重。
1.5.2 元素的测定
样品制备:在成熟期收获稻谷样品,盆栽中为每盆内混取,大田中为小区内五点取样。将收获后的稻谷放入烘箱70℃烘干(含水量12%左右),干燥后脱粒去糠,再使用高速破碎机粉碎,随后过100目筛,装袋备用。
样品消解:取0.5 g样品,装入微波消解仪(Mars)配套的消解罐中,再加入5 mL浓硝酸(GR)、3 mL超纯水和0.2 mL过氧化氢。消解程序为:25 min内升温至180℃,180℃保持10 min,15 min降温至80℃。消解结束后,打开消解罐,放入通风橱中冷却赶酸,结束后过滤到50 mL容量瓶中定容。混匀后取10 mL溶液装入离心管保存,送测ICP-MS(电感耦合等离子体质谱)。
1.5.3 稻米品质的测定
在田间试验的不同处理的各小区中,分别随机选取30个稻株,收获所有稻穗进行人工脱粒。稻谷晒干、风选后,参照国家标准《GB/T17891-2017优质稻谷》测定糙米率、精米率、整精米率、垩白率、垩白粒率、直链淀粉和蛋白质含量等。采用日本佐竹公司的STA1A米饭食味机测定米饭的硬度、黏度、平衡度及综合食味值。
1.6 数据处理
采用Microsoft Excel 2019对数据进行初级整理,采用SPSS 26.0进行相关性状值间的统计分析,其中盆栽试验中的相关性状采用完全随机模型进行统计显著性测定,田间试验中的相关性状采用随机模型进行统计显著性测定。采用GraphPad 8.0.2进行相关图形的绘制。
2 结果与分析
2.1 OsNramp5敲除系创制
通过搜索分析,我们在OsNramp5基因第9外显子区域中的一个PAM位点处设计sgRNA(图2-A),将合成的sgRNA片段与pC1300-Cas9载体进行酶切和连接,最终获得OsNramp5基因敲除载体pC1300-Cas9-OsNramp5-sgRNA(图2-B)。通过农杆菌介导的植物遗传转化方法,最终在南粳46背景中获得了多个T0转基因植株,测序分析显示有5株在目标区域分别存在不同的序列变异,其中3个分别插入了1个不同的碱基(bp),另两个分别缺失了2 bp和21 bp(图2-C)。将敲除变异后的序列均转换成氨基酸序列并与野生型(wild type,WT)序列进行比对,发现这5个敲除系中的OsNramp5蛋白质中的氨基酸均发生突变,表明均已失去原有功能(图2-D)。为了后续能够对这些突变体基因型进行快速鉴定与选择,我们针对其中的3个敲除变异分别成功开发了特异分子标记。对于#1和#2,分别开发了dCAPS标记OsNramp5-G (F:CGGGTGCAGGTTCTTCCTGTACCAG;R:TTGT CGGCGTCCTCTTGG)和OsNramp5-T (F:CGGGT GCAGGTTCTTCCTGTACTAG;R:TTGTCGGCG TCCTCTTGG)。OsNramp5-G标记前引物末端引入了一个错配碱基“C”,利用其扩增#1系时,因为该敲除变异系中存在一个“G”的插入变异而导致PCR产物中具有一个EcoRⅡ(^CCWGG)限制性内切酶的识别位点,相反在WT中PCR产物中由于不存在该插入变异而不能被EcoRⅡ酶切割,因此,在PCR产物经过EcoRⅡ酶切后,纯合突变型的PCR产物可被切掉21 bp,电泳检测后的片段大小明显小于WT(图2-E,泳道1和6),杂合型则呈现双带(图2-E,泳道3、5、7和8)。类似原理设计了OsNramp5-T标记,其前引物末端引入了一个错配碱基“T”,在#2系上该标记的PCR扩增产物可被限制性内切酶SpeI(A^CTAGT)切掉21 bp,使得其片段明显小于WT(图2-E,泳道1、4和8),杂合型则呈现为双带(图2-E,泳道5和6)。对于#5系,我们开发了一个InDel标记OsNramp5-CY(F:TTCGGGCTCCAGTACCAC;R:GAGACGACGGC GATGTTT),在#5系上的扩增片段比WT对照小21 bp(图2-E),因此可以特异鉴别其后代群体中的纯合突变型(图2-E)。
利用3个敲除变异系的特异分子标记及潮霉素和Cas9编码基因特异分子标记,我们在其各单株自交后代群体中连续进行了2个世代的标记辅助选择及验证,同时进行了表型筛选,最终在#1、#2和#5系的各T2群体中分别获得了相应突变型的纯合型敲除系(命名为nramp5ko-1、-2、-3),通过分子标记检测,我们证实这3个株系均不携带Hpt和Cas9蛋白编码基因(图2-F)。
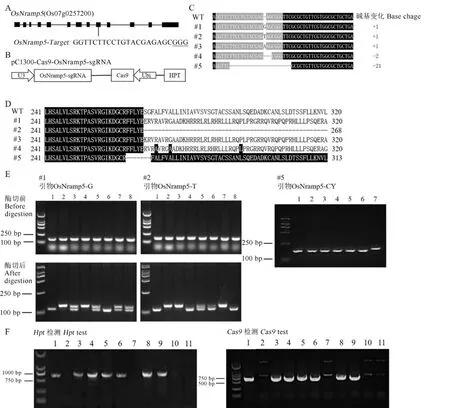
图2 OsNramp5敲除系的构建和基因型检测Fig. 2. Construction and genotyping of OsNramp5 knockout lines.
2.2 OsNra mp5敲除系中镉和锰等元素的积累分析
通过初步观测,我们发现敲除系nramp5ko-3的株高和分蘖力明显低于WT(图3-A、B),生育期也明显延迟,而另外两个系则与WT总体相近,因此在后续试验中,仅对nramp5ko-1和nramp5ko-2两个敲除系开展了相关性状分析。
为了明确敲除系对镉的吸收情况,我们将nramp5ko-1、nramp5ko-2和WT分别种植于镉含量不同的盆栽土壤中。在水稻成熟后,分别收获不同处理下的各材料稻根、地上部和籽粒,测定它们的镉含量。结果显示(图3-C),无论是根部、地上部还是籽粒中,nramp5ko-1和nramp5ko-2中的镉含量均极显著低于WT;当土壤中镉浓度提高时,WT不同部位中的镉含量均迅速上升,而两个敲除系中仅有根部镉含量明显提高,且均明显低于WT。除此之外,无论在C1还是C2镉浓度土壤中,敲除系籽粒中的镉含量均低于0.02 mg/kg,而WT中则均超过国家规定的大米镉含量限制标准0.2 mg/kg。这表明在粳稻中OsNramp5基因同样是负责地上部和籽粒中镉吸收的最关键基因,但在根中似乎还有其他基因参与,敲除该基因可以创制低镉粳稻品种。
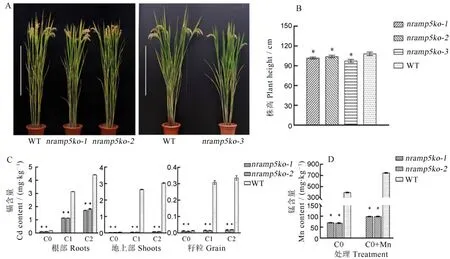
图3 OsNramp5敲除系植株的株型及镉和锰的含量分析Fig. 3. Whole plant and analysis of cadmium and manganese contents in OsNramp5 knockout lines.
由于OsNramp5在水稻中也参与对锰的转运,因此我们测量了敲除系及WT地上部锰的含量,同时分析了叶面喷施锰肥是否有利于补充地上部的锰含量。结果显示(图3-D),敲除系地上部中的锰含量分别为70.08和68.27 mg/kg,极显著低于WT中的392.69 mg/kg;在叶面喷施锰肥后,WT中的锰含量得到了明显提高,达到了743.81 mg/kg,敲除系中的锰含量也得到了显著提高,分别达到98.39 mg/kg和98.56 mg/kg,上升幅度明显低于WT。这些结果表明,OsNramp5确实也是粳稻中对锰的主要吸收和转运基因,但是叶面喷施锰肥可以在一定程度上提高OsNramp5-ko敲除系中的锰含量。
2.3 OsNramp5敲除系的产量性状分析
进一步在大田条件下比较了敲除系nramp5ko-1、nramp5ko-2及WT间的产量相关性状差异,同时设置了三种分蘖肥用量处理。由表1可知,随着分蘖肥用量的增加,水稻的分蘖数和穗粒数均呈上升趋势。有意思的是,在低分蘖肥处理中,WT的分蘖数明显少于敲除系,但在正常和高肥处理下,两个敲除系与WT间的分蘖数均无显著差异。无论在何种分蘖肥用量下,两个敲除系中的穗粒数都少于WT,减少量为25.4~32.8(粒),差异均达到了显著水平。在理论产量上,低分蘖肥用量处理中的两个敲除系与WT间无显著差异,但在正常和高浓度分蘖肥施用处理中,两个敲除系的理论产量均显著低于WT,且主要与穗粒数下降有关。以上结果显示,敲除OsNramp5基因会影响到NJ46的穗粒数并最终造成减产,但在低分蘖肥处理中,未发现明显影响。
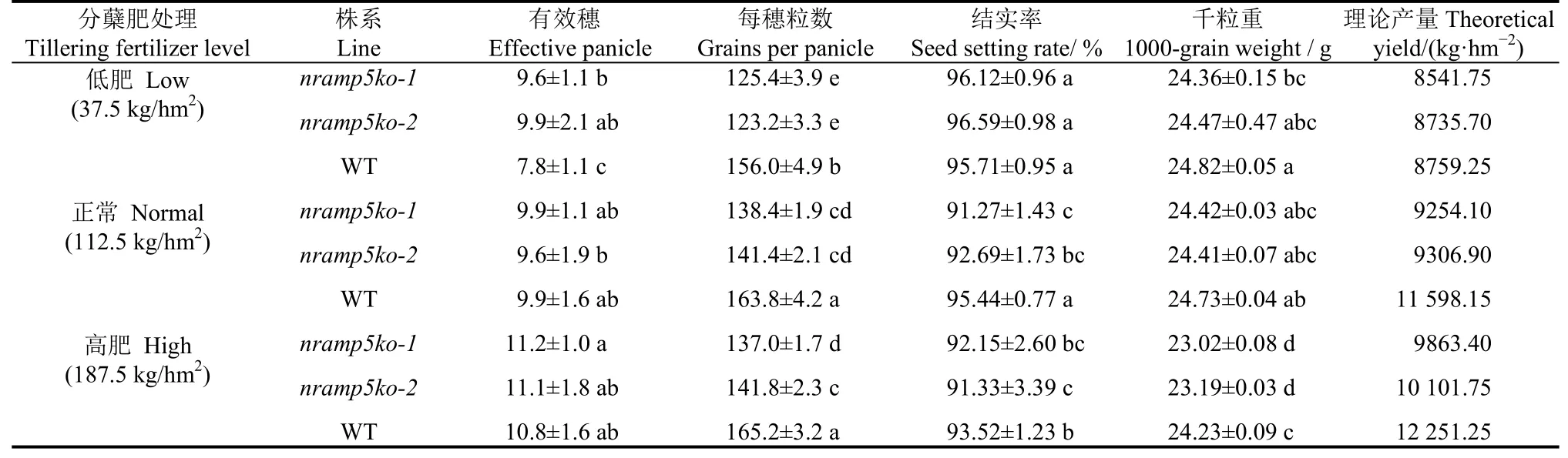
表1 OsNramp5敲除系与对照在不同分蘖肥处理间的理论产量及其构成因素比较Table 1. Comparison of theoretical yield and its components between OsNramp5 knockout lines and control at different tillering fertilizer levels.
2.4 OsNramp5基因敲除系对硒、锌等元素的积累分析
为了评估敲除系是否影响籽粒对人体健康关键营养元素锌和硒的吸收,我们分别在田间正常分蘖肥用量处理中,增设了锌肥和硒肥叶面喷施实验。结果显示(图4),在未喷施两种肥料的WT处理中,敲除系籽粒中的铜、锰和硒的含量均显著低于对照,其中锰含量只有对照一半左右,这与上述盆栽结果一致;镉的含量虽有降低但差异不显著,可能与试验田土壤中的镉含量较低有关;在铁和锌的含量上,敲除系与WT间均无明显差异。在锌肥喷施处理中,敲除系与WT籽粒中的锌含量均显著上升,虽然nramp5ko-1中的含量略高于nramp5ko-2和WT,但是三个材料的总体提升水平相当,敲除系籽粒中的锌平均含量从11.75 mg/kg提高至18.47 mg/kg,WT从11.68 mg/kg提高至18.22 mg/kg;敲除系中的锰和铁含量还是显著低于WT,其他元素含量间均无显著差异。在硒肥处理中,发现敲除系和WT籽粒中硒含量均明显高于未喷施对照,但是敲除系与WT间还是存在极显著差异,敲除系籽粒硒平均含量从对照处理中的0.032 mg/kg升高至0.08 mg/kg,WT从0.046 mg/kg提高至0.12 mg/kg;在其他元素含量上,除了锰之外,敲除系和WT间均无显著差异,如铜含量在未施锌硒肥处理的敲除系与WT间存在显著差异,但在施硒处理中则无差异,这可能与硒肥中含有一定量的铜元素有关。值得注意的是,我们发现施用锌肥和硒肥后,无论敲除系还是WT其籽粒中的镉含量均有所下降,其中施用硒肥下降的更为明显,分别比对照下降了38.3%和46.8%。
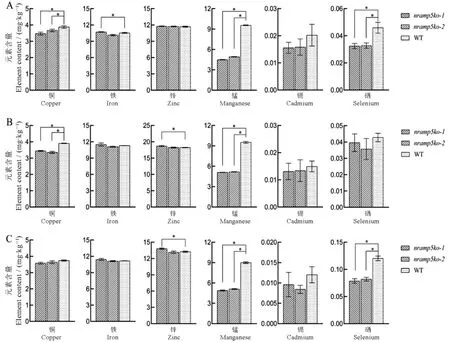
图4 三种施肥处理下OsNramp5敲除系和野生型籽粒中的元素含量Fig. 4. Element contents in grains of OsNramp5 knockout lines and wild type under three fertilization treatments.
2.5 OsNramp5基因敲除系稻米品质分析
为了进一步分析敲除OsNramp5基因是否影响稻米品质,我们分别收集并测定了正常分蘖肥处理及在此基础上增施锌肥和硒肥处理中的相关材料的稻米品质(表2)。就加工品质而言,不同处理内,两个敲除系与WT之间在糙米率和精米率上均无显著差异;整精米率上,施用硒肥和锌肥处理中,两个敲除系的整精米率均显著低于WT,分别降低了2.2%和2.18%。外观品质上,除了硒肥处理的垩白度在敲除系与WT间无显著差异外,其他处理中敲除系垩白粒率和垩白度均显著低于WT,说明敲除系的稻米外观品质总体优于WT。另外,值得注意的是,锌肥处理的所有材料垩白粒率和垩白度均显著高于对照,而施用硒肥则恰恰相反,说明施硒可以改善稻米外观,而施锌则有损外观(图5)。
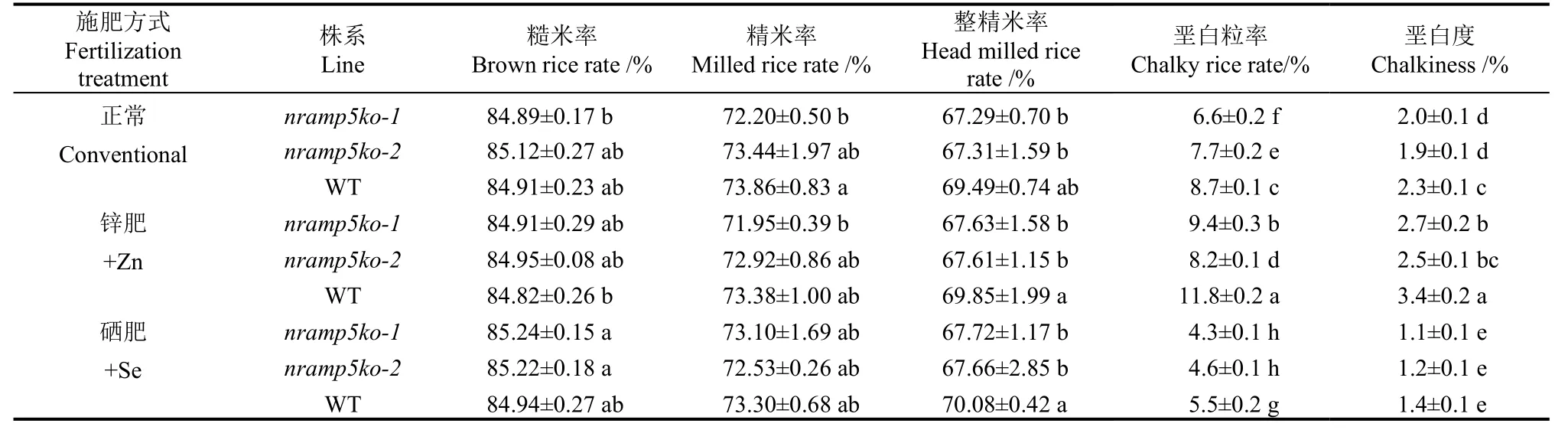
表2 OsNramp5-ko敲除系在不同施肥处理中的稻米加工和品质相关指标差异比较Table 2. Comparison of rice processing and quality related indexes of OsNramp5-ko knockout lines under different fertilization treatments.

图5 不同肥料处理下OsNramp5-KO及对照的精米外观Fig. 5. Milled rice appearance of OsNramp5-KO lines and control under different fertilizer treatments.
进一步对以上材料的营养品质和食味品质进行了测定(表3)。结果显示,在正常施肥处理中,两个敲除系的蛋白质含量和硬度均显著高于WT,而直链淀粉含量、黏度、平衡度和食味质均显著低于WT,其他指标无明显差异。在硒肥处理中,发现与正常施肥对照相比,WT的蛋白质含量得到了显著提高,从6.82%提高至7.25%,但敲除系中无明显影响;各材料的胶稠度均得到了显著降低,黏度尤其对于WT材料也明显降低;WT的食味值显著降低,但是敲除系变化不大。在锌肥处理中,发现正常施肥对照相比,施用锌肥中的各材料直链淀粉含量均显著提高,但在黏度值和食味值上仅有WT明显降低,其他性状均无明显变化。

表3 OsNramp5-ko敲除系在不同施肥处理中的稻米营养品质和食味品质相关指标差异比较Table 3. Comparison of OsNramp5-ko knockout lines in rice nutritional quality and food quality indexes under different fertilization treatments.
2.6 OsNramp5基因敲除系的耐逆性分析
水稻减数分裂期是对温度最敏感的时期,过高或过低的温度均会显著影响花粉育性并最终造成减产。纹枯病一直是威胁水稻安全生产的重要病害之一,为此,我们评价了敲除OsNramp5基因是否影响水稻对高低温和纹枯病的抗性。在高温处理下,我们发现各材料的结实率均显著降低,分别降至45.84%、52.09%和50.16%,彼此间无显著差异;
低温处理下,各材料间的结实率也均受到了显著影响,分别降至79.89%、78.16%和78.94%,敲除系与WT间也无显著差异(图6-A)。在纹枯病接种鉴定中,我们发现敲除系nramp5ko-1和nramp5ko-2的病斑长度分别为10.6 cm和11.5 cm,显著长于WT(7.2 cm)(图6-B)。同时测量了茎秆长度,进而计算获得相对病斑长度,敲除系的相对病斑长度也显著长于WT,结果表明,敲除OsNramp5基因对水稻在高低温环境下的结实率影响不显著,但显著降低水稻对纹枯病的抗性。

图6 OsNramp5敲除系在极端温度及纹枯病菌接种处理下与野生型的抗逆性差异Fig 6. Stress resistance difference between OsNramp5 knockout lines and wild-type under extreme high and low temperature and rice sheath blight inoculation.
3 讨论
3.1 OsNramp5基因在粳稻对镉、锰、铜和硒的吸收和/或转运中具有重要作用
前人研究表明,OsNramp5在水稻中主要负责锰、铁和镉的吸收,敲除该基因会显著影响水稻对三种元素的吸收,尤其是镉和锰[18]。本研究通过CRISPR-Cas9技术在高产优良食味粳稻品种南粳46背景下对该基因进行了敲除,通过测序及标记辅助选项,获得2个不同点突变纯合敲除系。表型鉴定发现敲除该基因可极显著降低粳稻米中的镉含量,尤其在土壤镉含量超标情况下,可确保籽粒中镉积累量远低于国家规定的优质大米中镉含量限值。另外,敲除该基因也确实显著影响了水稻对锰、铜和硒的吸收或转运,但是未发现铁和锌元素的积累受到显著影响。因此,在粳稻对镉和锰的吸收和/或转运上,OsNramp5仍然是最主要的基因,同时也参与对铜和硒的吸收或转运。为了防止锰的积累量下降会影响到水稻的生长发育,我们开展了叶面喷施锰肥试验,发现喷施锰肥可以显著增加敲除系中锰的含量。考虑到水稻对锰的依赖性总体较低,在正常生长条件下,对锰的吸收适当减少基本不会影响水稻的正常生长发育[42]。本研究发现敲除系中的锰含量仅为WT的一半左右,但在整个生长周期,敲除系未出现异常表现或缺锰的表型症状。当然,如果在生产中发现OsNramp5敲除系中存在缺锰现象,那么也是可以通过叶面喷施锰肥进行补充。
3.2 OsNramp5基因敲除系的产量、食味品质和纹枯病抗性下降明显
为进一步评估该基因敲除材料在生产应用中是否存在其他负面效应,我们分别在大田条件下设置多种处理,详细比较了敲除系与野生型在一些重要性状上是否存在差异。总体而言,我们发现敲除OsNramp5基因会导致株高和穗粒数下降,并最终造成单株产量下降,两个系分别比野生型对照减产21.2%和32.0%。在龙起樟和董家瑜等的研究中也发现敲除OsNramp5基因会导致水稻减产[42-43]。在Wang等[21]的研究中,发现了水稻产量等性状与OsNramp5基因突变的幅度有关,越强烈的突变导致了越严重的减产,这可能也是本研究中nramp5ko-3的产量和综合性状明显变劣的可能原因,后续有待进一步的研究。
对OsNramp5基因敲除系的品质性状研究较少,龙起樟等[43]发现敲除系的外观品质和蛋白质含量有所提高而直链淀粉含量略有降低,食味值变化不大。本研究中,我们发现敲除该基因对稻米加工品质基本无影响,但有利于改进外观品质,如降低垩白度和垩白粒率,我们分析这可能与穗粒数变少后更有利于灌浆和籽粒充实有关。敲除系的食味品质明显降低,相关参数指标中显示敲除系的蛋白质含量和硬度显著增加,而黏度和平衡度显著降低,这应该是敲除系食味值降低的最主要原因。此外,我们发现在不同温度处理下的敲除系与WT间的结实率无差异,但敲除系的纹枯病抗性受到了显著影响。这表明,虽然敲除OsNramp5有利于获得低镉育种材料,但也影响了产量、食味品质和抗病性,说明OsNramp5也参与了这些性状的发育调控,但是具体机制是什么,还有待进一步研究。生产中如果利用OsNramp5基因敲除系则需要注意纹枯病的防治。
3.3 叶面喷施锌肥和硒肥会显著影响稻米品质
考虑到锌和硒元素是人类四大生命健康元素,但在多数人日常饮食中的摄入不足,尤其是硒,造成隐性健康问题[44]。为了判断利用OsNramp5基因敲除系在降低镉的吸收基础上,是否影响稻米对锌和硒的吸收,我们对相关材料开展了体外喷施两种肥料试验。虽然发现敲除OsNramp5基因会影响籽粒中硒的含量,但是体外喷施硒肥同样可以显著增加籽粒中的硒含量,达到富硒稻米标准。敲除该基因完全不影响籽粒对锌的富集,且在施锌后可以进一步增加籽粒中的锌含量。以上这些数据结果表明,敲除OsNramp5基因在培育低镉、富硒、富锌粳稻品种中具有重要育种价值。随着基因编辑技术的快速发展,为了更好发挥该基因在培育低镉、健康、优质、高产粳稻新品种中的价值,下一步有必要通过基因编辑技术对该基因进行饱和变异,探索是否存在可以兼顾各种性状平衡的优异等位变异。
在增施锌肥和硒肥中,我们还发现施用锌肥后会导致WT的外观品质和食味值显著下降,施用硒肥虽然可以提升外观品质但同时也影响胶稠度和食味值,造成食味值明显下降。我们怀疑这可能与在抽穗杨花扬花期期施用这些元素后会激活或影响籽粒中的淀粉和蛋白合成或结构变异的基因表达有关,具体有待进一步分析研究。
猜你喜欢
粮油食品科技(2020年6期)2020-12-08
红楼梦学刊(2020年2期)2020-02-06
现代装饰(2018年4期)2018-05-22
耕作与栽培(2016年6期)2017-01-17
北方水稻(2016年2期)2016-12-02
西南农业学报(2016年4期)2016-05-17
西南农业学报(2016年6期)2016-04-16
现代农业(2016年5期)2016-02-28
乡村科技(2016年30期)2016-02-19
—— 品尝员的识别能力与适口性
北方水稻(2016年4期)2016-01-09
