
广西鸭坦布苏病毒流行株GXQZ01-DTMUV-2020全基因组序列分子特征分析
2023-01-17 09:01谢守玉熊陈勇施开创韦显凯冯淑萍屈素洁陆文俊骆永泉钟华训侯慧贤覃婉婷韦慧琦尹彦文
中国兽医学报 2022年12期
谢守玉,熊陈勇,施开创,李 军,郑 敏,韦显凯,冯淑萍,屈素洁,龙 凤,杨 蓉,陆文俊,骆永泉,钟华训,侯慧贤,覃婉婷,韦慧琦,尹彦文*
(1.广西动物疫病预防控制中心,广西 南宁 530001;2.广西大学 动物科学技术学院,广西 南宁 530005;3.贺州市水产畜牧站,广西 贺州 542699)
坦布苏病毒(Tembusu virus,TMUV)是属于黄病毒科黄病毒属恩塔亚病毒群不分节段的单股正链RNA病毒。1955年,首次从马来西亚的三带喙库蚊体内分离到TMUV。目前,已从鸭、鸡、鹅、麻雀及蚊子体内发现该病毒[1]。2010年,上海地区的麻鸭出现生长迟缓、高热、食欲减退、产蛋下降及死亡症状。随后,我国养鸭主产区的东南省份,如浙江、江苏、福建、山东、湖南等地相继有疫情发生,发病率高达90%,病死率为5%~30%,致病原曾被称为“鸭产蛋下降综合征病毒”,后经证实为鸭坦布苏病毒(duck Tembusu virus,DTMUV)[2-4]。马来西亚和泰国的蛋鸭养殖场陆续有DTMUV引发疫情的报道[5-6]。
DTMUV基因组全长约11 kb,包含5′端和3′端非编码区(UTR),以及1个开放阅读框(ORF)。ORF编码C、prM和E 等3种结构蛋白,以及NS1、NS2A、NS2B、NS3、NS4A、NS4B和NS5等7种非结构蛋白。E蛋白是DTMUV主要膜蛋白和结构蛋白,为宿主的重要保护性抗原,能够诱导动物机体发生特异性免疫应答,产生中和抗体[7]。NS1蛋白是胞外分泌性蛋白和多效应蛋白,也是在病毒复制过程中产生的重要抗原,可诱导机体产生细胞免疫反应[8]。E蛋白和NS1蛋白均具有良好的免疫原性,是DTMUV亚单位疫苗研发的重点对象。为了解广西DTMUV流行毒株基因组分子特征,本试验利用已建立的DTMUV检测方法[9],从诊断为DTMUV阳性的病料中扩增全基因组序列,与国内外参考毒株进行相似性比较,对ORF、E及NS1基因进行遗传进化分析,发现广西DTMUV流行毒株呈现遗传多样性的特征,以期为广西DTMUV流行情况调查提供数据支持,为DTMUV有效防控和致病机理研究奠定基础。
1 材料与方法
1.1 主要试剂核酸抽提试剂盒(批号:20120110T015)购于天隆科技公司;PrimeScriptTMOne Step RT-PCR Kit Ver.2(批号:ASF0454A)、MiniBEST Agarose Gel DNA Extraction Kit Ver 4.0(批号:AK11054A)、pMD18-T载体(批号:AK91481A)、大肠杆菌DH5α感受态细胞(批号:AK71028A)均购于宝生物(大连)公司。
1.2 病料检测病料来自2020年广西钦州某养鸭场发病的麻鸭。采集患病麻鸭肝脏、脾脏、肾脏、肺脏等组织样品,剪成小块后放入2 mL灭菌离心管中,加入适量PBS,经磨碎仪研磨至糜状,离心后取上清提取核酸,作为模板,利用本实验室已建立的DTMUV检测方法进行检测[9]。
1.3 病毒全基因组扩增测序与比对以DTMUV检测结果为阳性的核酸为模板,利用本试验设计的DTMUV全基因组测序引物(表1)进行RT-PCR扩增。建立50 μL反应体系:模板2 μL,PrimeScript One Step Enzyme Mix 2 μL,2×One Step Buffer 25 μL,上、下游引物各1 μL(20 μmol/L),无RNA酶灭菌水补足体系。反应程序:50℃ 30 min;94℃ 2 min;94℃ 30 s,50℃ 30 s,72℃ 2 min,35个循环。反应结束后,PCR产物通过1.0%琼脂糖凝胶电泳鉴定。

表1 全基因组测序引物
PCR产物胶回收纯化后,连接至pMD18-T载体,转化大肠杆菌DH5α感受态细胞,筛选阳性克隆测序,每个片段重复测序3次。使用DNAStar软件包中的SeqMan拼接测序获得的片段,最终得到GXQZ01-DTMUV-2020株全基因组序列,并将GXQZ01-DTMUV-2020株全基因组序列同参考毒株进行比对。
1.4 DTMUV全基因组序列分析应用BioEdit软件对GXQZ01-DTMUV-2020株及参考毒株(表2)全序列进行核苷酸和氨基酸相似性分析。应用MEGA7软件对比对后的ORF、E及NS1基因序列进行最佳核苷酸替换模型评估:ORF为GTR+G+I;E、NS1基因均为TN93+G。以最佳核苷酸替换模型为基础,采用最大似然法(maximum likelihood)绘制系统发育树,Bootstrap值设置为1 000次。应用RDP 4(recombination detection program 4)和SimPlot(ver 3.5.1)软件对GXQZ01-DTMUV-2020株及参考毒株的全基因组序列进行重组分析,检测是否有重组现象。应用BEAST(ver 1.10.4)软件包估算ORF、E及NS1基因的遗传进化速率,评估进化的快慢。

表2 DTMUV主要参考毒株信息
2 结果
2.1 全基因组测序与比对结果应用SeqMan程序拼接测序的片段,得到1株DTMUV,命名为GXQZ01-DTMUV-2020株。结果显示,GXQZ01-DTMUV-2020株全长为10 991 bp,5′端UTR为94 bp,3′端UTR为619 bp,含有1个开放阅读框(ORF)为10 278 bp,编码3 425个氨基酸。其中,3种结构蛋白基因C、prM和E长度分别为360,501,1 503 bp;7种非结构蛋白基因NS1、NS2A、NS2B、NS3、NS4A、NS4B及NS5长度分别为1 056,681,393,1 857,378,762,2 715 bp。MM_1775和GXQZ01-DTMUV-2020株的E、NS1蛋白氨基酸序列比对结果显示,E、NS1蛋白分别有14个和21个氨基酸位点发生变异(表3)。3′端UTR核苷酸序列比对发现,DTMUV-AH2011株在第82位至155位连续插入74个碱基。SD14株在第43位至49位,第71位至73位不连续插入共10个碱基。在第60个核苷酸位置,包括GXQZ01-DTMUV-2020株在内共有15株DTMUV插入1个G碱基。

表3 MM_1775和GXQZ01-DTMUV-2020株的E、NS1蛋白氨基酸序列对比
2.2 核苷酸与氨基酸相似性分析结果应用BioEdit软件对GXQZ01-DTMUV-2020株与参考毒株全序列相似性分析结果显示,其与广西DTMUV流行株的核苷酸相似性为94.1%~98.5%,氨基酸相似性为95.8%~100.0%;与水禽源TMUV参考毒株的核苷酸相似性为85.7%~99.7%,氨基酸相似性为92.0%~100.0%;与蚊虫源TMUV参考毒株的核苷酸相似性为85.8%~99.1%,氨基酸相似性为92.0%~100.0%。E、NS1基因与广西DTMUV流行株的核苷酸相似性为95.9%~96.4%和96.1%~97.6%,氨基酸相似性为99.0%~99.4%和98.2%~99.1%;与水禽源TMUV参考毒株的核苷酸相似性为89.3%~99.2%和87.4%~99.4%,氨基酸相似性为98.0%~99.8%和94.0%~100.0%;与蚊虫源TMUV参考毒株的核苷酸及氨基酸相似性为88.1%~98.0%和85.8%~99.0%,96.8%~99.4%和93.4%~99.7%;与鸡源TMUV(Sitiawan virus)核苷酸及氨基酸相似性为86.8%/96.8%,85.2%/92.6%(表4)。结果表明,GXQZ01-DTMUV-2020株与广西DTMUV毒株相似性较高,与水禽源TMUV相似性高于其他宿主源TMUV。

表4 GXQZ01-DTMUV-2020株与部分参考毒株同源性分析 %
2.3 遗传进化树绘制应用MEGA7对ORF、E及NS1基因绘制遗传进化树(图1,2)。ORF遗传进化树中(图1A),MM_1775株与 Sitiawan virus组成TMUV群,SD14株为3群,DK/TH/CU-DTMUV2007株为1群。GXQZ01-DTMUV-2020株与广西GX2012,GX2013E株属于2.2亚群,广西GX2011,GX2015株与国内主要DTMUV毒株组成2.1亚群。结果表明,广西DTMUV流行毒株进化方向不完全相同,呈现遗传多样性特征。从图1A 中可以得出,GXQZ01-DTMUV-2020株与参考毒株CHN-YC和DTMUV/Goose/CHN/2019/AQ-19遗传关系距离较近,这与相似性分析结果一致。E基因(图1B)和NS1(图2)基因遗传进化树分布与ORF相似。
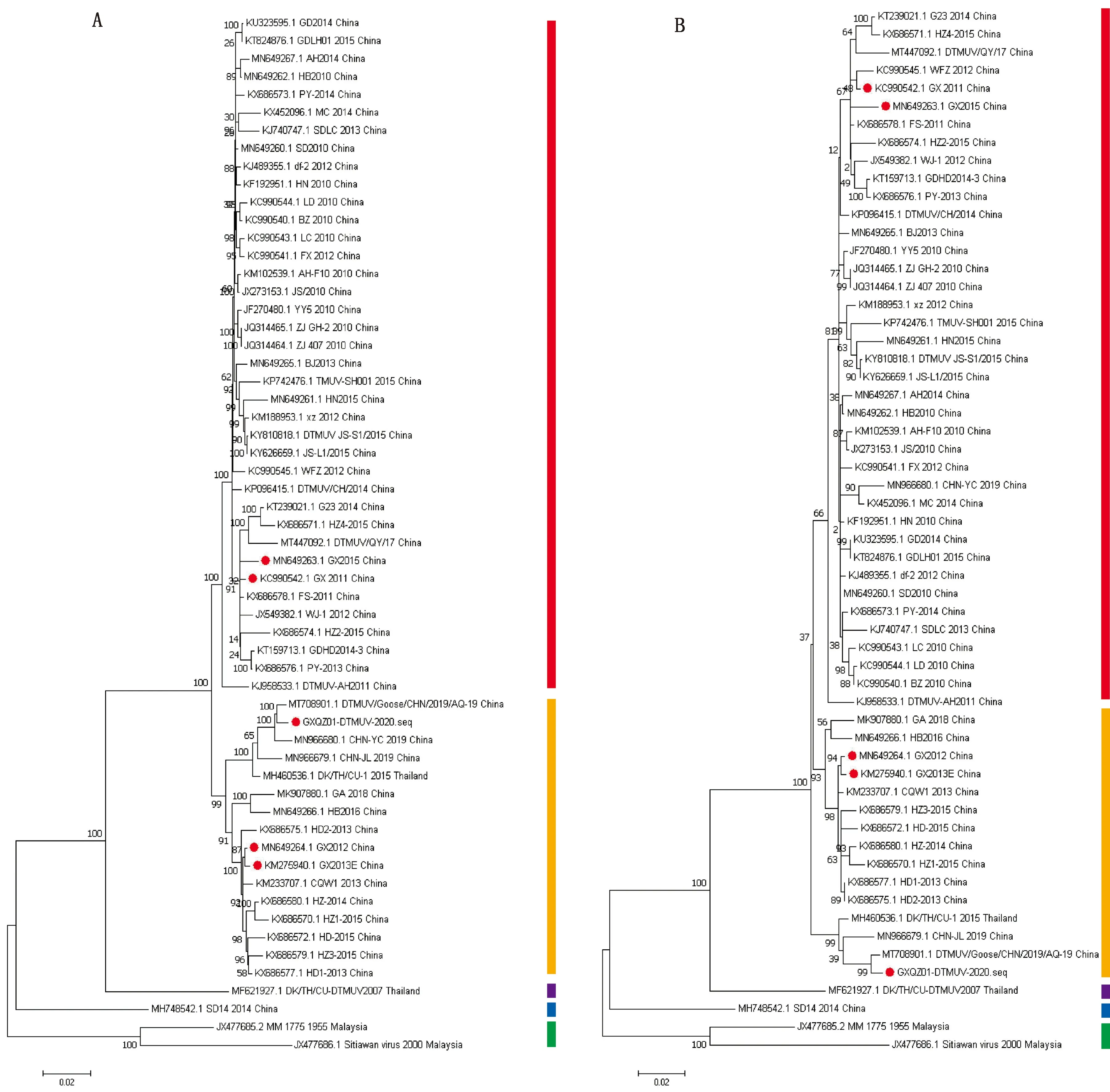
A.ORF;B.E基因。红色.2.2亚群;橙色.2.1亚群;紫色.1群;蓝色.3群;绿色.TMUV。下同
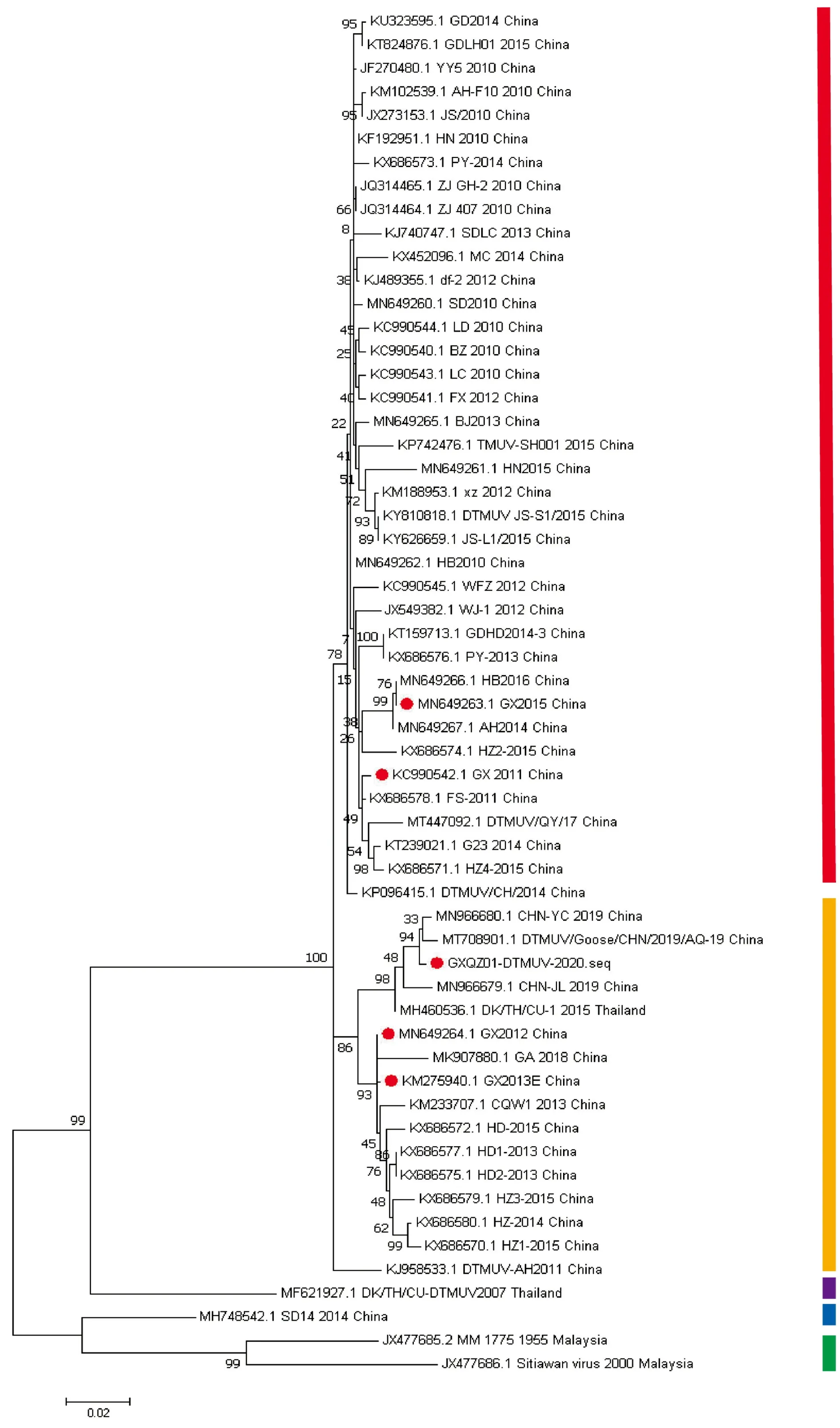
图2 DTMUV NS1基因遗传进化树
2.4 重组分析结果应用RDP4和SimPlot软件对GXQZ01-DTMUV-2020株与参考毒株的全基因组序列进行重组分析。结果显示,HB2016、HD2-2013、CHN-YC及AH2014株检测到重组信号(图3)。HB2016株重组的主要亲本为GA,相似性为98.9%,次要亲本为GX2015株,相似性为100%,重组位点为比对后的第2 049~4 054位;HD2-2013株重组的主要亲本为HD-2015株,相似性为99.3%,次要亲本为GDHD2014-3株,相似性为99.9%,重组位点为第5 675~7 237位;CHN-YC株重组的主要亲本为DTMUV/Goose/CHN/2019/AQ-19株,相似性为99.2%,次要亲本为MC株,相似性为100%,重组位点为第870~1 989位;AH2014株重组的主要亲本为HB2010株,相似性为99.9%,次要亲本为GX2015株,相似性为99.9%,重组位点为第2 044~4 002位。
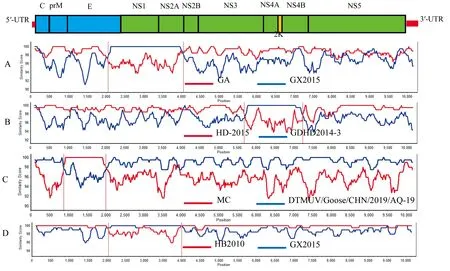
A.HB2016;B.HD2-2013;C.CHN-YC;D.AH2014
2.5 遗传进化速率估算结果应用BEAST软件对GXQZ01-DTMUV-2020株及参考毒株的ORF、E及NS1基因遗传进化速率估算结果为1.146×10-3,1.467×10-3,1.319×10-3替换/(位点·年),表明E基因进化速率最快。
3 讨论
广西壮族自治区地处沿海亚热带,与东南亚国家接壤,属于候鸟迁徙过冬地区,而TMUV宿主谱广泛,存在野鸟传播病毒的潜在风险。据报道,养鸭场从业人员的血清TMUV阳性检出率高达71.9%,说明TMUV宿主范围扩大,对公共卫生安全可能造成威胁[10]。因此,须加强对TMUV的流行情况调查及分子流行病学研究。本试验从广西钦州某蛋鸭养殖场采集病死鸭的脾脏、肝脏、肾脏、肺脏及卵巢组织,利用本实验室已建立的DTMUV病原诊断方法进行检测[9],以阳性核酸为模板,进行DTMUV全基因组序列扩增,得到GXQZ01-DTMUV-2020全序列。通过序列比对发现,E蛋白氨基酸共有14个位点发生变异,其中第154~157位NYPV变异为NYSA。TMUV E蛋白第156位氨基酸的改变将影响病毒的组织嗜性和在动物群体间的传播能力,以及第154位氨基酸的糖基化修饰作用[11-13]。NS1蛋白有2个糖基化位点,通过借助E蛋白疏水信号进入内质网,在参与病毒侵袭、组装、释放及致病性方面都起到关键作用[14-15]。本研究中NS1有21个氨基酸位点发生变异,尚需深入探讨突变的意义。在3′端UTR区域,DTMUV-AH2011株连续插入74个碱基,SD14株不连续插入10个碱基,并且在系统发育树上为独立的分支(数据未列出),尚未见其他文献有相关报道,至于突变对毒株的影响需要进一步研究。
相似性分析结果显示,GXQZ01-DTMUV-2020株基因组结构中,5′UTR、3′UTR、NS1、NS2A及NS4B与CHN-YC株核苷酸相似性最高;prM、E、NS2B、NS3及NS5与DTMUV/Goose/CHN/2019/AQ-19株核苷酸相似性最高。其中,C和NS4A基因核苷酸相似性与CHN-YC和DTMUV/Goose/CHN/2019/AQ-19株并列最高,为99.4%和98.9%,氨基酸相似性均为100%。以上结果表明,GXQZ01-DTMUV-2020基因组呈现分子遗传多样性特征。GXQZ01-DTMUV-2020与国内TMUV参考毒株的核苷酸相似性为85.7%~99.7%,氨基酸相似性为92.0%~100%。E、NS1基因与国内TMUV参考毒株的核苷酸相似性为89.3%~99.2%和87.4%~99.4%,氨基酸相似性为95.4%~99.8%和94.0%~100%。任丹等[16]分离到1株鹅源TMUV全基因组序列与国内DTMUV相似性最高达97.7%,与鹅源TMUV相似性为96.7% 左右。胡峰等[17]分离的2株水禽源TMUV,与参考毒株的相似性大于95.7%,E基因与参考毒株的核苷酸相似性为95.2%以上。李刚[18]分离的山东DTMUV毒株与2017年分离株的ORF氨基酸相似性为98%以上;与2010―2012年分离株的氨基酸相似性为96.8%~98.6%。农海连[19]从广西分离的4株DTMUV与参考毒株的核苷酸相似性高于96%,氨基酸相似性高于98%。刘烈发等[20]从江西省某蛋鸭养殖场分离到1株DTMUV,与已报道的DTMUV参考毒株核苷酸相似性在98.5%以上。李文俊等[21]从鸭组织病料中分离出1株DTMUV,其E基因与DTMUV参考毒株相似性最高达99.2%,与鸡源TMUV毒株相似性为86.7%。以上表明,国内DTMUV毒株之间的相似性普遍较高,与本研究的分析结果相似,说明国内DTMUV毒株遗传变异较小。
水禽源、鸡源及蚊虫源TMUV的ORF遗传进化树可分为4个进化群,GXQZ01-DTMUV-2020株与GX2012、GX2013E株等组成2.1亚群;GX2011、GX2015株与国内主要DTMUV组成2.2亚群,这是国内DTMUV优势群体;马来西亚毒株组成TMUV群;山东SD14株为3群;泰国DK/TH/CU-DTMUV2007株为1群。广西DTMUV有3株位于2.1亚群,2株位于2.2亚群,从毒株分离年份上尚未发现相关性。GXQZ01-DTMUV-2020株与DTMUV/Goose/CHN/2019/AQ-19和CHN-YC处于同一进化分支上,遗传关系距离最近,这与相似性分析结果一致。GX2012、GX2013E株和重庆CQW1株处于相邻进化分支上,GX2011、GX2015株与广东FS-2011株处于同一进化分支。并且,GXQZ01-DTMUV-2020株与广西其他4株DTMUV遗传关系较远,呈现不同的进化趋势。E基因的遗传进化树中,GXQZ01-DTMUV-2020株与2.2亚群的优势毒株遗传距离比GX2012、GX2013E更远,这与ORF和NS1不同。NS1遗传进化趋势与ORF相似。PENG等[22]对国内外TMUV毒株进行遗传进化分析,其中广西毒株的分群结果与本研究一致。NINVILAI等[23]对2015―2017年分离的泰国DTMUV毒株进行遗传特征分析,GX2013E处于2.1亚群,绝大多数中国DTMUV毒株位于2.2亚群。农连海[19]分离的4株广西DTMUV毒株,遗传进化分支上与CQW1遗传关系最近,这与本研究结果一致。综上,广西DTMUV毒株呈现不同的遗传进化趋势,表现遗传多样性的特征。
DTMUV全基因组序列重组分析发现,共有4株存在重组现象。其中,HB2016和AH2014重组的位点相似,位于E、NS1及NS2A基因,CHN-YC重组的位点位于E基因,HD2-2013重组的位点位于NS3、NS4A和NS4B基因。E蛋白作为宿主的重要保护性抗原,在病毒复制过程中,对病毒的吸附、融合及受体结合等发挥着至关重要的作用[7,24]。非结构蛋白在病毒核酸的复制、蛋白的合成加工与病毒粒子组装等方面起关键性作用[25]。因此,这些基因发生重组,须引起密切关注。DAI等[26]利用RDP 4软件检测到一株DTMUV发生重组,重组位点位于prM和E基因。本研究对ORF、E及NS1基因遗传进化速率估算的结果为1.146×10-3,1.467×10-3,1.319×10-3替换/(位点·年)。NINVILAI等[23, 27]对亚洲DTMUV毒株的ORF、E基因遗传进化速率估算结果为1.113×10-3,1.507×10-3替换/(位点·年)。YU等[28]对DTMUV的E基因进化速率评估结果为5×10-4替换/(位点·年)。DAI等[27]对TMUV全基因组序列遗传进化速率估算结果为5.9×10-4替换/(位点·年)。
综上,同一个基因或片段的进化速率估算结果有差异,可能与选择的参考毒株种类、数量及软件参数设置等因素有关。因此,须进一步加强DTMUV流行情况调查及基因组分子结构分析,为DTMUV的有效防控提供数据支持。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
河北画报(2020年8期)2020-10-27
中国饲料(2019年19期)2019-03-25
中国猪业(2017年11期)2017-12-11
浙江大学学报(工学版)(2016年2期)2016-06-05
