
干湿交替灌溉耦合施氮量对水稻籽粒灌浆生理和根系生理的影响
2023-01-16 05:05:28杨文博王越涛李本银王付华王生轩尹海庆
作物学报 2023年3期
付 景 王 亚 杨文博 王越涛 李本银 王付华 王生轩 白 涛 尹海庆,*
干湿交替灌溉耦合施氮量对水稻籽粒灌浆生理和根系生理的影响
付 景1王 亚1杨文博1王越涛1李本银2王付华1王生轩1白 涛1尹海庆1,*
1河南省农业科学院粮食作物研究所, 河南郑州 450002;2河南省农业科学院植物营养与资源环境研究所, 河南郑州 450002
干湿交替灌溉耦合施氮量对水稻根系生长和产量形成有重要影响, 但其对籽粒灌浆生理的影响, 以及与根系生理的关系尚不明确。为探讨干湿交替灌溉和施氮量对水稻籽粒灌浆、籽粒淀粉合成相关酶活性和激素含量变化及其根系生理的影响, 以超级稻品种南粳9108为材料, 大田种植, 设置常规灌溉(conventional irrigation,CI)和干湿交替灌溉(alternate wetting and drying, AWD) 2种灌溉方式及5个施氮水平, 全生育期不施氮肥(0N)、全生育期施氮肥90 kg hm–2(90N)、全生育期施氮肥180 kg hm–2(180N)、全生育期施氮肥270 kg hm–2(270N)和全生育期施氮肥360 kg hm–2(360N)。结果表明: 灌溉方式与施氮量存在显著的互作效应, 干湿交替灌溉增加了南粳9108籽粒最大灌浆速率和平均灌浆速率, 提高了籽粒中蔗糖合成酶、腺苷二磷酸葡萄糖焦磷酸化酶、淀粉合成酶、淀粉分枝酶的活性和玉米素+玉米素核苷、3-吲哚乙酸、脱落酸的含量, 增加了花后根系氧化力和根系中玉米素+玉米素核苷含量, 促进水稻生育前期茎鞘中储存的NSC向籽粒的运转, 且与270N耦合后产量最高, 为本试验最佳水氮耦合模式。表明通过适宜的水肥调控发挥水氮耦合效应, 可以提高水稻根系生理性能和籽粒灌浆生理活性, 实现水稻高产。
水稻; 干湿交替灌溉; 施氮量; 籽粒灌浆生理; 根系生理
水稻是我国主要粮食作物之一, 稳定提高水稻产量对于保障我国粮食安全具有重要的战略意义。目前以高投入为手段的栽培管理方式已成为提升我国水稻生产能力的重要途径[1-2]。过高的氮肥投入和不合理的灌溉方式给生态环境带来巨大的压力, 水氮资源利用效率低下是我国水稻生产的一个主要问题[2-3]。如何提高水氮资源利用效率已成为当前研究的热点。水分和氮素是作物生长发育过程中重要的影响因子, 水氮互作对改善作物生长环境, 特别是根系生长, 促进地上与地下部协调, 进而提高产量有重要意义。
我国水稻年种植面积达3000万公顷, 也是灌溉用水量最大的农作物, 约有70%的农业用水用于水稻灌溉, 随着人口增长、城镇和工业发展、全球气候变化以及环境污染加重, 用于作物灌溉的水资源愈来愈匮乏, 严重威胁作物特别是水稻生产的发展, 为应对水资源短缺问题, 农业科学家开发了一些水分高效利用新技术[4-5], 其中干湿交替灌溉是水稻种植行之有效的节水灌溉技术[6]。但是有关干湿交替灌溉对水稻产量的影响还存在争议[7], 有待于进行深入研究。除了水资源以外, 氮素是另一个决定作物产量的关键因素。水分和氮素对水稻生长发育的影响在数量和时间上存在着最佳的耦合效应。在水分亏缺条件下, 氮素可以一定程度上缓解干旱胁迫的压力, 而水又是肥效发挥的关键因素。水分和氮素既互为促进, 也互为制约[8]。只要水分和氮肥供应合理匹配, 就会产生相互促进机制, 实现作物产量、水分和氮肥利用效率的协同提高。以往有关水、氮的研究多集中在单因子管理上, 对于双因子效应的研究较少, 且存在争议[9-10]。水氮互作的研究多集中在对水稻产量、根系形态生理结构、叶片光合和水氮利用效率等方面[8,11-13], 而对水稻籽粒灌浆特性及其生理的研究尚未见报道, 本研究拟通过控制土壤落干程度, 观察节水灌溉和施氮量耦合对水稻籽粒灌浆的调控作用及其生理基础, 以期为实现水稻高产和水分、氮素高效利用提供理论及科学依据。
1 材料与方法
1.1 试验材料和栽培概况
于2017—2018年在河南省现代农业开发基地(新乡市平原新区, 35°00′N, 113°40′E)进行。供试品种为南粳9108。试验地气候属于暖温带大陆性季风气候, 2017年和2018年水稻生长季平均气温分别是23.2℃和23.5℃, 总降水量分别是295.9 mm和464.8 mm, 年日照时数分别是1148.0 h和1277.7 h。大田前茬作物为小麦, 土壤类型为沙壤土, 耕作层含有机质19.4 g kg–1、有效氮96.7 mg kg–1、速效磷26.2 mg kg–1、速效钾109.7 mg kg–1。2年试验均为5月6日播种, 6月8日移栽, 9月3日至5日抽穗, 10月25日收获。株行距为15 cm × 25 cm, 每穴3株苗栽插。
1.2 试验设计
采用灌溉方式×氮肥水平二因素随机试验。设置2种灌溉处理方式: 1) 常规灌溉(conventional irrigation, CI), 即保持浅水层, 分蘖末期进行轻度搁田与收获前一周断水; 2) 干湿交替灌溉(alternate wetting and drying, AWD), 除移栽至返青田间保持浅水层外, 其余时期采用干湿交替灌溉技术, 即自浅水层自然落干到土壤水势达-15 kPa时, 灌水1~2 cm, 再自然落干至-15 kPa, 再上浅水层, 如此循环。在AWD处理小区安装真空表式土壤负压计(中国科学院南京土壤研究所生产), 每小区安装4个土壤负压计监测15~20 cm深土壤水势。每天12:00记录土壤水势, 当读数达到阈值时, 灌1~2 cm水层。在进水管安装水表(LXSG-50流量计, 上海水分仪表制造厂)用以监测用水量。氮肥设5个水平, 即全生育期不施氮肥(0N, 以N计, 下同)、全生育期施氮90 kg hm–2(90N)、180 kg hm–2(180N)、270 kg hm–2(270N)和360 kg hm–2(360N)。氮肥运筹按照基肥(移栽前1 d)∶分蘖肥(移栽后7 d)∶促花肥(叶龄余数3.5)∶保花肥(叶龄余数1.5)=4∶2∶2∶2施用。基施过磷酸钙(含P2O513.5%) 300 kg hm–2。移栽前1 d和穗分化始期分别施用氯化钾(含K2O 52%) 135 kg hm–2和90 kg hm–2。每个处理均重复3次, 完全随机区组排列, 小区面积为5.4 m×4.5 m。全生育期严格防治病虫草害。
1.3 测定项目与方法
1.3.1 取样与籽粒灌浆速率测定 于抽穗期, 选择各小区生长整齐一致、同日开花的穗子200个挂上纸牌, 并标记开花日期。自开花至花后45 d每隔5 d取各处理标记穗45个(每小区15个), 用于测定籽粒重量和蔗糖-淀粉代谢途径关键酶活性及其激素含量。将约2/3籽粒样品置液氮冷冻1 min后放入-70℃冰箱保存用于内源激素含量及其蔗糖-淀粉代谢途径关键酶活性测定, 另1/3籽粒在70℃下烘干至恒重, 然后剥壳称重, 测定籽粒增重动态并按朱庆森等[14]方法用Richards[15]生长方程对籽粒灌浆过程进行拟合, 计算灌浆速率:
=/(1+e-1/N(1)
对方程(1)求导, 得到籽粒灌浆速率():
=e–kt/[(1+e–kt)(N+1)/N] (2)
方程中,为粒重(mg),为最终粒重(mg),为花后天数。为回归方程所确定的参数, 活跃灌浆期定义为达到最终粒重的5% (1)和95% (2)所经历的时间:
=2(+2)/(3)
这段时间内籽粒(糙米)增加的重量除以灌浆时间(2-1)为平均灌浆速率:
mean=/[2(+2)] (4)
1.3.2 籽粒中有关酶活性和激素含量测定 取35~40粒去壳籽粒加3~5 mL 100 mmol L–1Tricine- NaOH提取液[pH 8.0, 含有10 mmol L–1MgCl2, 2 mmol L–1EDTA, 50 mmol L–12-mercaptoethanol, 12% (v/v) glycerol, 5% (w/v) PVP 40]于研钵中研磨(温度保持在0℃), 15,000×下离心5 min (4℃), 取上清液(粗酶液)用于各酶活性测定。SuSase、AGPase、StSase和SBE的活性测定方法参照Yang等[16]的方法。参照Bollmark等[17]介绍的酶联免疫法提取与测定籽粒中玉米素+玉米素核苷、IAA和ABA含量。
1.3.3 根系氧化力和根系玉米素+玉米素核苷测定
于抽穗期和抽穗后10、20和30 d, 取样测定。为了确保所取样品具有代表性, 取样前连续考查每小区20穴植株的茎蘖数, 计算出平均茎蘖数, 按平均茎蘖数各处理取6穴, 每穴稻株以稻根基部为中心, 用取根器取出15 cm×25 cm×20 cm的土块, 剪去地上植株部分, 将地下部分装于70 mm的筛网袋中先用流水冲洗, 再用农用压缩喷雾器将其冲洗干净, 参照杨建昌等[18]的方法测定根系氧化力; 根系玉米素+玉米素核苷含量测定同籽粒中玉米素+玉米素核苷含量测定。
1.3.4 茎鞘中非结构性碳水化合物的测定 于抽穗期和成熟期考查每个小区20穴植株的茎蘖数, 计算出平均茎蘖数, 按平均茎蘖数各处理取6穴(抽穗期用根系取样时地上部植株), 将植株分解为叶、茎鞘和穗, 烘干后采用全自动样品快速研磨仪粉碎后过0.15 mm的样品筛, 参照杨建昌等[19]方法, 测定茎鞘中非结构性碳水化合物(NSC, 可溶性糖和淀粉之和)的含量。茎鞘中NSC的运转率(%)=(抽穗期茎鞘中NSC含量-成熟期茎鞘中NSC含量)/抽穗期茎鞘中NSC含量×100; 茎鞘中NSC对产量的贡献率(%)=(抽穗期茎鞘中NSC含量-成熟期茎鞘中NSC含量)/稻谷产量×100。
1.3.5 考种与计产 取成熟期各小区50穴考察每穴穗数, 10穴用于观察结实率(水漂法, 沉入水底者为饱粒)和千粒重。各小区去除边行后, 剩余稻株实收计产。
1.4 数据处理
采用Microsoft Excel 2013、SPSS16.0和SAS软件处理数据与统计分析, 用SigmaPlot 10.0绘图。
2 结果与分析
2.1 水稻产量和籽粒灌浆生理及根系生理的处理效应
在2017和2018两年试验中, 水稻产量、籽粒平均灌浆速率、籽粒和根系中细胞分裂素含量和根系氧化力在灌溉方式、施氮水平间存在显著差异(<0.01), 水分×氮肥存在互作效应, 其余的互作效应均不显著(表1)。所测定的主要指标年度间差异均不显著, 说明水分、施氮水平对产量及籽粒灌浆生理和根系生理的影响在年度间有较好的重演性。因此, 文中除产量外, 其余指标主要取2018年的试验数据作进一步分析。
2.2 产量及产量构成因素
由表2可知, 不同水分和氮肥处理对南粳9108产量及产量构成因素的影响不同, 水分和氮肥间存在明显的耦合效应(表2)。在同一灌溉方式下, 在0~360 kg hm–2施氮量范围内, 随施氮量的增加, 南粳9108的每穗粒数和总颖花量均显著增加, 结实率和千粒重逐渐降低, 单位面积穗数和产量在0~270 kg hm–2施氮量范围内, 随施氮量增加均显著增加, 在270~360 kg hm–2施氮量范围内, 单位面积穗数随施氮量增加变化不显著, 而产量却随施氮量增加显著减少。90N、180N、270N、360N处理在CI方式下分别比不施氮处理产量增加21.1%~21.7%、36.6%~ 37.1%、56.6%~57.1%和40.8%~41.6%, 在AWD方式下分别比不施氮处理产量增加18.7%~20.8%、30.5%~32.1%、48.8%~51.9%和36.0%~38.3%。在0~ 270 kg hm–2施氮量范围内, 单位面积穗数和每穗粒数的增加是其产量增加的主要原因, 而在270~360 kg hm–2施氮量范围内, 结实率和千粒重的降低是其产量又下降的主要原因(表2)。
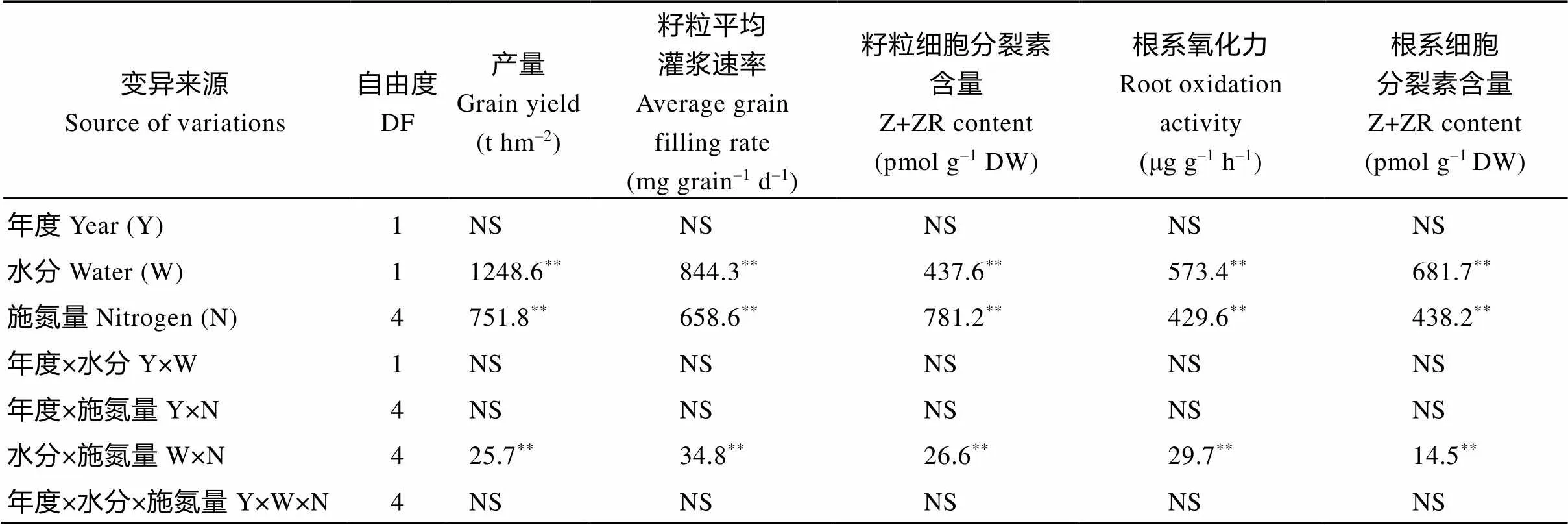
表1 水分和氮肥耦合下水稻产量及灌浆特性的方差分析
NS表示在0.05水平上差异不显著。*与**分别表示在0.05和0.01水平差异显著。
NS: not significant at> 0.05.*and**represent0.05 and0.01, respectively.
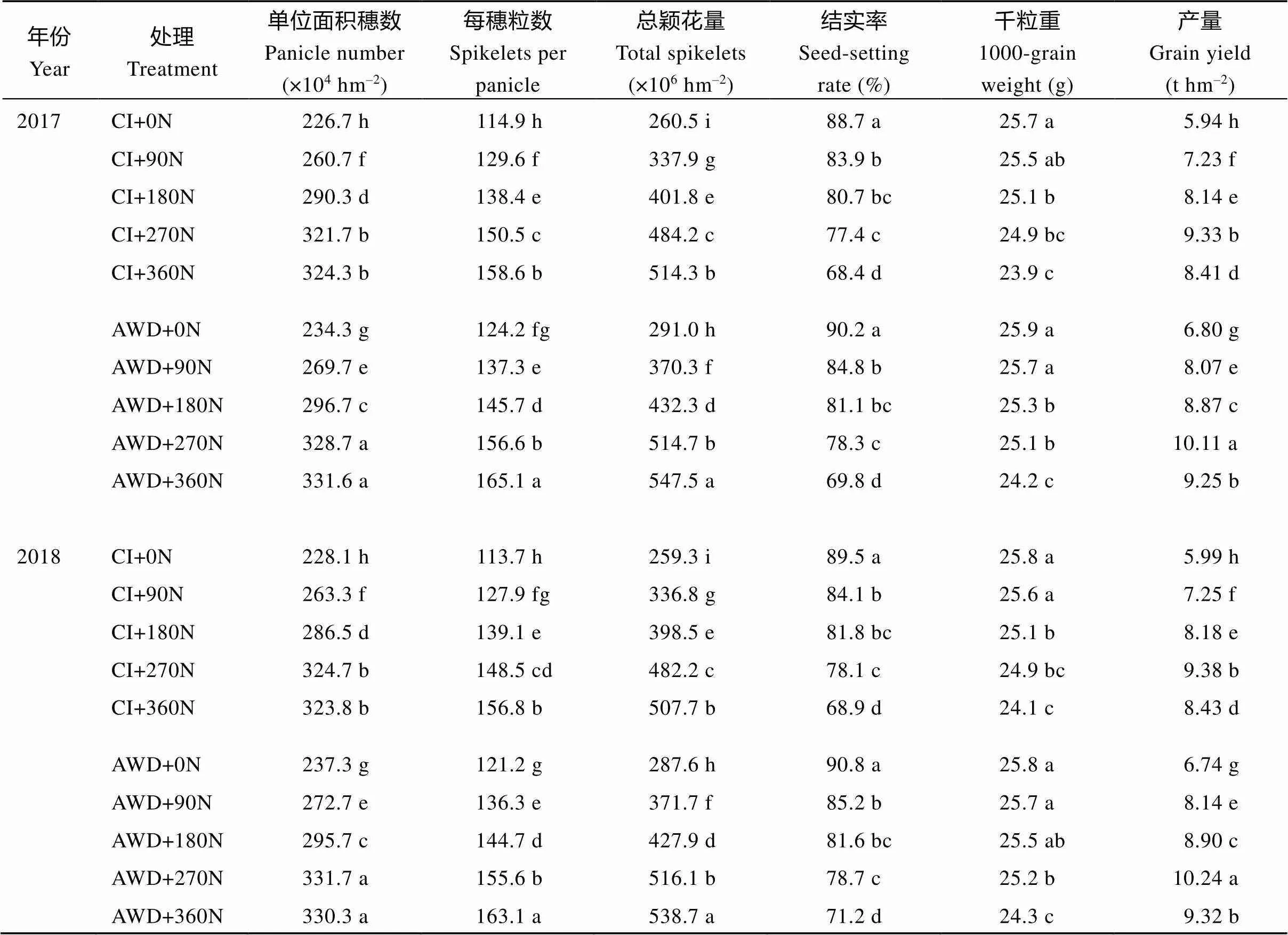
表2 水分和氮肥耦合对水稻产量及产量构成因素的影响
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2; 同一列标以不同字母表示0.05水平差异显著。
CI: conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2; Different lowercase letters within the same column indicate significant difference at the0.05 probability level.
在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉显著提高了南粳9108的产量、单位面积穗数、每穗粒数和总颖花量, 提高了结实率和千粒重, 但差异不显著。0N、90N、180N、270N和360N处理时, AWD比CI方式下产量增加12.5%~14.5%、11.6%~12.3%、8.8%~9.0%、8.4%~9.2%和10.0%~ 10.6%。从水分和氮肥耦合效应分析, 0N和90N在干湿交替灌溉条件下有利于产量的提高。
通过施氮量与产量的曲线方程, 计算出南粳9108在常规灌溉和干湿交替灌溉方式下的最适施氮量分别是286.5~287.4 kg hm–2和292.9~294.2 kg hm–2。表明南粳9108在不同灌溉方式下对氮肥的需求量不同, 适宜施氮量应根据灌溉方式进行调节, 干湿交替灌溉方式下应适当增加施氮量, 这样有利于提高产量, 水分和氮肥耦合效应更加显著(表3)。

表3 不同灌溉方式下水稻获得最高产量时的施氮量
CI: 常规灌溉; AWD: 干湿交替灌溉;opt: 获得最高产量时的施氮量;max: 最高产量。
CI: the conventional irrigation; AWD: the alternate wetting and drying;opt: the optimal nitrogen fertilizer rete for maximum grain yield;max: the maximum grain yield.
2.3 水稻籽粒灌浆
不同水分和氮肥耦合条件下南粳9108的籽粒增重动态以及由Richards生长方程拟合的籽粒灌浆速率曲线见图1。由图1可知, 南粳9108的籽粒重量和灌浆速率在不同的水分和氮肥耦合下存在差异。在同一灌溉方式下, 在0~360 kg hm–2施氮量范围内, 随施氮量的增加, 南粳9108的最大灌浆速率、平均灌浆速率和糙米重减小或者显著减小, 到达最大灌浆速率的时间推迟或显著推迟, 活跃灌浆期延长或显著延长(表4)。其中施氮量在270~360 kg hm–2范围内时变化幅度较大(图1和表4)。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高或显著提高了南粳9108的最大灌浆速率、平均灌浆速率和糙米重, 延迟或显著延迟了到达最大灌浆速率的时间, 缩短或显著缩短了活跃灌浆期(表4)。
2.4 水稻籽粒中蔗糖-淀粉代谢途径关键酶活性变化及其与籽粒灌浆的关系
由图2可知, 籽粒中蔗糖合成酶(SuSase)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合成酶(StSase)和淀粉分枝酶(SBE)活性到达峰值的时间有早有迟, 但均为单一的峰值曲线, 即随着灌浆进程, 籽粒中酶活性增加, 达到峰值后迅速下降。在同一灌溉方式下, 在0~360 kg hm–2施氮量范围内, 随施氮量的增加, 南粳9108的峰值出现推迟且峰值减小,之后籽粒酶活性下降速度减弱。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高或显著提高了南粳9108的籽粒酶活性(图2)。
对水分和氮肥耦合下南粳9108籽粒的蔗糖-淀粉代谢途径关键酶活性与灌浆速率进行相关分析, 结果列于表5。
籽粒中SuSase活性与灌浆速率呈极显著正相关(= 0.806**, 0.916**), AGPase和StSase活性与灌浆速率呈显著或极显著正相关(= 0.780*, 0.896**;= 0.677*, 0.835**), SBE活性与灌浆速率呈正相关(= 0.419, 0.569), 但未达到显著水平。表明SuSase、AGPase和StSase活性与籽粒灌浆速率有密切关系, 籽粒灌浆速率小, 粒重轻与其较低的SuSase、AGPase和StSase活性有关。
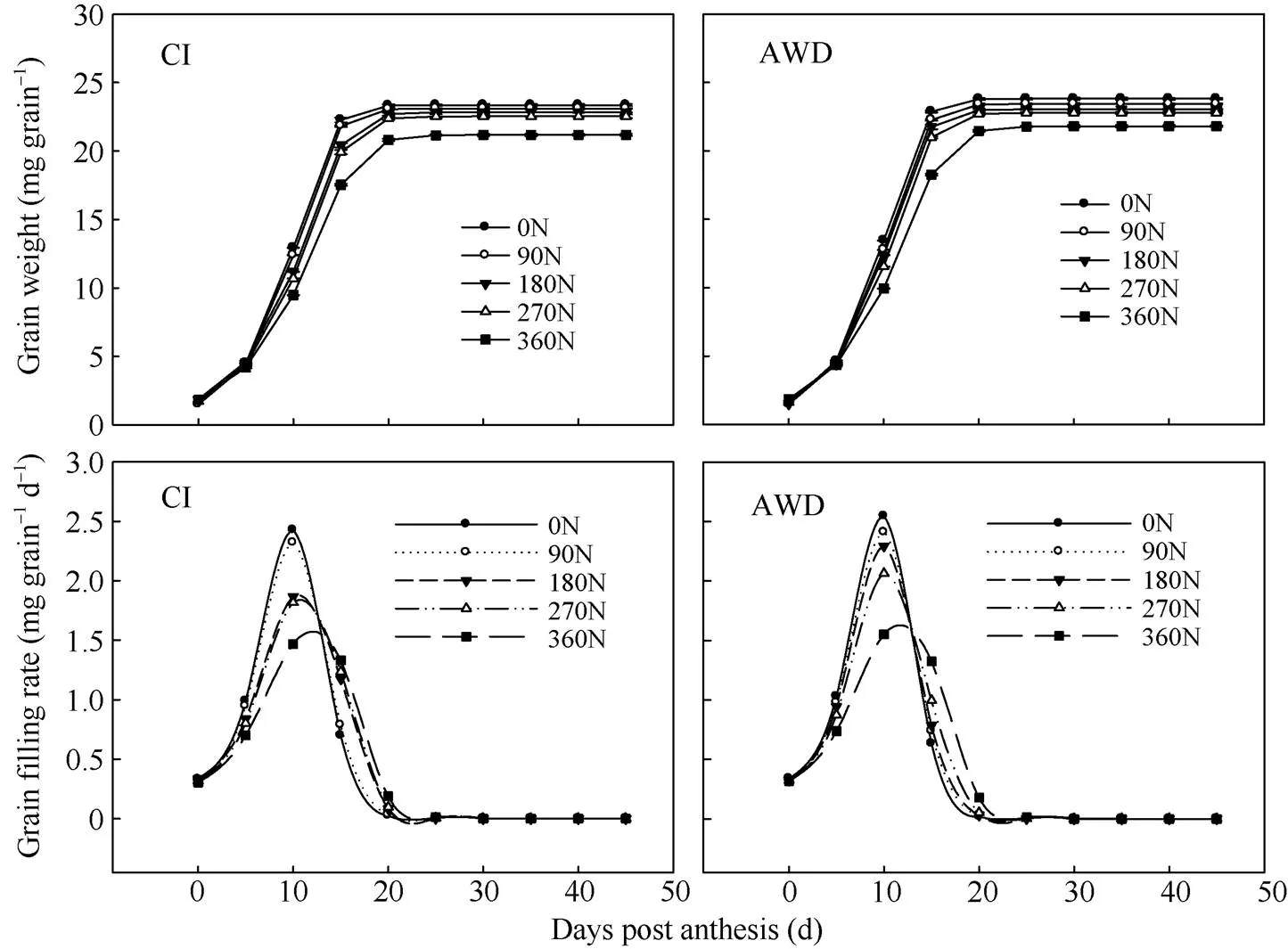
图1 水分和氮肥耦合下水稻籽粒增重动态和籽粒灌浆速率
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。
CI: the conventional irrigation; AWD: alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.
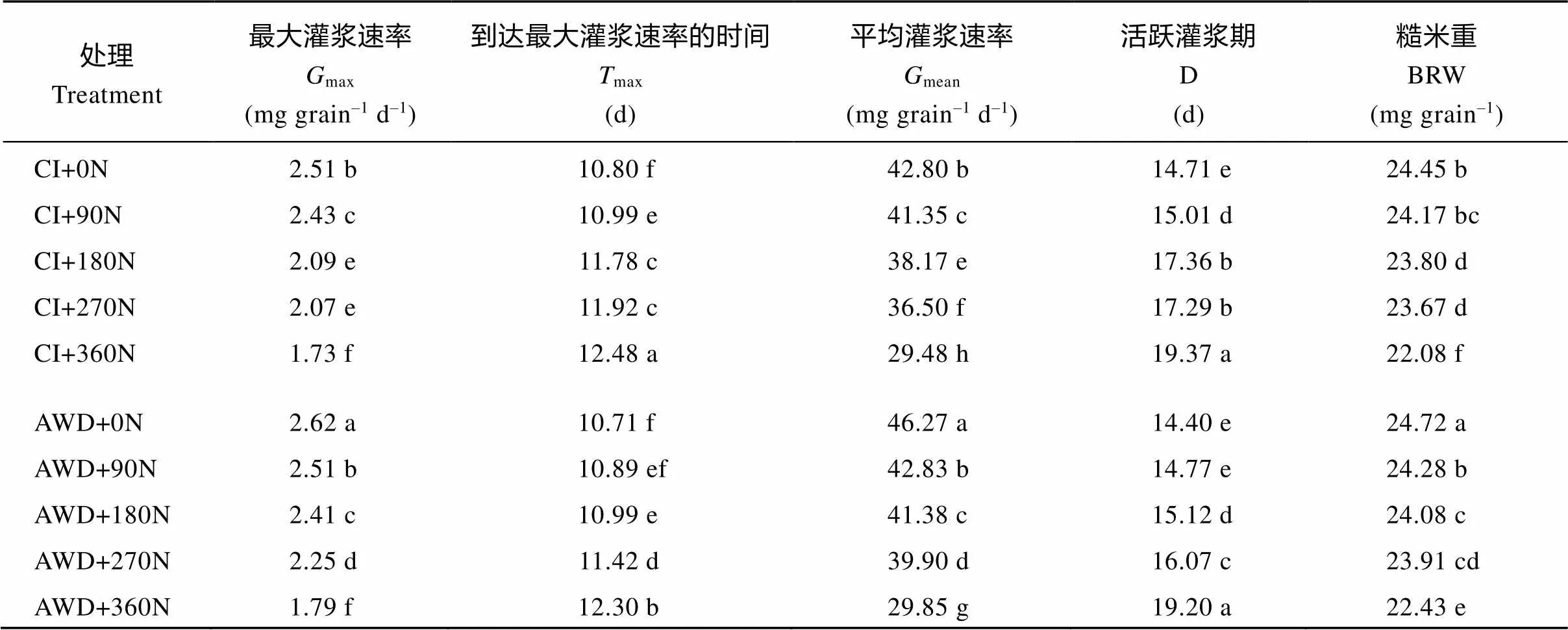
表4 水分和氮肥耦合下水稻籽粒灌浆特征参数
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm-2; 同一列标以不同字母表示=0.05水平差异显著。
CI: conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2; Different lowercase letters within the same column indicate significant difference at the0.05 probability level.
2.5 水稻籽粒中激素含量变化及其与籽粒灌浆的关系
由图3可知, 籽粒中玉米素+玉米素核苷(Z+ZR)、3-吲哚乙酸(IAA)和脱落酸(ABA)含量到达峰值的时间有早有迟, 但均为单一的峰值曲线, 即随着灌浆进程, 籽粒中激素含量增加, 达到峰值后迅速下降。在同一灌溉方式下, 在0~360 kg hm–2施氮量范围内, 随施氮量的增加, 南粳9108的峰值出现推迟且峰值减小, 之后籽粒激素含量下降速度减弱。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高或显著提高了南粳9108的籽粒激素含量(图3)。
对水分和氮肥耦合下南粳9108籽粒中激素含量与灌浆速率进行相关分析, 结果列于表6。籽粒中Z+ZR含量与灌浆速率呈显著或极显著正相关(= 0.728*, 0.875**), IAA含量与灌浆速率呈极显著正相关(= 0.832**, 0.960**), ABA含量与灌浆速率呈正相关(= 0.025, 0.379), 但未达到显著水平。表明Z+ZR和IAA含量与籽粒灌浆速率有密切关系, 籽粒灌浆速率小, 粒重轻与其较低的Z+ZR和IAA含量有关。
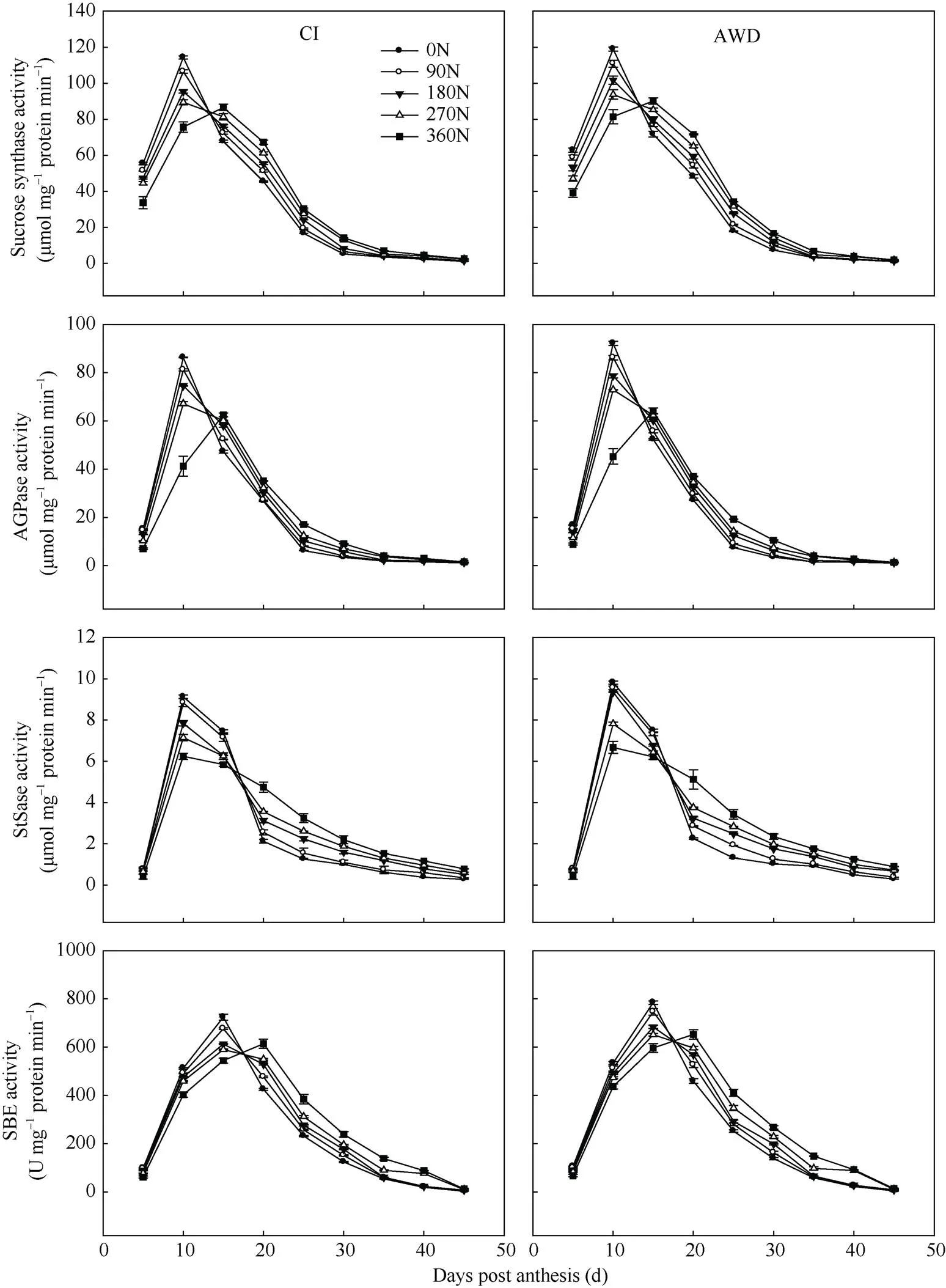
图2 水分和氮肥耦合下水稻籽粒蔗糖-淀粉代谢途径关键酶活性变化
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.
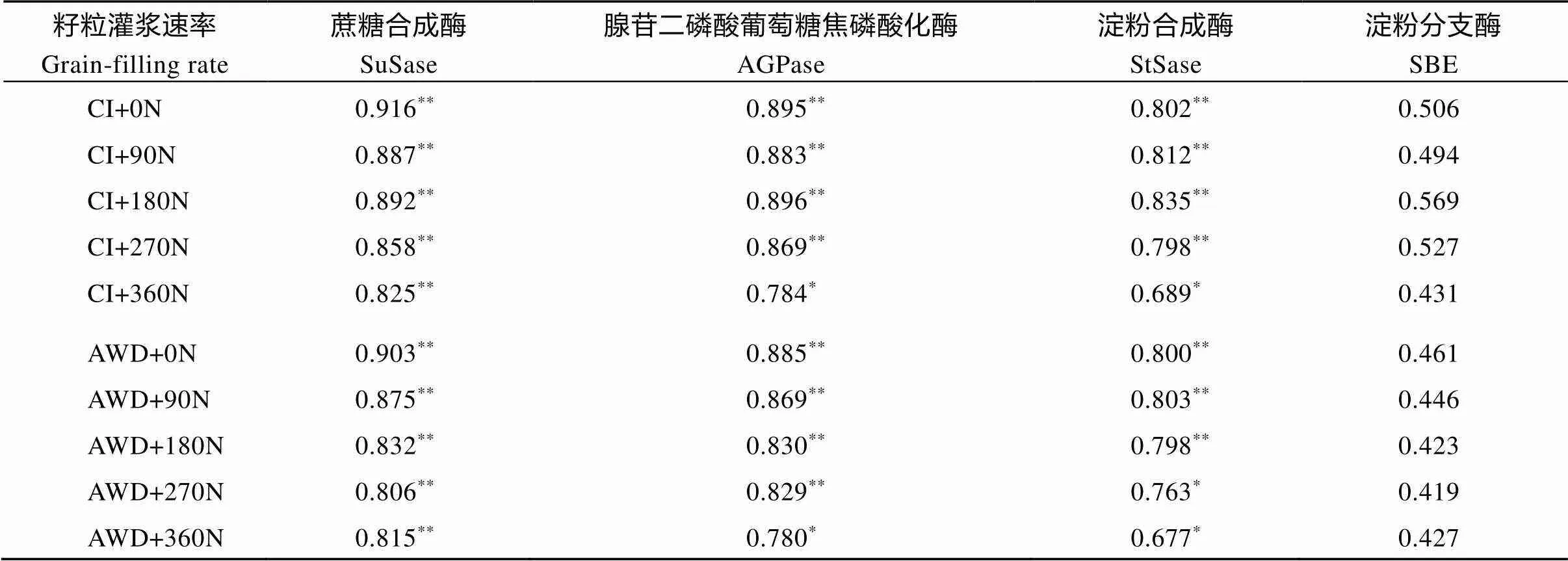
表5 水分和氮肥耦合下水稻籽粒灌浆速率与籽粒中蔗糖-淀粉代谢途径关键酶活性的相关
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。*与**分别表示在0.05和0.01水平差异显著。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.*and**represent0.05 and0.01, respectively.
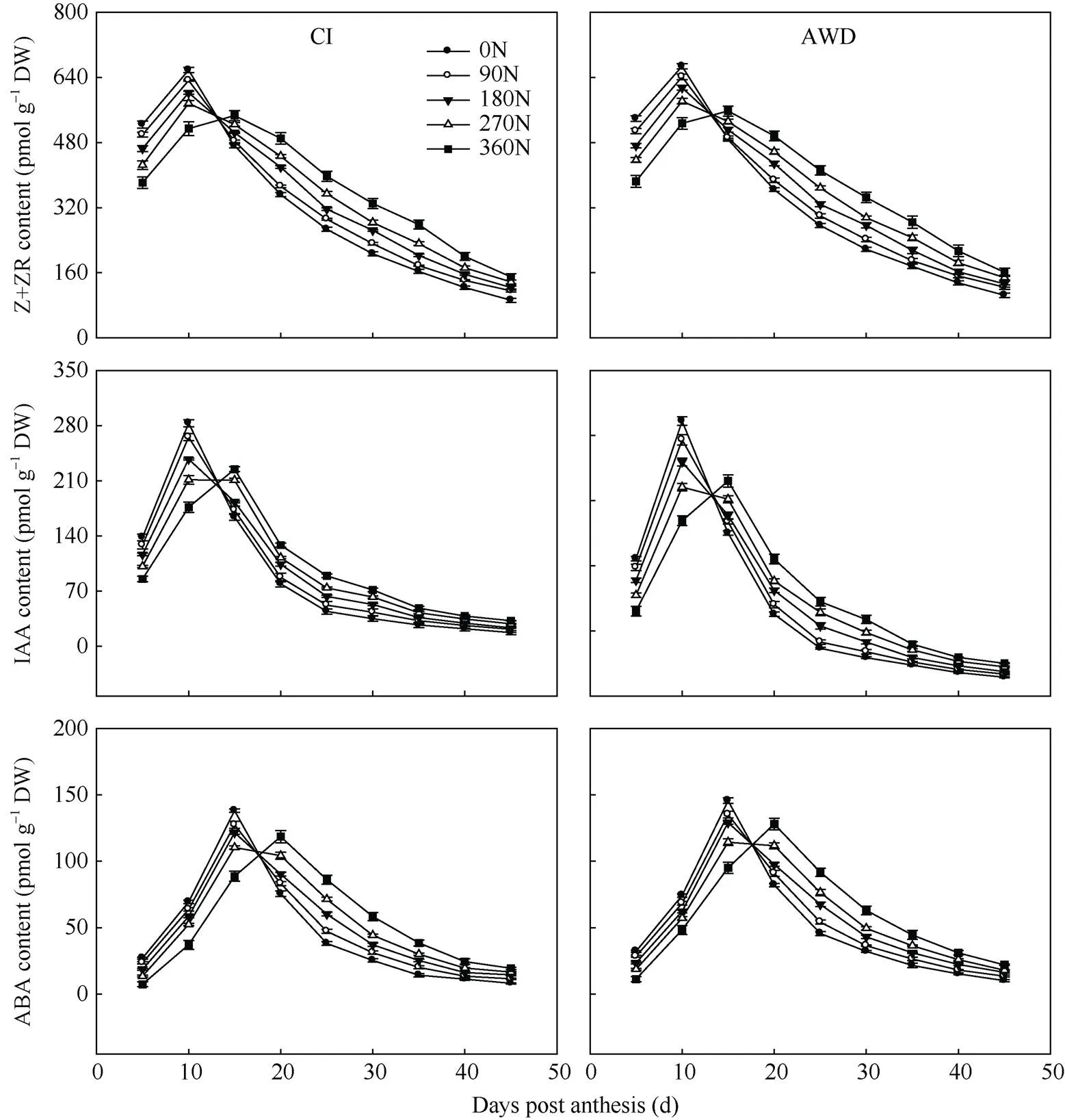
图3 水分和氮肥耦合下水稻籽粒激素含量变化
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.
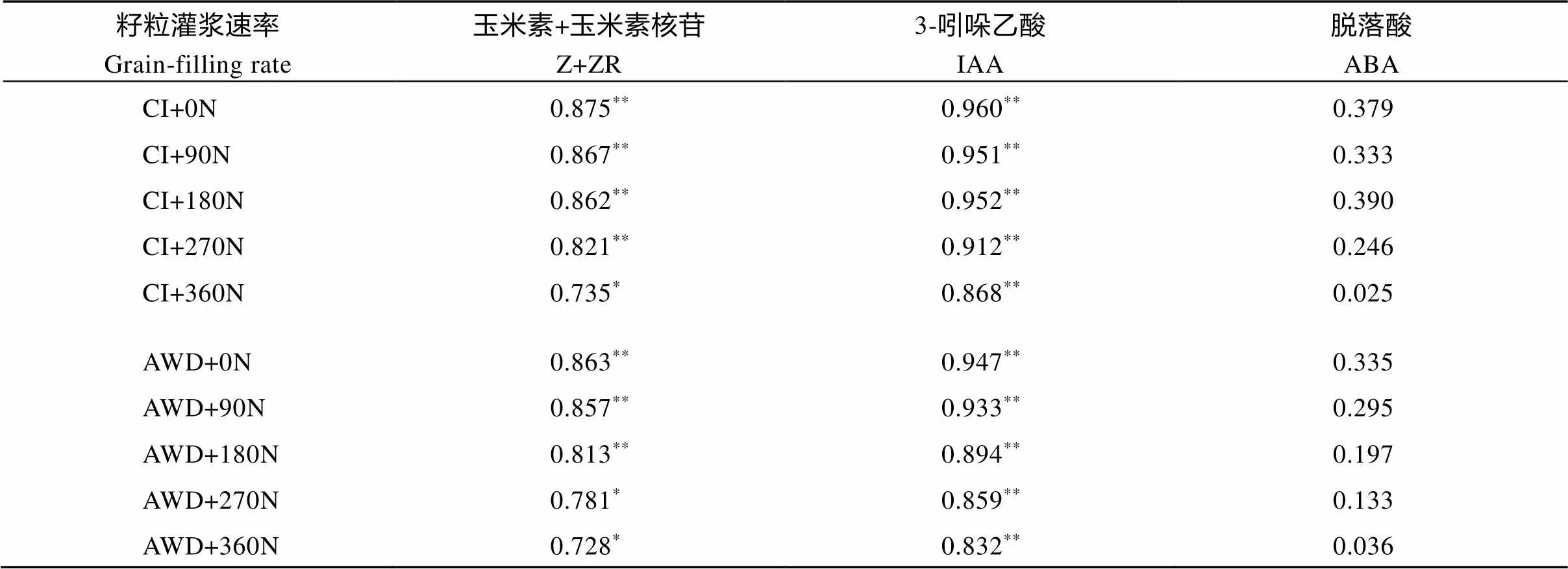
表6 水分和氮肥耦合下水稻籽粒灌浆速率与籽粒中激素含量的相关
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。*与**分别表示在0.05和0.01水平差异显著。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.*and**represent0.05 and0.01, respectively.
2.6 根系氧化力
由图4可知, 在抽穗开花期至花后30 d, 南粳9108的根系氧化力随籽粒灌浆时间增加而下降。其中花后10 d至20 d下降幅度较大。在同一灌溉方式下, 在0~ 360 kg hm–2施氮量范围内, 南粳9108的根系氧化力随施氮量的增加而提高, 施氮量270 kg hm–2和360 kg hm–2时差异不显著。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉显著提高了南粳9108的根系氧化力。
2.7 根系玉米素+玉米素核苷含量
抽穗开花期至花后30 d, 南粳9108的根系玉米素+玉米素核苷含量随籽粒灌浆时间增加而下降(图5)。其中花后10 d之后下降迅速。在同一灌溉方式下, 在0~270 kg hm–2施氮量范围内, 南粳9108的根系玉米素+玉米素核苷含量随施氮量的增加而显著提高, 在施氮量270~360 kg hm–2范围内时, 南粳9108的根系玉米素+玉米素核苷含量在抽穗开花期随施氮量的增加显著提高, 而在花后10 d之后差异不显著。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉显著提高了南粳9108的根系玉米素+玉米素核苷含量。
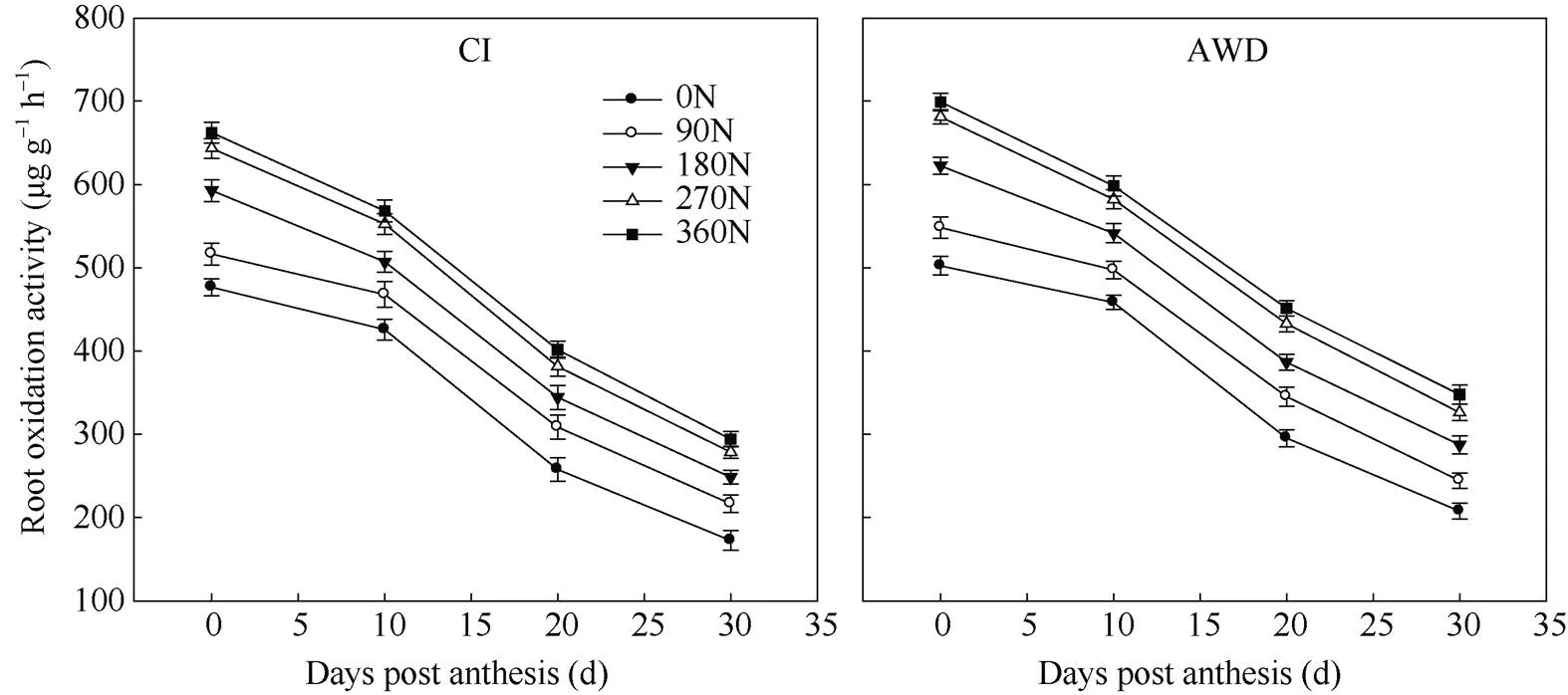
图4 水分和氮肥耦合下水稻根系氧化力变化
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.
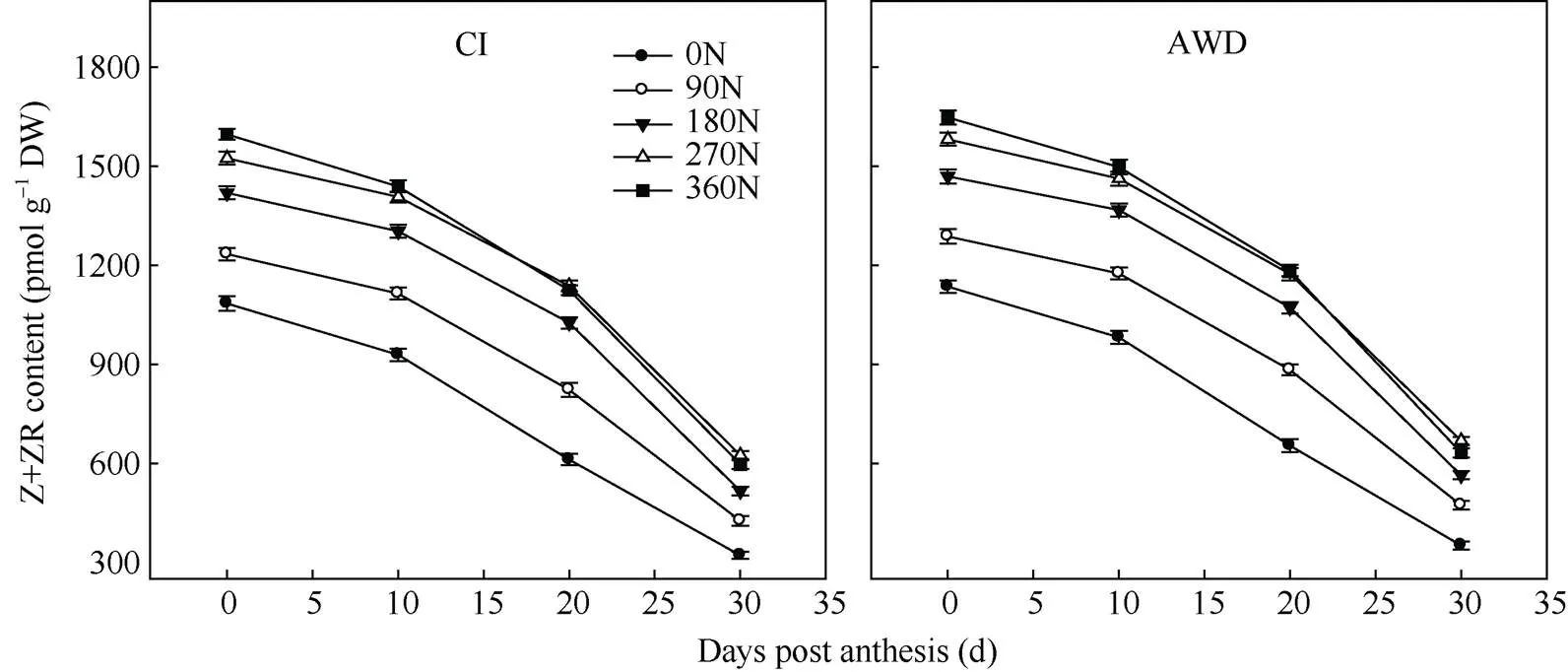
图5 水分和氮肥耦合下水稻根系玉米素及玉米素核苷含量的变化
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2.
2.8 茎鞘中NSC的运转
由表7可知, 在常规灌溉和干湿交替灌溉模式下, 在0~360 kg hm–2施氮量范围内, 南粳9108抽穗期和成熟期茎鞘中NSC含量随施氮量的增加显著增加。NSC运转率随施氮量增加降低, 除干湿交替灌溉模式下180N和270N无显著性差异。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉显著增加了抽穗期茎鞘中NSC含量, 降低了成熟期茎鞘中NSC含量, 提高了茎鞘中NSC转运率以及对产量的贡献率, 说明干湿交替灌溉可以促进水稻生育前期茎鞘中储存的NSC向籽粒的运转。
3 讨论
3.1 水分和氮肥耦合对水稻产量的影响
在水稻生长发育过程中, 水分和氮肥二因素往往是相互影响和相互制约的。前人关于水分或氮肥单因素对水稻产量调控的研究较多, 而有关水分和氮肥互作效应的研究较少, 且结论也不一致。有研究表明, 节水灌溉条件下, 适宜的氮肥运筹有利于提高水稻产量, 且水分和氮肥间存在显著的互作效应[9,13,20-21], 陈新红等[21]研究表明, 水稻抽穗开花期轻度土壤干旱时, 增施氮肥可以显著提高水稻产量; 程建平等[22]研究也证实, 轻度水分胁迫时, 水稻产量高低为高氮>中氮>低氮; 而当灌水充足或重度水分胁迫时, 则表现为中氮>高氮>低氮。也有研究表明, 干湿交替灌溉技术降低水稻产量, 水分和氮肥间并无互作效应, 在同一灌溉方式下, 施氮量在0~225 kg hm–2时, 产量随施氮量增加而增加, 但增产幅度显著下降, 施氮量为225~300 kg hm–2时, 产量随施氮量增加而下降[10]; Begg等[23]研究观点则认为, 在土壤水分有限条件下, 增施氮肥可使水分胁迫加重, 对作物产量造成不利的影响。这种正负效应可能是气候、品种的差异或土壤落干程度所致。本研究观察到, 南粳9108的产量及其构成因素在不同水分条件与施氮量间存在明显的互作效应。在灌溉方式和施氮量各处理的组合中, 干湿交替灌溉+270N处理的组合产量最高, 高产的原因是单位面积穗数和每穗粒数显著增加的幅度要大于结实率和千粒重减小的幅度。因此, 干湿交替灌溉+270N处理的组合为本试验最佳的水分和氮肥管理模式。
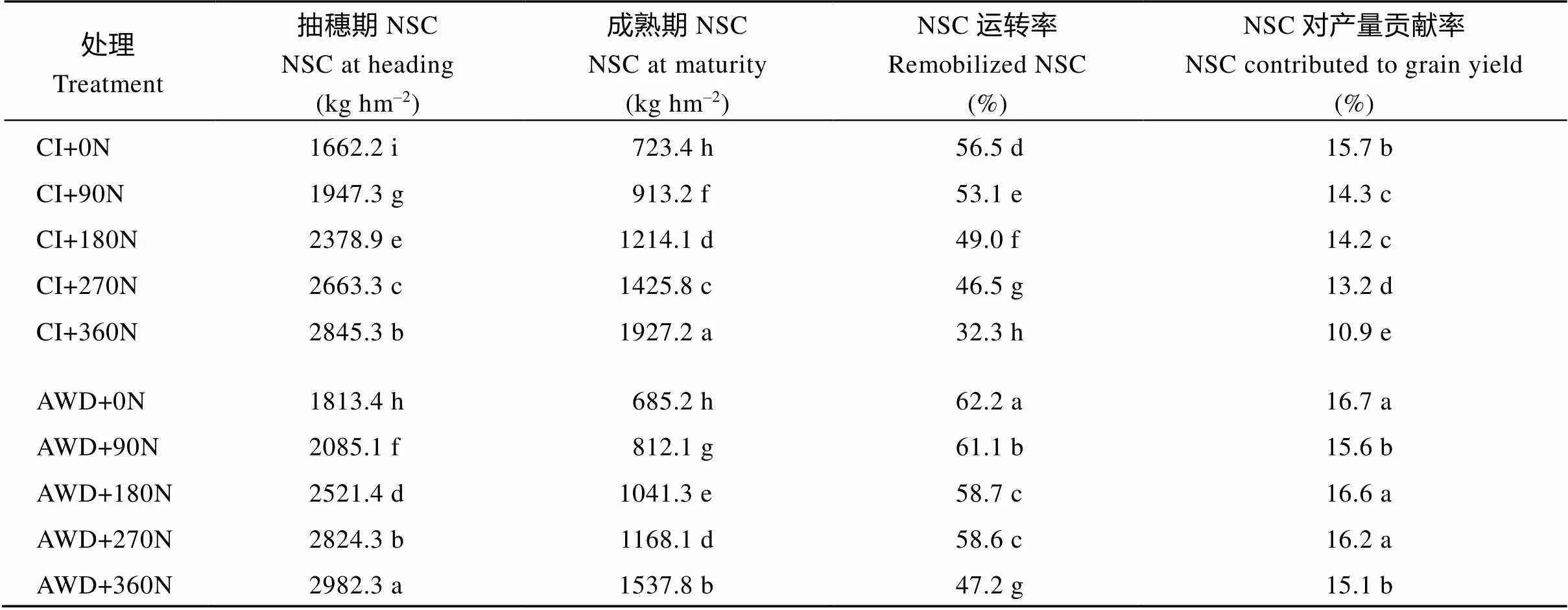
表7 水分和氮肥耦合对茎鞘中非结构性碳水化合物(NSC)运转的影响
CI: 常规灌溉; AWD: 干湿交替灌溉; 0N: 不施氮肥; 90N: 90 kg N hm–2; 180N: 180 kg N hm–2; 270N: 270 kg N hm–2; 360N: 360 kg N hm–2; 同一列标以不同字母表示= 0.05水平差异显著。
CI: the conventional irrigation; AWD: the alternate wetting and drying; 0N: no nitrogen applied; 90N: nitrogen 90 kg hm–2; 180N: nitrogen 180 kg hm–2; 270N: nitrogen 270 kg hm–2; 360N: nitrogen 360 kg hm–2; Different lowercase letters within the same column indicate significant difference at the0.05 probability level.
3.2 水分和氮肥耦合对水稻籽粒灌浆生理的影响
水稻籽粒灌浆过程受到水分和氮肥调控的影响,通常情况下, 水稻花前储存在茎鞘中的NSC对籽粒产量的贡献约占籽粒重量的1/6~1/3, 其量的多寡主要取决于水稻生长条件和氮素供应水平[24-25]。有研究表明, 水稻进入抽穗灌浆期后, 干旱胁迫会使植株早衰、物质生产减少、灌浆期缩短、粒重降低。虽然干旱胁迫可以增加茎中储存的NSC向籽粒转运,加速籽粒灌浆, 但茎中NSC向籽粒转运增加之得不能补偿物质生产减少和灌浆期缩短之失而造成粒重和产量降低[26-27]。Yang等[28]采用适度土壤干旱方法, 无严重抑制水稻植株的光合作用, 并且在夜间恢复植株水分状况, 即轻干湿交替灌溉处理, 可以使茎鞘中NSC向籽粒转运增加之得超过光合作用减少和灌浆期缩短之失, 这种处理不但不会显著降低籽粒灌浆速率, 反而会促进水稻籽粒灌浆, 从而提高粒重增加产量。本研究表明, 在2种灌溉方式下, 南粳9108的最大灌浆速率、平均灌浆速率和糙米重均随施氮量的增加而减小, 到达最大灌浆速率的时间推迟, 活跃灌浆期延长。其中施氮量在270~360 kg hm–2范围内时变化幅度较大。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高了南粳9108的最大灌浆速率、平均灌浆速率和糙米重, 延迟了到达最大灌浆速率的时间。
此外, 轻干湿交替灌溉可以提高籽粒中CTK含量, 促进胚乳细胞分裂, 茎秆中蔗糖合酶等淀粉合成关键酶活性增强, 促进营养组织中贮存同化物向籽粒调运, 灌浆速率显著增加, 起到增产效果。杨建昌[29]观察到, 花前储存在茎鞘中NSC不仅是籽粒灌浆物质的一部分, 而且是启动灌浆的重要物质基础, 其转运速率和转运量影响籽粒灌浆速率。脱落酸(ABA)通常被认为与植物衰老有关, Yang等[27,30-31]研究发现, ABA不仅能调控水稻的衰老, 而且能促进茎鞘中储存的NSC向籽粒的运转。花后适度的土壤干旱增加了内源ABA水平, 促进茎鞘中存储的NSC向籽粒运转。本研究观察到, 在2种灌溉方式下, 南粳9108的激素含量峰值随施氮量的增加出现推迟且减小, 之后籽粒激素含量下降速度减弱; 抽穗期和成熟期茎鞘中NSC含量随施氮量的增加显著增加, NSC运转率随施氮量增加而降低。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高了南粳9108的籽粒激素含量和茎鞘中NSC转运率以及对产量的贡献率, 说明干湿交替灌溉可以促进水稻生育前期茎鞘中储存的NSC向籽粒的运转。
水稻籽粒的主要成分是淀粉。在蔗糖-淀粉代谢途径中, 籽粒中蔗糖合成酶(SuSase)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合成酶(StSase)和淀粉分枝酶(SBE)在淀粉合成过程中起重要作用[32-34]。有研究认为, 谷类作物籽粒灌浆期干旱等逆境会降低籽粒蔗糖-淀粉代谢途径酶活性, 从而抑制籽粒库生理活性[35-36]。但Yang等[16,37]研究发现, 在水稻籽粒灌浆过程中, 适度干旱可以增强SuSase、StSase和SBE活性, 进而增加库强, 促进光合同化物向籽粒转运和籽粒中淀粉的合成。本研究观察到, 在2种灌溉方式下, 南粳9108的酶活性峰值随施氮量的增加出现推迟且峰值减小, 之后籽粒酶活性下降速度减弱。在同一氮肥水平下, 与常规灌溉相比, 干湿交替灌溉提高或显著提高了南粳9108的籽粒酶活性。
3.3 水分和氮肥耦合对水稻根系生理的影响
根系是作物吸收和运输水分和养分的重要器官之一, 对作物有支持、固定和保护的作用。水稻根系生长发育是水肥多因子交互作用的结果, 根系的形态及代谢特性与水分和养分吸收利用及地上部生长发育和产量关系密切。良好的根系生长有利于地上部生物量的积累并获得高产。建立强大的根系有利于抵御土壤干旱胁迫, 水分对根系的生长发育的影响直接影响着对养分的吸收[38]。重度干旱胁迫会抑制根系生长, 降低对氮素的吸收和运输能力[39]。水氮对根系的影响除了跟水氮营养有关, 还与根系是否受到胁迫有关, 土壤重度干旱和缺氮均不利于根系的生长[40]。轻度土壤干旱条件下增施氮素能够显著增加根干重、根体积和提高根系氧化力, 促进根系快速生长并扎深, 过度降低土壤水分则会对根系氧化力有抑制作用, 合理的水肥搭配有助于维持水稻根系活力, 延缓根系衰老[41]。本研究观察到, 抽穗开花后南粳9108的根系氧化力随施氮量的增加而提高, 施氮量270 kg hm–2和360 kg hm–2时差异不显著, 干湿交替灌溉显著提高了根系氧化力, 根系氧化力被认为是根系生理活性的重要指标。同时还观察到, 根系合成的细胞分裂素(玉米素和玉米素核苷)含量在抽穗开花期随施氮量的增加而显著提高, 花后10 d后, 在0~270 kg hm–2施氮量范围内随施氮量增加显著提高, 施氮量270 kg hm–2和360 kg hm–2时差异不显著。干湿交替灌溉条件下玉米素和玉米素核苷含量明显增加, 能够协调地上地下部生长, 促进水稻籽粒灌浆, 延缓植株衰老, 利于产量形成。
4 结论
水稻籽粒灌浆生理及根系生理在不同水氮耦合间存在明显差异。270 kg hm–2施氮量和干湿交替灌溉处理协同地上地下生长, 灌浆期具有较高的籽粒灌浆速率和激素含量, 能够维持较高的籽粒酶活性。根系具有较强的氧化力, 根系细胞分裂素含量增加。表明通过适宜的水氮耦合提高水稻根系生理代谢能力, 协调地上地下部生长, 将有利于籽粒灌浆及增产。
[1] 朱兆良, 金继运. 保障我国粮食安全的肥料问题. 植物营养与肥料学报, 2013, 19: 259–273.
Zhu Z L, Jin J Y. Fertilizer use and food security in China., 2013, 19: 259–273 (in Chinese with English abstract).
[2] Peng S B, Tang Q Y, Zou Y B. Current status and challenges of rice production in China., 2009, 12: 3–8.
[3] Tilman D K, Cassman K G, Matson P A. Agricultural sustain-ability and intensive production practices., 2002, 418: 671–678.
[4] Peng S B. Water resources strategy and agricultural development in China., 2011, 6: 1709–1713.
[5] Zhang J H. China’s success in increasing per capita food production., 2011, 62: 3707–3711.
[6] Kukal S S, Hira G S, Sidhu A S. Soil matric potential-based irrigation scheduling to rice ()., 2005, 23: 153–159.
[7] Carrijo D, Lundy M, Linquist B. Rice yields and water use under alternate wetting and drying irrigation: a meta-analysis., 2017, 203: 173–180.
[8] 孙永健. 水氮互作对水稻产量形成和氮素利用特征的影响及其生理基础. 四川农业大学博士学位论文, 四川温江, 2010.
Sun Y J. Effects of Water-nitrogen Interaction on Yield Formation and Characteristics of Nitrogen Utilization in Rice and its Physiological Basis. PhD Dissertation of Sichuan Agricultural University, Wenjiang, Sichuan, China, 2010 (in Chinese with English abstract).
[9] 杨建昌, 王志琴, 朱庆森. 不同土壤水分状况下氮素营养对水稻产量的影响及其生理机制的研究. 中国农业科学, 1996, 29(4): 58–66.
Yang J C, Wang Z Q, Zhu Q S. Effect of nitrogen nutrition on rice yield and its physiological mechanism under different status of soil moisture., 1996, 29(4): 58–66 (in Chinese with English abstract).
[10] Cabangon R, Tuong T, Castillo E. Effect of irrigation method and N-fertilizer management on rice yield, water productivity and nutrient use efficiencies in typical lowland rice conditions in China., 2004, 2: 195–206.
[11] Santiago-Arenas R, Hadi S N, Fanshuri B A, Ullah H, Datta A. Effect of nitrogen fertilizer and cultivation method on root systems of rice subjected to alternate wetting and drying irrigation., 2019, 175: 388–399.
[12] 徐国伟, 陆大克, 王贺正, 陈明灿, 李友军. 干湿交替灌溉与施氮量对水稻叶片光合性状的耦合效应. 植物营养与肥料学报, 2017, 23: 1225–1237.
Xu G W, Lu D K, Wang H Z, Chen M C, Li Y J. Coupling effect of wetting and drying alternative irrigation and nitrogen application rate on photosynthetic characteristics of rice leaves., 2017, 23: 1225–1237 (in Chinese with English abstract).
[13] 褚光, 陈婷婷, 陈松, 徐春梅, 王丹英, 章秀福. 灌溉模式与施氮量交互作用对水稻产量以及水、氮利用效率的影响. 中国水稻科学, 2017, 31: 513–523.
Chu G, Chen T T, Chen S, Xu C M, Wang D Y, Zhang X F. Effects of interaction between irrigation regimes and nitrogen rates on rice yield and water and nitrogen use efficiencies., 2017, 31: 513–523 (in Chinese with English abstract).
[14] 朱庆森, 曹显祖, 骆亦其. 水稻籽粒灌浆的生长分析. 作物学报, 1988, 14: 182–193.
Zhu Q S, Cao X Z, Luo Y Q. Growth analysis on the process of grain filling in rice., 1988, 14: 182–193 (in Chinese with English abstract).
[15] Richards F J. A flexible growth function for empirical use., 1959, 10: 290–300.
[16] Yang J C, Zhang J H, Wang Z Q, Zhu Q S, Liu L J. Activities of enzymes involved in sucrose-to-starch metabolism in rice grains subjected to water stress during filling., 2003, 81: 69–81.
[17] Bollmark M, Kubat B, Eliasson L. Variations in endogenous cytokinin content during adventitious root formation in pea cuttings., 1988, 132: 262–265.
[18] Zhang H, Xue Y G, Wang Z Q, Yang J C, Zhang J H. Morphological and physiological traits of roots and their relationships with shoot growth in “super” rice., 2009, 113: 31–40.
[19] 杨建昌, 杜永, 刘辉. 长江下游稻麦周年超高产栽培途径与技术. 中国农业科学, 2008, 41: 1611–1621.
Yang J C, Du Y, Liu H. Cultivation approaches and techniques for annual super-high-yielding of rice and wheat in the lower reaches of Yangtze River., 2008, 41: 1611–1621 (in Chinese with English abstract).
[20] Sandhu S S, Mahal S S, Vashist K K, Buttar G S, Brar A S, Singh Maninder. Crop and water productivity of bed transplanted rice as influenced by various levels of nitrogen and irrigation in northwest India., 2012, 104: 32–39.
[21] 陈新红. 土壤水分与氮素对水稻产量和品质的影响及其生理机制. 扬州大学博士学位论文, 江苏扬州, 2004.
Chen X H. Effects of Soil Moisture and Nitrogen Nutrient on Grain Yield and Quality of Rice and Their Physiological Mechanism. PhD Dissertation of Yangzhou University, Yangzhou, Jiangsu, China, 2004 (in Chinese with English abstract).
[22] 程建平, 曹凑贵, 蔡明历, 原保忠, 翟晶. 不同土壤水势与氮素营养对杂交水稻生理特性和产量的影响. 植物营养与肥料学报, 2008, 14: 199–206.
Cheng J P, Cao C G, Cai M L, Yuan B Z, Zhai J. Effect of different nitrogen nutrition and soil water potential on physiological parameters and yield of hybrid rice., 2008, 14: 199–206 (in Chinese with English abstract).
[23] Begg J E, Turner N C. Crop and water deficits., 1976, 28: 161–218.
[24] Li G H, Pan J F, Cui K H, Yuan M S, Hu Q Q, Wang W C, Mohapatra P K, Nie L X, Huang J L, Peng S B. Limitation of unloading in the developing grains is a possible cause responsible for low stem non-structural carbohydrate translocation and poor grain yield formation in rice through verification of recombinant inbred lines., 2017, 8: e1369.
[25] Okamura M, Arai-Sanoh Y, Yoshida H, Mukouyama T, Adachi S, Yabe S, Nakagawa H, Tsutsumi K, Taniguchi Y, Kobayashi N, Kondo M. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield., 2018, 219: 139–147.
[26] Asseng S, van Herwaarden A F. Analysis of the benefits to wheat yield from assimilates stored prior to grain filling in a range of environments., 2003, 256: 217–229.
[27] Yang J C, Zhang J H, Wang Z Q, Zhu Q S, Liu L J. Water deficit-induced senescence and its relationship to the remobilization of pre-stored carbon in wheat during grain filling., 2001, 93: 196–206.
[28] Yang J C, Zhang J H, Wang Z Q, Zhu Q S, Wang W. Remobilization of carbon reserves in response to water-deficit during grain filling of rice., 2001, 71: 47–55.
[29] 杨建昌. 亚种间杂交稻籽粒充实特征及其生理基础研究. 中国农业大学博士学位论文, 北京, 1996.
Yang J C. Characteristics and Physiological Bases of Grain Filling in Intersubspecific Hybrid Rice. PhD Dissertation of Chinese Agricultural University, Beijing, China, 1996 (in Chinese with English abstract).
[30] Yang J C, Zhang J H, Ye Y X, Wang Z Q, Zhu Q S, Liu L J. Involvement of abscisic acid and ethylene in the responses of rice grains to water stress during filling., 2004, 27: 1055–1064.
[31] 杨建昌, 张建华. 促进稻麦同化物转运和籽粒灌浆的途径与机制. 科学通报, 2018, 63: 2932–2943.
Yang J C, Zhang J H. Approach and mechanism in enhancing the remobilization of assimilates and grain-filling in rice and wheat., 2018, 63: 2932–2943 (in Chinese with English abstract).
[32] Beck E, Ziegler P. Biosynthesis and degradation of starch in higher plants., 1989, 40: 95–117.
[33] Tian B, Talukder S K, Fu J M, Fritz A K, Trick H N. Expression of a rice soluble starch synthase gene in transgenic wheat improves the grain yield under heat stress conditions., 2018, 54: 216–227.
[34] Zi Y, Ding J F, Song J M, Humphreys G, Peng Y X, Li C Y, Zhu X K, Guo W S. Grain yield, starch content and activities of key enzymes of waxy and non-waxy wheat (L.)., 2018, 8: e4548.
[35] Ahmadi A, Baker D A. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat., 2001, 35: 81–91.
[36] Boyer J S, Westgate M E. Grain yields with limited water., 2004, 55: 2385–2394.
[37] Yang J C, Zhang J H, Wang Z Q, Xu G W, Zhu Q S. Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling., 2004, 135: 1621–1629.
[38] 占爱. 提高养分、水分吸收的根系形态和生理调控. 西北农林科技大学博士学位论文, 陕西杨凌, 2015.
Zhan A. Root Morphology and Physiological Regulation to Improve Nutrient and Water Absorption. PhD Dissertation of Northwest A&F University, Yangling, Shaanxi, China, 2015 (in Chinese with English abstract).
[39] Wang H, Siopongco J, Wade L J, Yamauchi A. Fractal analysis on root systems of rice plants in response to drought stress., 2009, 65: 338–344.
[40] 徐国伟, 王贺正, 翟志华, 孙梦, 李友军. 不同水氮耦合对水稻根系形态生理、产量与氮素利用的影响. 农业工程学报, 2015, 31(10): 132–141.
Xu G W, Wang H Z, Zhai Z H, Sun M, Li Y J. Effect of water and nitrogen coupling on root morphology and physiology, yield and nutrition utilization for rice., 2015, 31(10): 132–141 (in Chinese with English abstract).
[41] 张凤翔, 周明耀, 周春林, 钱晓晴. 水肥耦合对水稻根系形态与活力的影响. 农业工程学报, 2006, 22(5): 197–200.
Zhang F X, Zhou M Y, Zhou C L, Qian X Q. Effects of water and fertilizer coupling on root morphological characteristics and activities of rice., 2006, 22(5): 197–200(in Chinese with English abstract).
Effects of alternate wetting and drying irrigation and nitrogen coupling on grain filling physiology and root physiology in rice
FU Jing1, WANG Ya1, YANG Wen-Bo1, WANG Yue-Tao1, LI Ben-Yin2, WANG Fu-Hua1, WANG Sheng-Xuan1, BAI Tao1, and YIN Hai-Qing1,*
1Cereal Research Institute, Henan Academy of Agricultural Sciences, Zhengzhou 450002, Henan, China;2Institute of Plant Nutrition, Agricultural Resources and Environmental Science, Henan Academy of Agricultural Sciences, Zhengzhou 450002, Henan, China
Soil moisture and nitrogen nutrient are the two principal factors affecting rice production. Alternating dry and wet irrigation (AWD) coupled with nitrogen application plays important roles in root growth and yield formation in rice. However, its effect on grain filling physiology and their relationship with root physiology is not clearly understood. To explore the effects of alternating dry and wet irrigation and nitrogen application coupling on grain filling, changes of key enzyme activities involved in sucrose-to starch conversion and hormone contents, and root physiology of rice, the super rice variety Nanjing 9108 was cultivated as the material in the field. The field experiments were conducted with two irrigation regimes [conventional irrigation (CI) and AWD] and five nitrogen application rates [no nitrogen fertilizer (0N), 90 kg hm–2(90N), 180 kg hm–2(180N), 270 kg hm–2(270N), and 360 kg hm–2(360N) nitrogen fertilizer]. The results showed that there was a significant interaction effect between irrigation method and nitrogen application rate. Alternating dry and wet irrigation increased the maximum grain filling rate and average grain filling rate of Nanjing 9108, and increased the activities of the sucrose synthase enzymes, adenosine diphosphate glucose pyrophosphorylation enzymes, starch synthase enzymes, and starch branching enzymes, and the contents of zeatin + zeatin riboside, 3-indoleacetic acid and abscisic acid, increased root oxidation activity, and the content of zeatin + zeatin riboside in roots after heading, promoted the transfer of NSC stored in the stem sheath to the grain in the early growth stage of rice, and the highest yield was obtained after coupling with 270N, which was the best water-nitrogen coupling mode in this experiment. In conclusion, the water-nitrogen coupling effect can be exerted by appropriate regulation of water and fertilizer, which can improve the physiological performance of rice roots and grain-filling physiological activity, and achieve high rice yield.
rice; the alternate wetting and drying irrigation; nitrogen rate; grain filling physiology; root physiology
10.3724/SP.J.1006.2023.22032
本研究由河南省科技攻关计划项目(222102110019), 河南省农业科学院自主创新基金项目(2022ZC09), 河南省现代农业产业技术体系建设专项资金项目(HARS-22-03-S), 河南省农业科学院水稻科技创新团队项目(2022TD29)和河南省农业科学院基础性科研工作项目(2022JC05)资助。
This study was supported by the Henan Provincial Science and Technology Research Program (222102110019), the Henan Academy of Agricultural Sciences Independent Innovation Fund Project (2022ZC09), the Special Fund for Henan Agriculture Research System (HARS-22-03-S), the Henan Academy of Agricultural Sciences Rice Science and Technology Innovation Team Project (2022TD29), and the Fundamental Research Project of Henan Academy of Agricultural Sciences (2022JC05).
通信作者(Corresponding author):尹海庆, E-mail: yinhq98@163.com
E-mail: fujing8210@sina.cn
2022-05-13;
2022-07-21;
2022-08-12.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20220811.1844.002.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
现代畜牧科技(2021年4期)2021-12-05 15:34:45
现代畜牧科技(2021年10期)2021-11-19 08:42:26
中国化肥信息(2021年5期)2021-07-28 07:05:00
中国化肥信息(2019年4期)2019-05-31 10:13:02
中国化肥信息(2019年3期)2019-04-25 01:56:14
河北农业科学(2018年2期)2018-07-26 11:28:14
中国化肥信息(2017年3期)2017-12-23 06:29:25
商洛学院学报(2017年2期)2017-05-17 05:19:53
植物营养与肥料学报(2014年1期)2014-03-11 20:24:27
植物营养与肥料学报(2012年2期)2012-10-26 02:52:24
