
Tempo-spatial distribution of Ulva spp. micro-propagules in the Yellow Sea during and after green tide in 2019*
2023-01-04 03:04JinZHAOFanzhouKONGQianchunLIUFengjieLIXiuWEITianYANPengJIANG
Jin ZHAO , Fanzhou KONG , Qianchun LIU ,5, Fengjie LI , Xiu WEI ,6,Tian YAN , Peng JIANG ,**
1 CAS and Shandong Province Key Laboratory of Experimental Marine Biology, Center for Ocean Mega-Science, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
2 Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology (Qingdao),Qingdao 266237, China
3 CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
4 Laboratory of Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China
5 University of Chinese Academy of Sciences, Beijing 100049, China
6 College of Life Science, Qingdao University, Qingdao 266071, China
Abstract Large-scale green tides occur frequently in summer in the Yellow Sea in recent decade. Micropropagules, providing “seeds” for the seaweeds, play an important role in the formation of green tide. We studied the distribution pattern and genetic composition of the Ulva micro-propagules in the Yellow Sea during and after the green tide in 2019. In April, a great amount of Ulva micro-propagules was detected in the Subei Shoal, with the average density of 908 inds./L, being similar to that of previous studies. U. prolifera was found out at a low percentage (4%), and 25% of them were positive when tested by the specif ic marker for the f loating ecotype. The percentage of U. prolifera was obviously lower than that in the same season from 2009 to 2016. Therefore, the quantity of U. prolifera micro-propagules sharply decreased. Signif icant variation in composition of the micro-propagules in Subei radial sand ridges was revealed in the background of the alternation of macroalgal tides in the Yellow Sea in the latest years. During the green tide in June and July,Ulva micro-propagules were detected from some nearshore sites where the f loating seaweeds distributed, and U. prolifera was the dominate species. In September, when the green tide ended, no Ulva micro-propagule was detected from the open sea area of the Yellow Sea. It is indicated that the green algae micro-propagules were unlikely to exist in the open water of the Yellow Sea for a long time according to the large-scale surveys. The mechanism of preserving micro-propagules in Subei radial sand ridges, and the destination of U. prolifera micro-propagules in the open sea area of the Yellow Sea after green tide deserves further study.
Keyword: green tide; Ulva prolifera; micro-propagule; Yellow Sea
1 INTRODUCTION
The Yellow Sea green tide re-occurred frequently in spring and summer in the recent decade since 2007,which brought serious troubles to both the ecology and the economy. The f loating ecotype ofUlvaproliferahas been proved to be the dominant algae(Zhao et al., 2013, 2015). It was proved that Subei radial sand ridges are the geographical origin for the green tide as seen from satellite remote sensing data and cruise surveys (Keesing et al., 2011; Zhang et al., 2014; Wang et al., 2015; Xing et al., 2019). It is suggested that the micro-propagules ofUlvaspp. in the environment, including gametes, spores, zygotes,micro-germlings, and vegetative fragments, can attach to the nori aquaculture facilities, grow up and accumulate their initial biomass (Huo et al., 2014;Song et al., 2018). During the nori harvest, green algae were cleaned up from the facilities and became f loating at seawater surface, which is thought to be the direct source of the bloom-forming algae (Liu et al., 2009, 2010a, 2021a, b, 2022; Zhang et al., 2019).Micro-propagules, which is considered as the “seed bank” of green algae, play an important role to the annual occurrence of green tide (Liu et al., 2012,2013).
Based on the studies carried out between 2009 to 2016, green algal micro-propagules were persistently detected in the environment in Subei radial sand ridges in the year (Liu et al., 2012, 2013, 2017; Li et al., 2014; Song et al., 2015; Huo et al., 2016;Miao et al., 2020).U.proliferawas found to be the dominant species, accounting more than 50% of the micro-propagules in April (Liu et al., 2013; Huo et al., 2016; Song et al., 2018). However, most of previous studies paid little attention to the genetic diversity at the intra-species level ofU.prolifera.It has been proved that the Yellow Sea green tide is dominated by a unique ecotype ofU.prolifera,which is genetically diff erent from the attached populations widely distributing along the coastline of Jiangsu Province (Zhang et al., 2011; Zhao et al., 2015). The proportion of the f loating ecotype ofU.proliferamicro-propagules in Subei radial sand ridges was reported in May 2016 only, accounting for approximately 88% (Miao et al., 2020). It is necessary to do further study at intra-species level to compare the inter-annual variation in quantity of the bloom-forming algae. Moreover, most of the reports on the tempo-spatial distribution of the micropropagules in Subei radial sand ridges were before 2017. It was reported that the macroalgal blooms in the Yellow Sea varied obviously in recent years,and the golden tide caused bySargassumhornerico-occurred with the green tide every summer since 2017 (Zhang et al., 2019; Xiao et al., 2020a). A great amount of strandedS.horneriwere found from nori aquaculture facilities and might aff ect the epiphytic green algae on the rafts, which were considered to be the contributors to the micro-propagules to the water column (Li et al., 2014; Miao et al., 2020; Xiao et al., 2020b). Therefore, the community structure of green algal micro-propagules in Subei radial sand ridges might be altered, which deserves more indepth studies.
During the northward drifting of the f loating seaweeds, micro-propagules were detected in the open sea area of the Yellow Sea (Li et al., 2014;Huo et al., 2016). The f loating matured thalli are considered as the donor of micro-propagules, andU.proliferawas the dominant species in this stage(Li et al., 2014; Miao et al., 2020). These studies covered the blooming period of the Yellow Sea green tide. However, the tempo-spatial distribution pattern ofU.proliferamicro-propagules after green tide in the Yellow Sea was poorly studied. At present, only a few studies paid attention to the existence time of micro-propagules along Qingdao coastline (Liu et al.,2010b; Miao et al., 2018; Zhao et al., 2018), and the condition in the open sea area is unknown.
In this study, the tempo-spatial distribution and genetic composition at both the inter- and intra-species level of green algae micro-propagules in the Yellow Sea during and after green tide in 2019 were studied.In addition, previous studies on the micro-propagules in the Yellow Sea before 2019 were reviewed and compared with the new data. The aims are (1) to investigate the potential variation of the micropropagule composition in Subei radial sand ridges in response to the alternation of macroalgal tides in the Yellow Sea in the latest years; (2) to specify the duration of micro-propagules of the f loating ecotype after green tide in the open sea area of the Yellow Sea,and (3) to speculate the f inal destination of the micropropagules left from f loating thalli.
2 MATERIAL AND METHOD
2.1 Cruise investigation and sampling sites
Four survey cruises, including a nearshore cruise in the Subei Shoal and three major cruises covering a large open sea area of the Yellow Sea, were conducted to investigate the distribution and species diversity of green algae micro-propagules during and after the Yellow Sea green tide in 2019. The nearshore cruise was conducted in mid-April, 2019, when green tide just started. Two of the three major cruises were conducted during green tide in the mid-June and late July, respectively. And the last one was carried out in the early September, when the green tide ended,to investigate the duration of the micro-propagules in the open sea environment of the Yellow Sea. Seawater and surface sediment samples were collected during the cruises (Fig.1; Supplementary Table S1), and stored in cooling boxes before being transported to the laboratory.

Fig.1 Deployment of sampling sites of the four research cruises in this study
2.2 Micro-propagule cultivation from the seawater and sediment samples
Green algae micro-propagules in the seawater and sediment samples were cultivated using the method reported by Liu et al. (2013) with some modif ications as described by Zhao et al. (2018). For seawater,500 mL of each sample was enriched with 500 μL of 1 000× VSE (von Stosch’s enriched) solution in 500-mL glass beaker. For sediments, a wet weight of 100 g of each sample was put into a 1 000-mL glass beaker f illed with 500-mL sterile VSE medium. The sediment suspension was stirred vigorously and then the liquid was f iltered out with the Nylon mesh (with the bore size of 75 μm) into a 500-mL glass beaker.Saturated GeO2solution of 250 μL was added in each beaker to inhibit the growth of diatoms (Shea and Chopin, 2007). Triplicated treatments were set up for each seawater or sediment sample in an incubator (Jiangnan, Ningbo, China) at 20 °C with 100 μmol photons/(m2·s) and 12 h꞉12 h light꞉dark light cycle. After three-week culture, the number of green algae germlings was counted. The abundance of micro-propagules (A, inds./L) was calculated asA=N/VorA=N/m(N: total number of germlings;V: volume of seawater sample cultured;m: weight of sediment sample cultured). The individual thallus was randomly selected from each sample for further DNA extraction and species identif ication.
2.3 Molecular identif ication
Genomic DNA was extracted from individual thallus using a Plant Genomic DNA Kit (TIANGEN,Beijing, China) following the manufacture’s protocols.All of the specimens were identif ied by their internal transcribed spacer (ITS) sequences (Leskinen and Pamilo, 1997). Then, for the LPP (U.linza-proceraprolifera) clade, morphological features and 5S rDNA spacer sequences were analyzed to separateU.linzaandU.prolifrea(Shimada et al., 2008; Tseng, 2009).The most prominent morphological feature ofU.proliferais the dense branching of the tubular thallus,whileU.linzahas a distromatic thallus with bilateral thin tubular margins. The sequence characterized amplif ied region (SCAR) marker developed by Zhao et al. (2015) was used to trace the unique f loating ecotype ofU.proliferadominating in the Yellow Sea green tide. A specif ic 830-bp band can be amplif ied from the f loating ecotype, while is absent from the attached populations. PCR products were checked and purif ied on 1.5% Tris Acetate-EDTA (TAE) agarose gels stained with DuRed (US Everbright Inc.) and sequenced by Sangon Biotech (Shanghai) Co., Ltd.Sequences were aligned using ClustalX (1.83) (Larkin et al., 2007). Eighteen ITS sequences fromUlva,Blidingia, andUrosporaspecies and ten 5S rDNA space sequences from LPP clade are downloaded from GenBank as references to elucidate the relationship among the algal samples. Phylogenetic trees are built using Maximum Likelihood (ML) and Neighbor-Joining (NJ) methods with Mega (4.0) (Tamura et al.,2007). Robustness of the phylogenetic tree was tested by boot strapping with 1 000 replicates of the data.
3 RESULT
3.1 Phylogenetic analysis of individual specimens cultivated from micro-propagules
A total number of 238 green algae seedlings cultured from the seawater and sediment samples were selected for molecular identif ication. ML analysis of the ITS sequences showed that all the individual samples fell into three distinct clades: the LPP clade(160 samples),U.aragoënsisclade (75 samples), andU.compressaclade (3 samples, Fig.2). Phylogenetic tree built by NJ method shows similar topological structures to the ML tree (Supplementary Fig.S1). It should be noted that the species, which has long been identif ied asU.f lexuosaaccording to the ITS sequence,was just corrected toU.aragoënsisrecently (Hiraoka et al., 2017; Krupnik et al., 2018), and the sequences from the micro-propagule samples belonging to this complex were previously named afterU.f lexuosa(Liu et al., 2012, 2013; Huo et al., 2016; Song et al., 2018).Morphological and 5S rDNA spacer identif ication of the samples in the LPP clade show that 38 out of the 160 LPP samples wereU.linza, and the remaining 122 samples wereU.prolifera(Supplementary Fig.S2). Based on the PCR results of the specif ic SCAR marker, the f loating ecotype accounted for 81% of theU.proliferaindividual samples.
3.2 Density and species composition of green algae micro-propagules in seawater of the Subei Shoal in April 2019
A nearshore cruise was conducted on April 18–22, 2019, to investigate the density and species composition of the green algae micro-propagules in the Subei Shoal at the initially formatting stage of the green tide. Seawater samples were collected from seven sites. The density of the micro-propagules ranged 178–2 104 inds./L on average of 908 inds./L.The highest micro-propagule density was found in two sites around the 33°N transect (Fig.3a;Supplementary Table S1). Four species were detected from the green algae micro-propagules, includingU.aragoënsis(61%),U.linza(33%),U.prolifera(4%), andU.compressa(2%).U.aragoënsisandU.linzawere the major species of green algae micropropagules in the Subei Shoal during survey in this study.U.proliferamicro-propagules were detected at a low percentage and 25% of theU.proliferaindividuals were SCAR-positive (Fig.3b).
3.3 Density and community composition of green algae micro-propagules in the Yellow Sea during and after green tide
During June 11–19, 2019, seawater and surface sediment were sampled in 20 and 21 sites respectively along f ive transects (33°00ʹN, 34°00ʹN, 35°00ʹN,36°00ʹN, and 36°30ʹN). In the seawater samples,after three-week cultivation,Ulvasprouts grew out in f ive samples and the micro-propagule density ranged from 0 to 60 inds./L (Fig.4a; Supplementary Table S1). Three species were detected, i.e.,U.prolifera(85%),U.aragoënsis(14%), andU.compressa(1%) (Fig.4b). In the sediment samples,Ulvamicropropagules were detected from three samples, in which the micro-propagule density ranged from 0 to 40 inds./kg (Fig.4c; Supplementary Table S1).Three species were detected, i.e.,U.prolifera(59%),U.linza(33%), andU.aragoënsis(8%) (Fig.4d).U.proliferawas the dominantUlvaspecies in both seawater and sediment samples. Eighty-four percentages ofU.proliferaindividuals were f loating ecotype as revealed by the SCAR marker detection.
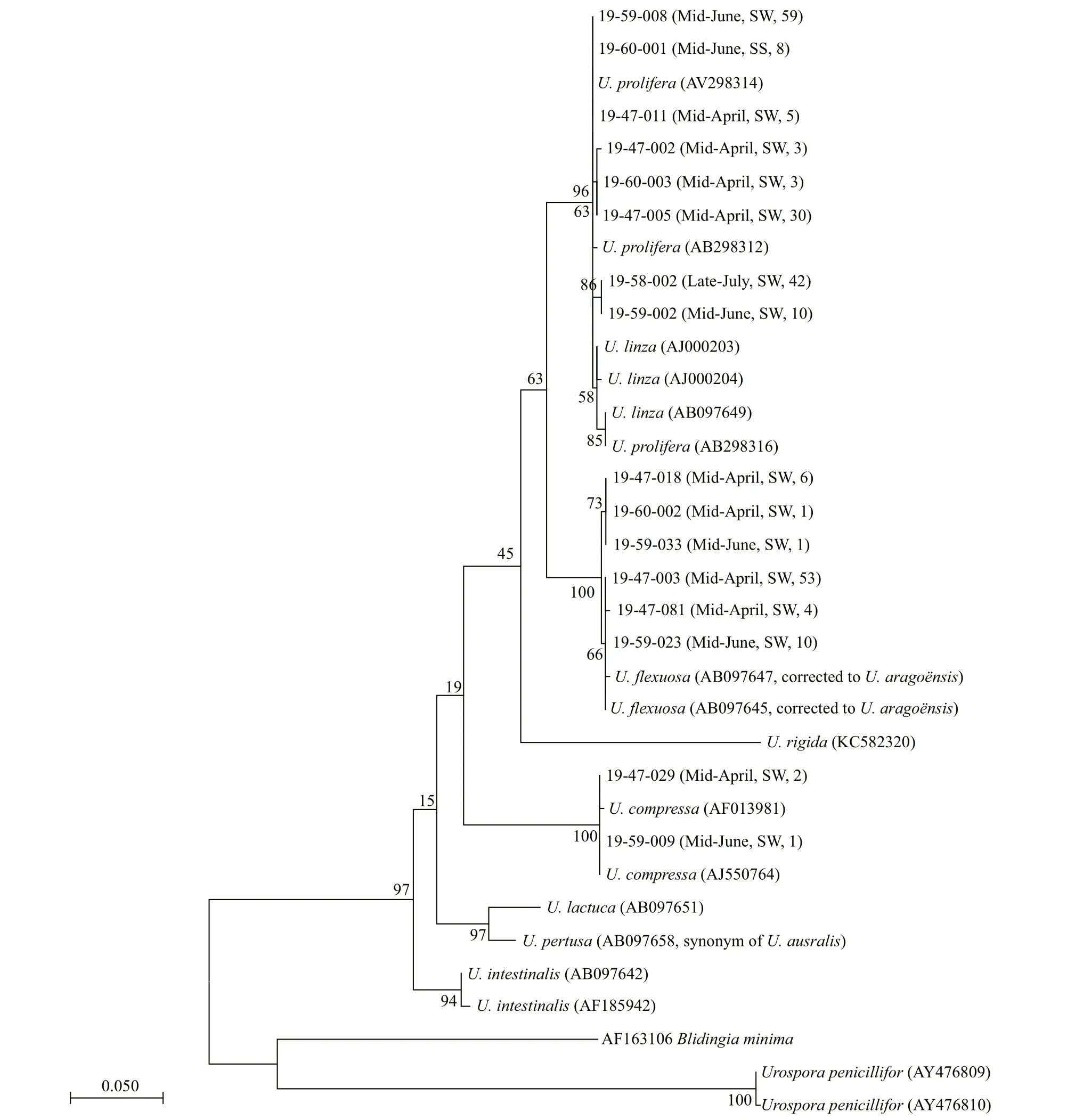
Fig.2 Phylogenetic tree of the ML analysis inferred from the ITS sequences of the green algae seedlings cultured form the seawater and sediment samples
Twenty-two seawater samples and 26 surface sediment samples were collected from six transects(33°00ʹN, 34°00ʹN, 35°00ʹN, 36°00ʹN, 36°30ʹN,and 37°00ʹN) on July 21–28, 2019.Ulvagermlings grew out from f ive seawater samples. The micropropagule density ranged from 0 to 62 inds./L(Fig.5a; Supplementary Table S1). Only one species,U.proliferawas detected from the germlings, and 81%of theU.proliferaindividuals were SCAR-positive(Fig.5b). NoUlvamicro-propagule was detected from all the sediment samples (Supplementary Table S1).
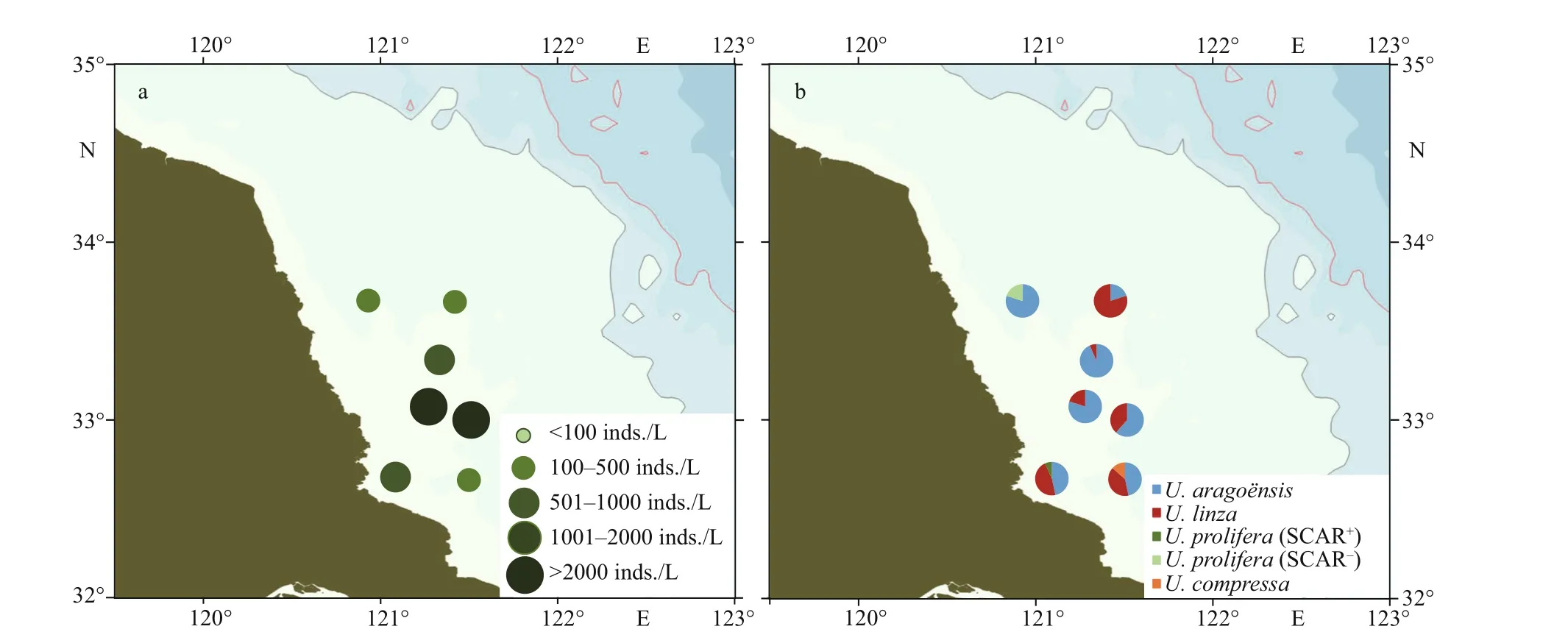
Fig.3 Density (a) and community composition (b) of green algae micro-propagules in seawater of the Subei Shoal in mid-April, 2019
To investigate the duration of the micro-propagules existence in the open sea environment of the Yellow Sea, a cruise was carried out on September 3–6, 2019 when green tide ended. Seawater and surface sediment samples were collected from nine sites along three transects (36°00ʹN, 36°30ʹN, and 37°00ʹN). However,Ulvamicro-propagule was not detected from all of the samples (Supplementary Table S1).
4 DISCUSSION
4.1 Signif icant variation in community composition of micro-propagules in Subei radial sand ridges under the background of the alternation of macroalgal tides
In the present study, a great number of green algae micro-propagules were detected in all the investigation sites in the Subei Shoal in April, 2019,with the average density of 908 inds./L. The high density of micro-propagules was in accordance with the results of many former studies in this sea area in the season. In the Aprils of 2012 and 2013,Ulvamicro-propagule densities were reported to be 4 800 inds./L and 1 189 inds./L, respectively (Huo et al., 2014, 2016). On the other hand, it was found that the composition percentages of diff erentUlvaspp. micro-propagules in April 2019 in this area varied dramatically from that in the same season of previous years. In this study,U.aragoënsisandU.linzawere found the major species of green algae micro-propagules in the Subei Shoal, andU.proliferamicro-propagules were detected at a low percentage(4%) during the investigation period in April 2019. From 2009 to 2014, studies on the species composition ofUlvamicro-propagules in this region found repeatedly that the proportion ofU.proliferawas more than 50%, andUlvaspecies was dominant in April (Song et al., 2015; Huo et al., 2016). In May,2016,U.proliferawas the second dominant species,composing 35% of the total green algae micropropagules in the Subei Shoal (Miao et al., 2020). The quantity ofU.proliferamicro-propagules in the Subei Shoal sharply decreased in April 2019 from that in the same season of earlier years from 2009 to 2016.
It was considered that the attached green macroalgae on nori aquaculture rafts were the only or substantial contributors to the micro-propagules in water column of the Subei Shoal (Huo et al., 2016;Miao et al., 2020). The community structure of raft-attached green algae was reported signif icantly diff erent in 2017 from that in previous years for having a sharp decreased proportion ofU.prolifera(Xiao et al., 2020b). The changed macroalgae community on raft might aff ect the community structure of micropropagules in the environment of this region. On the other hand, variation in the community structure of green algae micro-propagules may further inf luence the species composition of attached algae on rafts,which deserves further study.
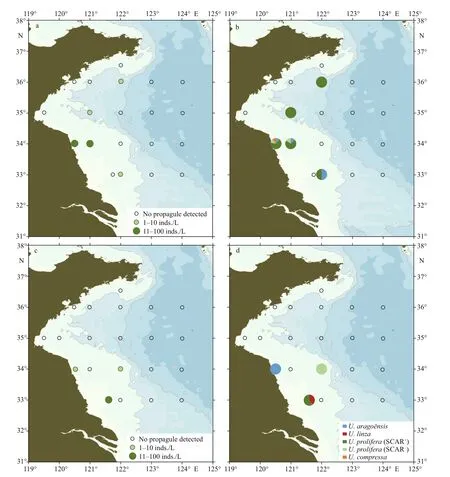
Fig.4 Distribution and community composition of green algae micro-propagules in the Yellow Sea in mid-June, 2019
It is interesting that theUlvaspecies identif ied from micro-propagules in Subei radial sand ridges were major fouling species on nori aquaculture rafts,too (Tian et al., 2011; Shen et al., 2012; Han et al.,2013; Huo et al., 2015; Li et al., 2015). Although Subei radial sand ridges are proximal to the coastline,common green algae ofUlvaspecies includingU.simplexandU.meridionalisalong the natural coast of Jiangsu Province (Xie et al., 2020) were not detected from the micro-propagules cultured in the present study. This phenomenon indicates that it is diffi cult for the transportation of micro-propagules along natural coastline into Subei radial sand ridges. On the other hand, abundantU.aragoënsismicro-propagules in June around the 33°N transect were rarely detected at nearshore sites in the 34°N transect, nor in the open sea area, indicating that the dispersing ability ofUlvamicro-propagules from Subei radial sand ridges to the open sea area is limited.
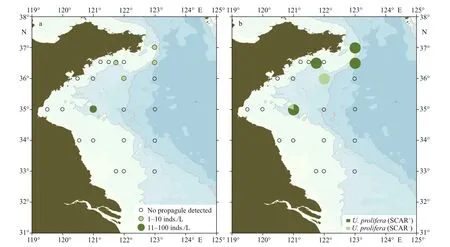
Fig.5 The density (a) and community composition (b) of green algae micro-propagules in water samples of the Yellow Sea in late July, 2019
Micro-propagules of f loating ecotype ofU.proliferawere proved to be present in Subei radial sand ridges.It was reported that the attached populations ofU.proliferaalong the natural coast of the Yellow Sea are genetically diff erent from that of the f loating ecotype that has not settled down in large scale (Zhao et al.,2015, 2018). Therefore, the natural coast could not be the source of the f loating seaweeds. At present, the attached adults of f loating ecotype with considering biomass are found from the nori aquaculture rafts in Subei radial sand ridges only (Zhang et al., 2018).Micro-propagules in the seawater and sediments are considered as the “seeds” of the fouling green algae on rafts (Liu et al., 2013; Huo et al., 2014; Wang et al.,2015). More studies will be conducted to understand whether the decreased quantity ofU.proliferamicropropagules in Subei radial sand ridges could aff ect the scale of the Yellow Sea green tide.
4.2 The green algae micro-propagules not likely exist in the open water of the Yellow Sea for a long time
The f loating green algae release micro-propagules into the environment during the northward drifting.The distribution ofU.proliferamicro-propagules was found closely related to the f loating algal mates in the open seawaters off the Subei Shoal in the Yellow Sea (Li et al., 2014; Miao et al., 2020). Results of the present study also support this viewpoint. In the cruise in June 2019, micro-propagules of three species,U.prolifera,U.aragoënsis, andU.compressawere detected from the seawater samples in Subei coastal area, while onlyU.proliferawas detected from the sites north of 35°N. In the late stage of the Yellow Sea green tide in July 2019, onlyU.proliferamicropropagules were detected from the seawater samples.The distribution pattern and species composition of the green algae micro-propagules corresponded to that of the f loating mats. For the sediment samples,micro-propagules were only present in the sites closed to the coastal area, but were absent from the open sea areas. Moreover, noUlvamicro-propagule was detected after the green tide in September from the seawater nor the sediment samples in any site in the open sea area of the Yellow Sea. Former studies proved that the duration of micro-propagules of the f loating ecotype in the seawater and sediments around Shandong Peninsula is very limited (Miao et al., 2018;Zhao et al., 2018). These results indicate that these areas are not suitable for the foating ecotype micropropagule preservation, and are impossible to be the“seed bank” of the green tide.
Additional to the Yellow Sea green tide, the coastal areas of Qinhuangdao in the western coast of the Bohai Sea, North China, is also an area of green tide reoccurrence from late April to late September since 2015. Cruise observations and molecular identif ications of f loating algae proved that the Bohai Sea green tides are originated locally, and the native attached macroalgae on seaweed beds are the major original source (Song et al., 2019a, b), which is similar to the most of green tides in relatively closed bays worldwide. Investigations on the distribution of green algae micro-propagules inshore and off shore of the Qinhuangdao coast show that the density of the micro-propagules in green-tide-aff ected areas is remarkable higher compared to other areas (Han et al., 2019).
Because of a huge algal biomass drifting for a long distance every year, it is generally believed that the formation and progression of the Yellow Sea green tides are quite diff erent from those of small-scale blooms with local origin (Keesing et al., 2011; Han et al., 2019). In this study, although someU.prolieramicro-propagules were found from the green-tideaff ected area during the blooming period, they were not persistently detected from the investigated open sea area. There are at least two reasons to explain the disappearance of micro-propagules.
(1) The micro-propagules might not be able to survive long time in the open sea area. However,many studies have proved that the micro-propagules ofU.proliferacould tolerate the harsh environmental conditions (Schories and Reise, 1993; Schories,1995; Liu et al., 2012). Fang et al. (2012) reported that even after one year of low-temperature and dark treatments, about 20% of green algal micropropagules germinated and developed into mature thalli. Therefore, it seems possible that some micropropagules remain vital in a short period. Another possible explanation is that micro-propagules might be eaten by some predators. In an intertidal zone,predator stress is insuffi cient to aff ect the germination of living micro-propagules and to maintain the local attachedUlvaspp. populations (Miao et al., 2018;Zhao et al., 2018). Although it is still unclear about the predator stress in the outer sea, the possibility that all of the micro-propagules are eaten in a short time seems very small.
(2) The micro-propagules might have been transferred to somewhere else by ocean currents.Seawater movement in the western Yellow Sea is complex, because of the intense interaction between the land and ocean. Water masses, continental shelf front, and the frontal region of the estuaries compose the unique current system in this sea area (Wei et al., 2011). The currents are considered to aff ect the drifting path of seaweeds (Keesing et al., 2011; Qi et al., 2017). Studies of the golden tide in the Yellow Sea show that the f loatingS.horneriin the sea area out of Shandong Peninsula could drift along with ocean currents to Subei radial sand ridges in autumn and winter (Xing et al., 2017). The same transportation might also occur on theU.proliferamicro-propagules in the study area. If such a case occurs, the micropropagules, left in the open waters of the Yellow Sea post green tide in summer, might return to Subei radial sand ridges via ocean f low and overwinter to seed the green tide outbreak in the next year. Future researches are needed to demonstrate whether this hypothesis is true.
5 CONCLUSION
In this study, a great number ofUlvamicropropagules in high density were detected in the Subei Shoal in April, 2019, which agrees with that of former studies. However, the proportion ofU.proliferawas signif icantly lower than that in the same season from 2009 to 2016, indicating that the quantity ofU.proliferamicro-propagules in 2019 was signif icantly decreased. During green tide outbreak in June and July, 2019,Ulvamicro-propagules were detected from the sites where the f loating seaweeds distributed, andU.proliferawas the dominate species.In September 2019 when the green tide ceased, noUlvamicro-propagule was detected in the open sea area of the Yellow Sea. Therefore, green algae micropropagules were unlikely to stay in the open sea water of the Yellow Sea for a long time. The mechanism of preserving the micro-propagules in Subei radial sand ridges, and the f inal destination ofU.proliferamicropropagules in the open sea area of the Yellow Sea after green tide deserves further study in the future.
6 DATA AVAILABILITY STATEMENT
The datasets analyzed during the current study were available from the corresponding author on reasonable request.
 Journal of Oceanology and Limnology2022年6期
Journal of Oceanology and Limnology2022年6期
- Journal of Oceanology and Limnology的其它文章
- Overview of harmful algal blooms (red tides) in Hong Kong during 1975–2021
- Information standardization for typical toxic and harmful algae in China’s coastal waters—a case study of Karenia mikimotoi*
- Biochemical composition of the brown tide causative species Aureococcus anophageff erens cultivated in diff erent nitrogen sources*
- Identif ication of paralytic shellf ish toxin-producing microalgae using machine learning and deep learning methods*
- Screening for lipophilic marine toxins and their potential producers in coastal waters of Weihai in autumn, 2020*
- First observation of domoic acid and its isomers in shellf ish samples from Shandong Province, China*