
珍稀植物银缕梅濒危机理及繁育保护研究
2022-12-27 05:26赖辅鑫阮向东台昌锐阳艳芳
安庆师范大学学报(自然科学版) 2022年4期
赖辅鑫,阮向东,赵 凯,台昌锐,阳艳芳,吴 彦*
(1.安庆师范大学 生命科学学院,安徽 安庆 246133;2.国家林业局和草原局调查规划设计院,北京 100714)
银缕梅(Parrotia subaequalis)为金缕梅科弗特吉族银缕梅属(Parrotia)下的落叶小乔木,是仅存我国并被发现的活化石树种,其在距今6 700万年前的中生代白垩纪化石中就已出现[1]。银缕梅于1999年被收录在我国《国家重点保护野生植物名录(第一批)》,其野生种群于2009年被国家林业局列入全国极小种群野生植物保护目录[2]。银缕梅作为最古老的植物物种之一,在金缕梅科的系统研究中具有很大的价值[3-4]。银缕梅树姿古朴,树干斑驳,于每年立春后开花,且花较叶先开放,夏天叶繁色绿,秋季叶面成黄、红等颜色,无论树形还是叶色都极具观赏价值,可用于盆栽制作[5]。将银缕梅引入城市绿地可以起到生物多样性保护和城市增色的作用,同时也能增加城市生态系统的服务功能[6]。由于自身繁衍困难、种群分布地碎片化等原因,目前银缕梅处于极度的濒危状态,亟待保护。本文从银缕梅的濒危原因,以及人工繁育和保护方面系统地总结了近三十年来的研究进展,为银缕梅的保护提供了科学依据。
1 银缕梅的地理分布
目前,全国已开展了银缕梅分布区的种群调查(表1),结果发现其为窄域分布的濒危保护物种,主要分布在我国华东地区[7]。银缕梅的生长生境大都降雨充沛(年平均降水量1 100 mm~1 400 mm),气候湿润(相对湿度75%以上),年平均气温在8.3°C~16.0°C,而海拔分布范围较宽,大多在400 m~700 m,少数可达1 000 m或以上,且生存土壤多为黄壤。目前该物种主要分布在江苏、浙江、安徽,同时河南、重庆等地也有发现。
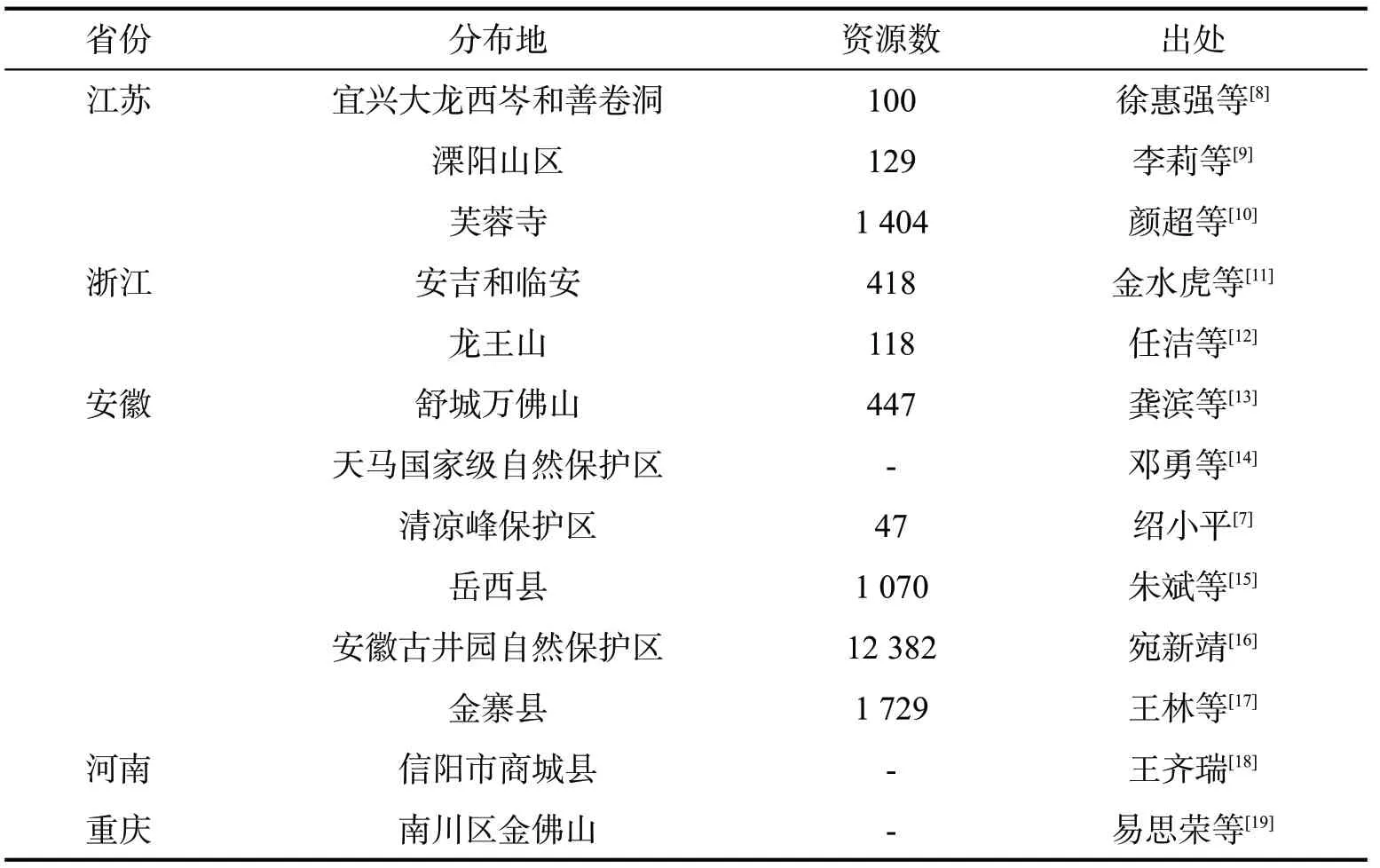
表1 银缕梅野外种群数据
2 银缕梅的濒危机理
关于银缕梅濒危的机理,学者主要从该物种自身的遗传水平,以及外部的环境因素等方面开展了探索研究,如表2所示。

表2 银缕梅的濒危原因

续表2
2.1 遗传水平
基因组的大小是植物生物多样性的特征参数之一。银缕梅基因组就属于极小基因组,同时又表现为高重复高杂合,加之居群数量较少,极易出现遗传漂变,这会在很大程度上影响物种的遗传多样性、基因表达和种群繁衍。银缕梅的居群遗传多样性可用平均期望杂合度(He)来衡量,该物种He总体较低,但稍高于其它濒危植物(He=0.42)。同时,从该植物的叶绿体基因组微卫星标记(cpSSR)来看,虽然其呈现高度片段化和局限性分布,但其单倍型多态性值与我国其它孑遗植物基本一致甚至高于它们。较高水平的多态性也意味着银缕梅各居群间存在一定程度的基因流限制。
2.2 环境胁迫
银缕梅与其伴生树种相比,对光适应的生态幅度较窄(表3),幼树与中树会受到大树遮蔽,导致叶片的有效光合辐射明显降低。在生长成为中树的过程中,银缕梅幼苗的光合生理特性和叶片形态均表现出对遮蔽环境的不适应,导致种群的中等径级个体急剧减少。作为阳生性树种,由于受到其他树木的遮挡,银缕梅幼苗生长后期的光资源不足会导致其植株处于“光饥饿”状态,从而制约了银缕梅对光的利用,最终导致了该物种的濒危。

表3 银缕梅及其伴生植物的光饱和点和光补偿点
土壤含水量也会影响银缕梅的生长。当土壤相对含水量为60%时,银缕梅的光合速率最快,且光饱和点最高。而在含水量60%以下时,随着土壤含水量的降低,该物种的平均光合速率、平均蒸腾速率、平均水分利用效率和平均气孔导度显著降低。与此同时,银缕梅的叶面积亦随着土壤含水量的增加而显著减少。
2.3 授粉影响
银缕梅存在几年一开花的现象,这很可能与严重的早春寒灾害有关。当银缕梅在花芽萌动期遭遇强寒流,会影响花芽生长,且花期的间隔性会导致结实机会变低。银缕梅无花瓣,雌雄花异熟,当花丝下垂,传粉受精将受阻。同时,银缕梅多生长在孤立的高山峡谷之中,地理隔离在一定程度上阻碍了风媒传粉。花粉流与种子流比值低容易导致该物种的近交和遗传变异减少,从而造成居群有效种群数量减少,最终产生不良效应。
2.4 种间竞争
银缕梅种群在争夺光照、水分、土壤养分等环境资源时不存在优势,特别是从更新层到演替层的发展阶段,较群落内其它阔叶树种的死亡率更高。例如,与青冈等阔叶树种在环境资源利用上相比,银缕梅在群落中仅处于从属和次要地位。银缕梅在不同生境下的种群数量,种内、种间竞争强度均有所不同。银缕梅在沟谷生境下比山坡生境下的种间竞争强度更小。银缕梅植株大小也会影响竞争强度,当胸径小于16 cm时竞争强度达到最大。种间竞争力不强是制约银缕梅生存和生长的一个主要因素,其将导致濒危现象的产生。
3 银缕梅繁育和保护
3.1 人工繁育
许多学者利用各种处理因素和条件对银缕梅的有性繁殖或无性繁殖开展了系列研究,结果发现各种处理下银缕梅的繁育效果均不相同(表4)。目前,银缕梅有性繁殖研究多是通过播种的方式开展,胡国伟等采用的方法能最大提高发芽率至73.73%;而在无性繁殖研究方面,主要是通过扦插的方式进行。扦插作为无性繁殖一种重要途径,可大量繁殖、快速成型。同时,银缕梅扦插实验主要围绕激素处理开展,可以说银缕梅在无性繁殖上有着较高的可行性。例如,黄绍辉等介绍的处理方法使得银缕梅扦插成活率达到88%以上,同时显著提高了苗的生长、生物量积累,以及生根数量。

表4 银缕梅人工繁育
3.2 保护措施
就地保护和迁地保护对于目前濒危物种而言是十分有效的保护方式。就现有的保护区来说,已实施了银缕梅种质资源库建设计划[39],主要采用就地保护的方式进行物种保护(表5)。然而,已确定的种质资源库虽是银缕梅种群较为集中的区域,但其在整个森林生态中分布不均、密度不大,且有着缩小的风险,进行物种补植将更有利于种群的恢复。同时,可将银缕梅引种到一些植物园中进行长期有效地繁育,也可逐步生长为成树[40](表6)。为保护银缕梅野生种群,提高其数量和质量,自然保护区多建立了专门的保护机构,培育专业技术人员并聘用护林人员开展了长期有效的保护工作。

表5 银缕梅种质资源保护方式

表6 南京盲人植物园引种的银缕梅幼树生长状况
4 展望
目前,还可以利用随机扩增多态性DNA技术、限制性片段长度多态性技术、扩增片段长度多态性技术等来研究银缕梅遗传多样性以揭示该物种的濒危机理。同时,可利用生态位模型来判断银缕梅的生态需求,并预测其实际分布和潜在分布。
针对银缕梅繁育,目前主要开展了播种和扦插两种人工繁育研究,同时,也可进行其他的人工繁育研究。例如无性系选育,选择自然界存在的优良基因型或人工杂交选育的优异基因型,经过无性繁殖、测定和筛选等过程,培养遗传性和表现型一致的优良群体。此外,也可以利用组织培养技术,利用银缕梅的一小部分组织或器官来培养无菌的优良苗木,该技术既不损伤原物种又可获得较高的增殖数量,可以更好地保护现有种群数量。最后,可开展银缕梅野外回归试验,选择合适的繁殖体、繁殖方式、回归地点和生境,优化定植时间和方式,开展回归后的长期监测。
在银缕梅保护方面,应加大原生境保护,在现有银缕梅种群及其他阔叶树种群间营造交错生境,根据银缕梅的生态习性有计划地减少其他物种对其生长的不利影响。同时,采取疏伐群落的方式,促进银缕梅种子、幼苗、幼树的生长。而且,需要对野生银缕梅开展封禁保护,防止人为的偷盗挖掘,保持种群数量和生态完整性,以期达到恢复和保护银缕梅野外种群的目的。
猜你喜欢
今日农业(2022年15期)2022-09-20
世界科学技术-中医药现代化(2022年3期)2022-08-22
落叶果树(2021年6期)2021-02-12
红土地(2018年7期)2018-09-26
西南农业学报(2016年6期)2016-04-16
体育科技(2016年2期)2016-02-28
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
河南医学研究(2014年7期)2014-02-27
