
胶孢炭疽菌C6型转录因子Cgctf1的功能分析
2022-12-18 03:24刘沙玉王地广柳志强
生物学杂志 2022年6期
曹 健, 刘沙玉, 王地广, 柳志强
(海南大学 生命科学学院, 海口 570228)
橡胶树是一种高大的乔木树种,发源于亚马逊热带森林,是天然橡胶重要来源[1]。橡胶炭疽病是橡胶树一种重要的叶部病害,可直接导致橡胶产量锐减[2]。橡胶炭疽病主要是由胶孢炭疽菌(Colletotrichumgloeosporioids)所引发的,常引起叶片枯黄、掉叶和嫩梢回枯等症状,造成严重的经济损失。橡胶炭疽病在全球范围内的感染面积每年都在扩大,造成橡胶产量急剧下滑[3]。目前,国内外对胶孢炭疽菌的研究主要针对生物学特性和化学防治等方面,有关该病菌致病基因功能,以及菌体与宿主之间的相互作用机制等方面的报道很少,极大地限制了相关防治技术的发展。
转录因子又称为反式作用因子,能够与顺式作用元件互作,确保目标基因在特定的时间和空间上准确表达[4]。锌指蛋白家族是最广泛地存在于原核及真核生物体内的转录因子家族,其结构域通过Cys和His的残基再加上处于游离态的锌离子共同结合而成,在基因表达的调控中起着重要作用[5]。根据Cys和His残基在数量大小和空间位置分布的差异,可将其锌指蛋白分为C2H2型、C4型和C6型3种[6]。对真菌而言,C6型锌指蛋白是一类特异的转录因子,通常会含有GAL4结构域,参与调控真菌生长、发育及致病等多个方面。如在酿酒酵母中,钙调神经磷酸酶通过调节C6型转录因子Crz1p影响钙依赖性基因表达的变化[7];在球孢白僵菌中,2个C6型转录因子Bbstr1和Bbstr2与氮源的利用相关,同时参与氧化应激反应和分生孢子的生长[8];在小麦镰刀菌中,EBR1的缺失造成孢子的萌发速度变快、萌发率降低,产孢量大量减少与致病力减弱[9];在粗糙脉孢菌中,转录因子vad-5对孢子发育有促进作用[10];在稻瘟病菌中,转录因子MoCRZI参与钙离子调节、分生孢子发育、细胞壁完整性及致病性[11];在蝗绿僵菌中,转录因子MaRcp缺失后该菌在正常培养条件下产孢方式发生了转换,即由正常产孢转换为微循环产孢[12]。
在前期研究中,我们从胶孢炭疽菌中鉴定了1个C6型转录因子Cgctf1,目前还未有关该转录因子生物学功能的研究。研究利用同源重组的方法对该转录因子基因进行敲除,通过表型分析探究其生物学功能,为深入认识胶孢炭疽菌致病的分子机理奠定基础。
1 材料与方法
1.1 材料
1.1.1 供试菌株
菌株:胶孢炭疽菌野生型菌株WT、大肠杆菌感受态DH5α;质粒:pMD18-T、pCB1532和pUC18载体;限制性内切酶、连接酶及相关试剂盒等购于宝日医公司。
1.1.2 供试培养基
CM培养基,PDA培养基,MM培养基,Czapek培养基[13]。
1.2 方法
1.2.1 基因克隆及序列分析
采用CTAB法获得胶孢炭疽菌基因组[14]。采用试剂盒提取菌株WT中的总RNA并合成cDNA。参照GenBank中胶孢炭疽菌基因组(参考基因ID:CGGC5_v011271)设计引物Cgctf1F和Cgctf1R(表1)。以cDNA为模板进行PCR扩增,PCR产物与pMD18-T载体连接,送测序公司进行测序。

表1 引物名称及序列
Cgctf1的蛋白结构域利用SMART(http:∥smart.embl.de/)进行在线分析;利用NCBI中BLASTp工具对Cgctf1蛋白进行序列比对,运用MEGA7.0软件中的邻近相接法构建系统进化树。
1.2.2 基因敲除与验证
基因敲除采用同源重组的方法,原理见图1。利用引物Cgctf1upF/Cgctf1upR和Cgctf1downF/Cgctf1downR(表1)分别对Cgctf1基因的上、下游序列进行扩增,连接到pCB1532上,得到敲除载体pKno-Cgctf1。利用PstI酶切对载体进行线性化,然后转入WT制备的原生质体中。原生质体制备及转化过程参照文献[15]。利用添加氯嘧磺隆的培养基进行筛选,提取转化子的基因组DNA,利用引物(Cgctf1F+Cgctf1R、Cgctf1UU+PI、Cgctf1DD+PI1)进行PCR验证。经电泳检测,Cgctf1F+CgctfR不能扩增得到ORF框,而Cgctf1UU+PI、Cgctf1DD+PI1均能扩增出片段的转化子,即为敲除突变体。

图1 Cgctf1基因的敲除原理Figure 1 Gene knockout mechanism of Cgctf1
利用引物Cgctf1hbF和Cgctf1hbR扩增Cgctf1基因的互补片段,与含潮霉素抗性基因的质粒pUC18相连接,构建互补载体pComp-Cgctf1。将互补载体转化到以△Cgctf1制备的原生质体中,在含有潮霉素的培养基中筛选抗性转化子,利用引物Cgctf1F+Cgctf1R进行PCR验证。能够扩增出Cgctf1完整的ORF框的抗性转化子,即为互补株。
1.2.3 敲除突变体表型分析
(1)营养生长及胁迫因子敏感性分析。将野生型、突变体和互补株接种于CM、MM、PDA和Czapek的平板上,28 ℃培养7 d,观察各菌株的生长情况。将上述菌株接种到添加有H2O2(20 mmol/L)、NaCl(0.8 mol/L)、SDS(0.01%)、KCl(1 mol/L)的MM平板上,28 ℃培养7 d,以接种MM培养基的菌株作为对照,观察并记录菌落直径,试验设置3个重复,并计算抑制率:
抑制率=(对照组菌落直径-实验组菌落直径)/
(对照组菌落直径-5 mm)×100%。
(2)孢子大小、孢子萌发率、产孢量和附着胞形成率测定。菌株活化后,接种于PDB培养基中,28 ℃,150 r/min振荡培养2 d,取1 mL滤液7 000 r/min离心8 min,弃上清液,添加1 mL无菌水重悬孢子,重复上述操作3次得到孢子悬液,利用血球计数板统计分生孢子的数量。将悬浮液的浓度调整为5×105mL-1,取25 μL分生孢子悬液(5.0×105mL-1)滴在载玻片上,28 ℃培养,分别于4、8和12 h观察分生孢子萌发及附着胞形成情况。每组处理设置3个重复。
(3)洋葱表皮侵染实验。取10 μL孢子悬液(2.0×103mL-1)滴加在洋葱内表皮上,28 ℃保湿培养,于8、12 h观察附着胞侵入和生长情况,每组处理设3个重复。
(4)玻璃纸实验。取20 μL孢子悬液(5.0×105mL-1)滴于已经铺有玻璃纸的PDA培养基上,28 ℃培养2、3 d后揭开玻璃纸,再培养1 d后观察结果,每组处理设3个重复。
(5)致病性分析。选取无伤的橡胶叶片,用酒精擦拭消毒。无菌针头微创2 mm伤口,采用菌饼及分生孢子悬液(1.0×106mL-1)两种方式,分别接种在橡胶叶片伤口位置,对照组(CK)分别为PDA琼脂块(菌饼方式)和无菌水(孢子悬液方式)。将叶片置于保鲜盒内28 ℃保湿培养,5 d后观察橡胶叶片的发病情况,每组试验设置3个重复。
2 结果与分析
2.1 Cgctf1基因生物信息学分析
利用PCR获得Cgctf1基因(GenBank登录号:MZ759779)的ORF序列,该基因共编码405个氨基酸,含有1个GAL4域 [图2(a)],属于C6型锌指蛋白。利用MEGA7.0以邻近相接法构建Cgctf1系统进化树 [图2(b)]。Cgctf1蛋白与热带炭疽菌(C.tropicale)的同源蛋白相似度在99%以上,与大丽轮枝菌同源蛋白的相似度为62.96%。与稻瘟病菌和酿酒酵母同源蛋白相似度仅为44.44%和44.90%。

(a)Cgctf1蛋白结构域分析,GAL4代表GAL4结构域;(b)Cgctf1系统进化树,标尺表示进化距离。图2 Cgctf1蛋白结构域及系统进化分析Figure 2 Protein domain and phylogenetic analysis of Cgctf1
2.2 Cgctf1基因敲除与验证
将敲除载体pKno-Cgctf1线性化后导入WT的原生质体中。经初步筛选和验证,发现转化子10号和43号能够在Cgctf1UU+PI、Cgctf1DD+PI1为引物时扩增出目标片段,而Cgctf1F+Cgctf1R为引物时,无法扩增出目的片段(图3)。最终确定第10和43号转化子为敲除突变体,命名为△Cgctf1-10和△Cgctf1-43。

(a)引物Cgctf1F+Cgctf1R上的扩增结果;(b)引物Cgctf1UU+PI上的扩增结果;(c)引物Cgctf1DD+PI1上的扩增结果。M:Marker 2000 bp; 1:WT; 2:△Cgctf1-10; 3:△Cgctf1-43; 4:△Cgctf1/ctf1。图3 Cgctf1基因敲除及互补验证Figure 3 Cgctf1 gene knockout and complementary verification
将互补载体pComp-Cgctf1转化到△Cgctf1- 43制备的原生质体中,利用潮霉素做抗性标记,初步筛选得到30个转化子,经过验证发现8号转化子可以扩增出Cgctf1基因的目的片段,确定为互补转化子,并命名为△Cgctf1/ctf1。
2.3 营养生长及胁迫因子敏感性分析
将野生型、突变体和互补株接种在PDA、MM、Czapek以及CM平板上,在28 ℃培养7 d,随后观察其生长状况。△Cgctf1-10和△Cgctf1-43在4种培养基上的菌落形态、菌丝生长速率同野生型相比无明显差异。将各菌株接种到含不同胁迫因子的MM培养基上,28 ℃培养7 d,结果发现敲除突变体对NaCl和KCl的耐受能力减弱,抑制率略高于野生型;相较于野生型,△Cgctf1对SDS和H2O2的抗性有一定程度增强,抑制率低于野生型(图4)。

(a)菌株在不同胁迫因子条件下的抑制程度比较;(b)生长抑制率显著性分析统计(差异显著水平P<0.05)。图4 胁迫因子对野生型、突变体和互补株生长的影响Figure 4 Influence of stress factors on wild type,mutant,complementary strains
2.4 Cgctf1参与调控分生孢子产生、萌发及附着胞形成
利用血球计数板统计各菌株的分生孢子产量,发现△Cgctf1的孢子产量明显低于WT[图5(a)]。野生型的孢子产量约为△Cgctf1孢子产量的3.65倍,说明Cgctf1参与调控分生孢子的产生。野生型分生孢子呈短棒状,大部分芽管较短,而△Cgctf1的分生孢子形态相对较大,接种4 h后,孢子萌发率为41%,附着胞形成率约为萌发率的一半,12 h后,孢子萌发率达到73%,附着胞形成率为41%。野生型在4 h时孢子萌发率就已经达到了90%水平,附着胞形成率也在85%以上 [图5(b)~(d)]。由此可见,Cgctf1与分生孢子萌发以及附着胞的形成有关。

(a)分生孢子产量统计;(b)孢子萌发率统计;(c)附着胞形成率统计;(d)分生孢子萌发及附着胞形成。co:分生孢子;gt:芽管;ap:附着胞。标尺:20 μm;差异显著水平P<0.05。图5 分生孢子产量,萌发及附着胞的形成Figure 5 Conidia production,germination and appressoriumformation
2.5 Cgctf1参与调控分生孢子的侵入能力
将各菌株的分生孢子悬液接种到洋葱表皮上,8 h后,野生型已经形成了明显的附着胞并且分化出侵染结构,12 h后侵染菌丝进一步扩大;突变株△Cgctf1分生孢子在8 h后萌发出芽管和附着胞,12 h后附着胞开始分化出侵染结构。相比WT,△Cgctf1在形成侵染结构时间上有所滞后(图6)。

co:分生孢子;gt:芽管;ap:附着胞;ih:侵染菌丝。标尺:20 μm。图6 洋葱表皮侵染实验Figure 6 Infection tests on onion epidermis
进一步测试分生孢子在玻璃纸上的穿透能力,突变株△Cgctf1在培养2 d时不能穿透玻璃纸,而野生型菌株已经穿透玻璃纸并形成菌落;培养3 d时,△Cgctf1能够穿透玻璃纸,但形成的菌落明显小于野生型。可以看出,Cgctf1参与调控分生孢子的侵入能力(图7)。

(a)菌株穿透玻璃纸形成的菌落大小比较;(b)菌落直径统计(差异显著水平P<0.05)。图7 玻璃纸穿透实验Figure 7 Cellophane penetration tests
2.6 致病性分析
将野生型、△Cgctf1与△Cgctf1/ctf1的菌饼和分生孢子悬液分别接种到离体橡胶叶片上,培养5 d后观察致病效果(图8)。无论是接种菌饼还是孢子悬液,△Cgctf1所形成的病斑直径均小于野生型菌株,致病力下降。因此,Cgctf1参与调控胶孢炭疽菌的致病性。
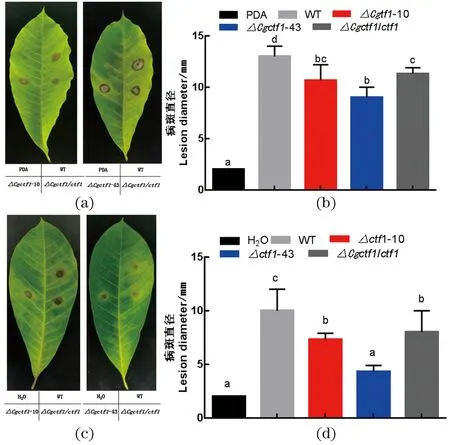
(a)菌饼接种橡胶叶片试验;(b)菌饼接种试验病斑大小统计;(c)孢子悬液接种橡胶叶片试验;(d)孢子悬液接种试验病斑大小统计。差异显著水平P <0.05。图8 致病性试验Figure 8 Pathogenicity tests
3 讨论与结论
研究从胶孢炭疽菌中鉴定了1个C6型转录因子Cgctf1,系统进化分析发现该转录因子的同源蛋白在其他真菌中普遍存在,但其生物学功能在其他病原真菌中还未有报道。研究成功获得了Cgctf1基因的敲除突变体,表型分析发现,Cgctf1的缺失并不影响胶孢炭疽菌的营养生长,但在NaCl、KCl、SDS、H2O2等胁迫因子的影响下,突变体生长与野生型相比表现出一定的差异,说明Cgctf1的敲除在一定程度上影响该病菌对高渗胁迫的响应及氧化应激反应。Cgctf1参与调控胶孢炭疽菌分生孢子的产生,其敲除突变体的分生孢子产量明显低于野生型;另外,从孢子萌发率及附着胞形成率来看,突变体分生孢子的活力与野生型相比明显降低。Cgctf1的敲除也造成了该病菌致病力的减弱,我们推测突变体分生孢子活力及侵入能力的减弱,可能是造成其致病力下降的主要原因。
目前有关胶孢炭疽菌转录因子功能的研究相对较少,已报道的转录因子如下:C2H2型转录因子CgAzf1参与调控胶孢炭疽菌分生孢子产生、萌发、附着胞的形成、黑色素合成以及致病性等[16];Cgglt1参与调控附着胞形成,且与黑色素分泌相关[17];bHLH型转录因子CgbHLH6与该病菌产孢能力、分生孢子形态、菌丝生长和抗氧化能力有关[18];bZIP型转录因子CgHapX是调控生物体内铁离子代谢的关键因子,该基因的缺失直接导致菌丝生长速率、产孢量、孢子萌发率及附着胞形成率显著下降[19]。在其他真菌中,C6型转录因子在参与调控生长发育及致病过程中发挥着不同的作用。在烟曲霉中,C6型转录因子LeuB参与调控亮氨酸生物合成和铁的获取[20];在大丽轮枝菌中,VdFTF1 参与调控细胞壁降解酶相关基因的表达,进而影响其致病性[21];在稻瘟病菌中,C6型转录因子PIG1参与黑色素的生物合成[22];芸薹生链格孢菌中,C6型转录因子AB04837参与调控营养生长,分生孢子发育,胁迫响应及致病性等[23]。由此可见,C6型转录因子的功能并不保守,此类转录因子在进化过程中功能不断分化,参与调控不同的生长发育过程。
综上所述,Cgctf1是胶孢炭疽菌一个新的C6型转录因子,其敲除突变体表现为对盐胁迫较为敏感,对H2O2和SDS的抗性增强,分生孢子产量减少,孢子萌发率和附着胞形成率明显降低,致病力减弱。因此,Cgctf1参与调控胶孢炭疽菌分生孢子产生及萌发,附着胞形成及侵入,胁迫响应及致病性等。
猜你喜欢
疯狂英语·新策略(2021年9期)2021-11-02
农业科技通讯(2021年1期)2021-03-06
热带作物学报(2019年4期)2019-06-11
武警医学(2018年10期)2018-11-06
中国森林病虫(2018年4期)2018-09-19
中成药(2017年9期)2017-12-19
上海农业学报(2017年3期)2017-04-10
天津医科大学学报(2015年2期)2015-12-22
塔里木大学学报(2015年1期)2015-04-25
中国当代医药(2015年20期)2015-03-01
