
大豆种子蛋白和油脂含量调控的研究进展
2022-12-17 07:11:52刘虹洁王金星刘昭军刘书侯兴亮李晓明
热带亚热带植物学报 2022年6期
刘虹洁,王金星,刘昭军,刘书,侯兴亮,李晓明*
(1.中国科学院华南植物园,华南农业植物分子分析与遗传改良重点实验室,广东省应用植物学重点实验室,广州 510650;2.中国科学院大学,北京 100049;3.黑龙江省农业科学院,哈尔滨 150086)
栽培大豆(Glycine max)起源于中国黄淮海地区,由野生大豆(G.soja)驯化而来[1]。与水稻(Oryza sativa)、小麦(Triticum aestivum)等粮食作物一样,大豆以种子为主要食用对象。因其含有丰富的蛋白质(约40%)和油脂(约20%)[2–3],大豆已成为人类食品和动物饲料中不可替代的植物蛋白和重要食用油来源。目前,全球近50%的植物蛋白质和植物油脂来源于大豆(www.soystats.com)。然而,面对全球快速增长的人口问题及养殖业的快速发展,选育高产优质的大豆品种已成为育种家们的首要目标[4–5]。
大豆种子蛋白和油脂含量是多基因控制的数量性状,同时还受环境因素的影响。多因素和多种调控途径共同决定了大豆种子的蛋白和油脂含量,虽然目前有一些研究报道了调控大豆蛋白油脂含量的基因位点,但关键基因的功能鉴定和调控网络还不清晰。此外,蛋白质和油脂含量间呈现此消彼长的关系[2],这也成为限制大豆蛋白和油脂含量共同提高的主要因素。解析大豆种子蛋白和油脂含量形成的分子机理,揭示两者之间的协同调控网络,是创制高产优质大豆品种的重要理论基础[6]。近年来,随着大豆基因组学和现代分子生物学研究的不断深入,利用基因定向改良等手段,精确调控大豆种子蛋白和油脂含量,以实现高产优质大豆品种的育种目标。本文针对大豆蛋白和油脂品质性状的分子调控机制进行综述,并讨论了目前大豆蛋白和油脂性状分子研究面临的挑战和前景,以期为大豆种子品质定向改良提供理论参考。
1 大豆种子贮藏物质的组成
种子发育是大豆生长发育的关键一环,决定了产量的高低和品质的优劣[7]。在大豆的生长发育过程中,叶片等“源”器官经过光合作用产生蔗糖并运输到种子中。种子发育初期首先是淀粉积累,到种子发育中后期,淀粉水解以提供大量能量和原料,合成蛋白质和油脂。成熟种子中淀粉含量占籽粒干重的1%~3%,此外,还含有26%~30%的碳水化合物。碳水化合物组成复杂,其中,包括棉子糖、水苏糖在内的大豆低聚糖占比最高。蛋白质和油脂是大豆种子的主要组成部分,分别约占大豆组成的40%和20%[8]。蛋白质和油脂含量也是大豆用途和品质的主要决定因素。然而,大豆种子中蛋白质含量和油脂含量呈负相关关系,二者之间存在底物竞争关系[2]。
2 大豆种子油脂含量的调控
2.1 油脂生物合成关键酶
大豆种子的油脂累积与多数植物种子相似,受到脂肪酸(FAs)生物合成、三酰基甘油(TAGs)组装以及油脂和其他细胞代谢物之间碳分配过程这3 方面的影响[9–10]。脂肪酸(FAs)在质体中合成,此过程受到乙酰辅酶A 羧化酶(ACCase)和3-酮脂酰-酰基载体蛋白合酶(KAS)等多种关键酶的调控。积累的FAs随后转移至内质网,经过G3P 酰基转移酶(GPAT)、溶血磷脂酸酰基转移酶(LPAAT)、磷酸酶(PAP)和二酰基甘油酰基转移酶(DGAT)的酰化和酯化(肯尼迪途径),完成TAGs 的组装[11]。其中,大豆中TAGs产生的主要途径是由磷脂酰胆碱(PC)转化为1,2-sn-二酰基甘油(DAG),DAG 再酰化产生TAGs[12]。在碳分配方面,丙酮酸脱氢酶激酶(PDHK)和胞质D-葡萄糖-6-磷酸脱氢酶(Glu6PDH)影响FAs 的合成进而影响种子油脂含量[13–14](图1)。
在种子胚胎发育后期,脂肪酶介导TAG 转变导致大豆种子脂肪酸含量下降,寡糖、棉子糖和水苏糖含量升高。研究表明,抑制脂肪酶介导TAG 的转变可以增加脂肪酸含量,减少不可消化的低聚糖的存在。大豆中的GmSDP1基因与拟南芥(Arabidopsis thaliana)脂肪酶基因同源,编码TAG 脂肪酶。Gm-SDP1 介导TAG 转换的同时影响脂肪酸的组成,抑制SDP1基因的表达不仅能够促进种子油分的积累,增加油酸含量,降低亚油酸的含量还能使蛋白质含量呈上升趋势[15–16]。
除了增加油脂的合成与累积,改变大豆中脂肪酸的组成比例,提升油脂利用率也是目前的育种策略之一。大豆的油脂以酰基甘油(TAGs)形式储存在种子中[11],具体可分为棕榈酸(16∶0)、硬脂酸(18∶0)、油酸(18∶1)、亚油酸(18∶2)和亚麻酸(18∶3)这5 种脂肪酸[3],其中,亚油酸和亚麻酸约占68%,属于多不饱和脂肪酸(PUFAs),易于氧化,过多的PUFAs会降低大豆油的氧化稳定性,影响大豆品质[3]。
2.2 转录因子
转录因子在种子油脂合成与累积过程中也发挥着重要作用。目前,大豆种子中已鉴定出多个油脂合成相关的重要转录因子,如GmDofs、GmMYB73、LECs(Leaf cotyledon)和WRI1 (Wrinkled1)等(图1)[17–22]。
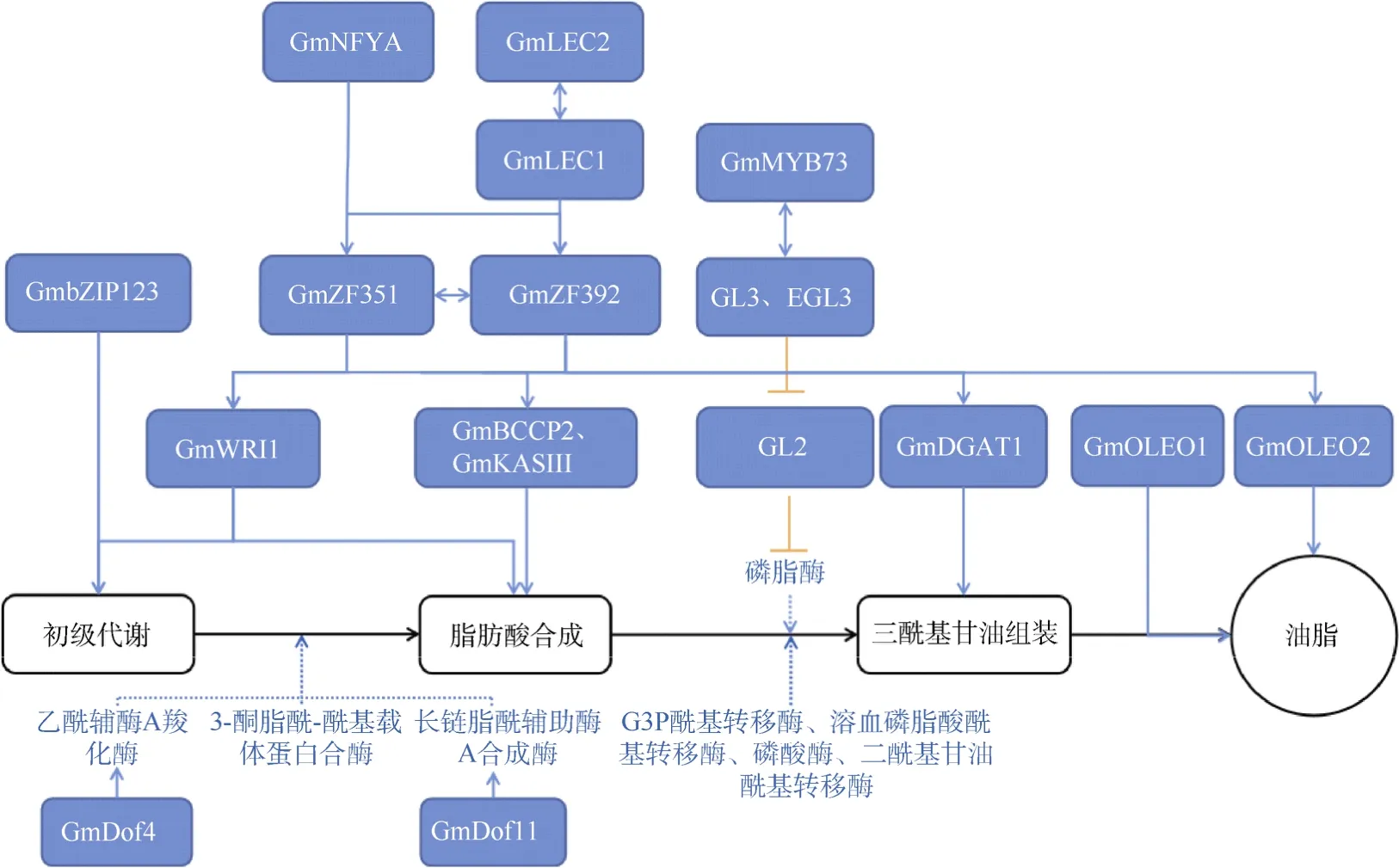
图1 大豆油脂合成的转录调控模块Fig.1 Transcriptional regulatory modules of soybean oil synthesis
GmDof4 (DNA binding with one finger 4)和GmDof11 是2 个油脂合成相关的转录因子,参与FAs 的合成调控。GmDof4 和GmDof11 分别激活了ACCase 和长链脂酰辅酶A 合成酶(LACS)基因的表达,提高转基因拟南芥种子中总脂肪酸和脂质的含量。Zhang 等[23]将GmDOf4基因转入椭圆小球藻,GmDof4能够显著促进其油分累积。此外,GmDof4和GmDof11能够直接结合储存蛋白基因CRA1的启动子,下调储存蛋白基因的表达[17]。
在大豆中特异表达的GmMYB73 属于MYB 家族的转录因子,参与TAGs 的合成调控。研究表明,GmMYB73 能够与bHLH 型转录因子GL3 (Glabra 3)和EGL3 (Enhancer of glabra 3)互作,抑制油分累积的负调控因子GL2 (Glabra 2)的表达,进而解除GL2对磷脂酶PLDα1 的抑制。解除抑制的PLDα1 促进PC 水解并最终合成DAG 和TAG[18]。此外,GqOil2上的候选基因GmOLEO1编码油球蛋白能够影响TAG 的代谢,促进油分的积累[24]。
GmLEC2a 是含有B3 结构域的转录因子。Gm-LEC2a基因位于第20 号染色体上,与拟南芥LEC2同源。拟南芥的GmLEC2a过表达株系的TAG 含量增加34%,长链脂肪酸组成增加4%。Manan 等[19]将GmLEC2a在大豆毛状根中异位表达,结果多个与油脂合成相关的转录因子表达上调,包括GmLEC1、GmFUS3、GmABI3 (ABA insentive3)、GmDof11 和GmWRI1。
WRI1 是编码AP2/乙烯响应元件结合蛋白(APE TALA2/EREBP)家族的一个转录因子。在GmWRI1的3 个同源基因GmWRI1a、GmWRI1b和GmWRI1c中,GmWRI1a主要在大豆种子中表达。GmWRI1a过表达大豆转基因株系种子的总油量和脂肪酸含量增加。此外,GmWRI1a能够与保守的AW-box顺式作用元件特异性相互作用,控制大豆油分相关生物合成基因的表达[25]。
转录因子GmbZIP123 可以直接结合蔗糖转运体基因SUC1、SUC5和细胞壁转化酶基因cwINV1、cwINV3和cwINV6的启动子,并促进这些基因的表达,控制糖从光自养组织进入种子,参与大豆种子中脂质积累的调控[26]。此外,Hu 等[27]鉴定了1 个种子驯化过程中的选择基因GmZF351,其编码串联CCCH 锌指蛋白。GmZF351能够激活脂质生物合成相关基因GmBCCP2(biotin carboxyl carrier protein 2)、GmKASIII、GmDGAT1(diacylglycerol-O-acyltransferase 1)和GmOLEO2(oleosin 2)的表达,增强WRI1 活性,正向调节脂质的生物合成[28]。锌指蛋白GmZF392 同样能够提高大豆种子的油脂含量,一方面GmZF392 能够与启动子区富含TG 和TA 的双元元件结合,激活脂质生物合成途径中基因的表达;另一方面,GmZF392 与GmZF351 互作,协同促进下游基因的表达。不仅如此,NF-Y 类的核因子GmNFYA 分别通过直接和间接调控促进GmZF392和GmZF351基因的表达,提高转基因大豆种子中的含油量[29](表1)。

表1 种子油脂含量的主要调控因子Table 1 Major regulatory factors that control seed oil content
3 大豆种子蛋白质含量的调控
大豆种子蛋白质根据功能可分为11 组,其中,含量最丰富的是种子储藏蛋白。储藏蛋白主要由球蛋白(11S)和β-半球蛋白(7S)组成[30–31],两者共同影响着大豆蛋白的含量与品质[32–34]。大豆种子球蛋白和β-半球蛋白分别为六亚基和三亚基聚合蛋白,分别由多基因Gy1-Gy7和CG1-CG15编码[35–36],其中,7S 上的β亚基编码基因CG4由1 个位于第20 号染色体上的显性位点qBSC-1控制,影响大豆蛋白的质量和加工性能[37]。相较于大豆种子油脂,对控制大豆种子蛋白的基因功能鉴定较少。Han 等[38]利用整合转录组学和蛋白质组学特征对1 个野生大豆品系的染色体片段替换系(CSSL)和亲本系进行分析,鉴定出27 个差异表达基因(DEGs)和23 个差异积累蛋白(DAPs)调控种子储藏蛋白。这些基因编码的蛋白参与光合作用、蛋白质加工、蛋白质分选和储存蛋白质积累过程(图2)。
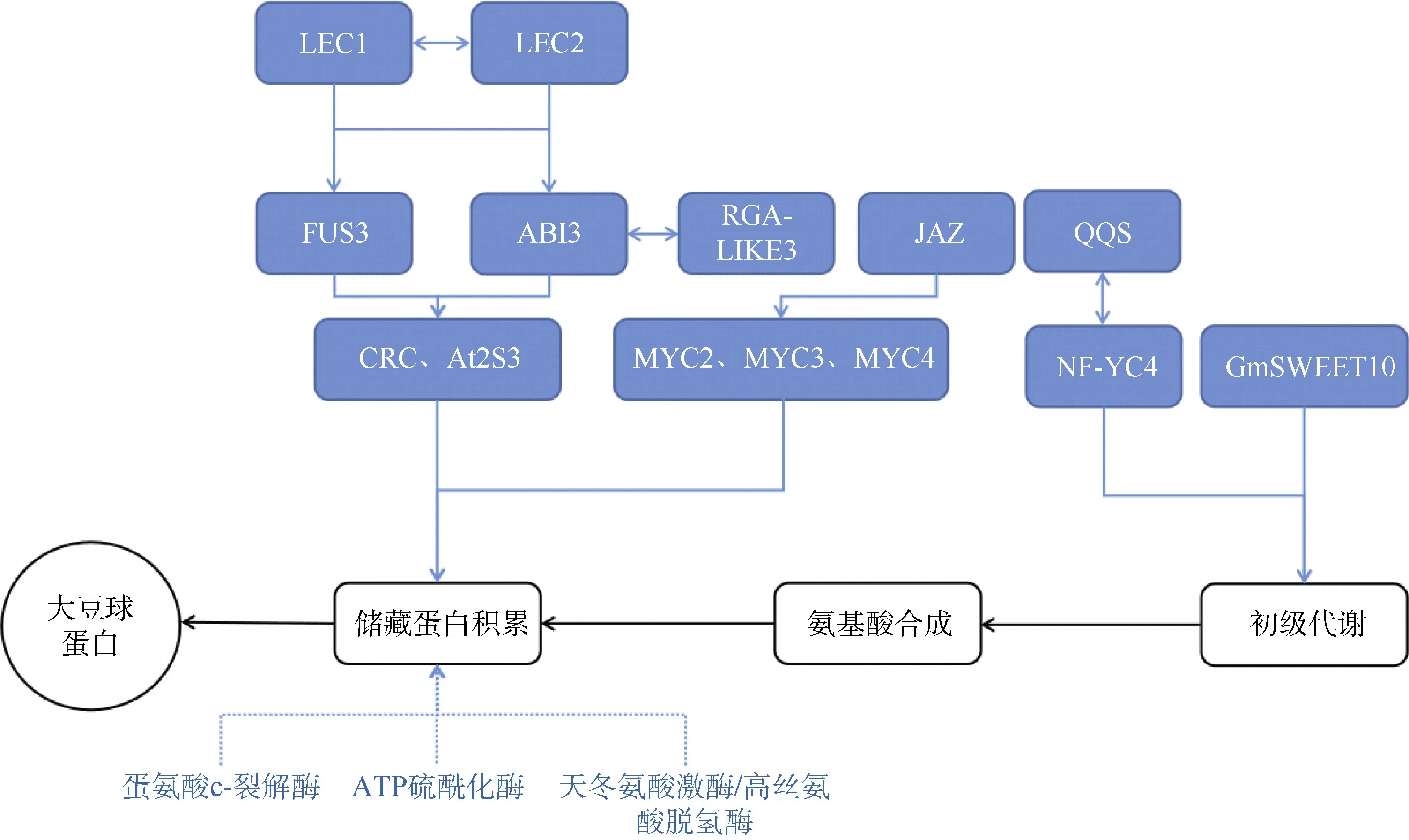
图2 大豆蛋白合成的转录调控模块Fig.2 Transcriptional regulatory module of soybean protein synthesis
种子蛋白质和油脂合成所需的原料主要来源于糖类物质的分解。糖的分配影响胚胎的发育也调节脂肪酸的生物合成和蛋白质的生物合成[39–40]。位于15 号染色体上的糖转运体基因Glyma.15G049200(GmSWEET10)调控着种子蛋白质和油脂的积累,该基因编码的蛋白定位于质膜,属于SWEET 家族成员[41]。此外,Glyma.15G049200基因存在等位基因GmSWEET10a和GmSWEET10b,两者功能冗余,主要表达于种皮的薄壁组织和表皮中[42],可能通过影响糖从母体种皮到子代胚的传递来调节油脂和蛋白质的积累[43]。
碳源和氮源的分配影响着氨基酸和脂肪酸的合成进而影响种子蛋白质和油脂的含量。Li 等[44]报道在拟南芥中特有的“孤儿基因”QQS能够与转录因子NF-YC4 相互作用调控碳和氮分配促进种子的蛋白质合成。此外,NF-YC4 在植物中广泛存在,其同源物能够与QQS 蛋白结合互作,QQS能够调控拟南芥、大豆等物种的碳和氮分配,从而影响种子蛋白质的合成。大豆储存蛋白最初在内质网上合成,然后遵循高尔基体介导的运输途径沉积至蛋白质储存液泡(PSV)。Wei 等[45]报道在大豆子叶发育过程中,小的GTPase 编码基因GmRab5a及其鸟嘌呤核苷酸交换因子(GEFs),参与调控大豆储存蛋白的运输,影响蛋白质的积累。
一些酶也调控着大豆蛋白质的组成和积累。研究表明,蛋氨酸c-裂解酶(MGL)调控大豆种子中甲基蛋氨酸的积累[46]。蛋氨酸和半胱氨酸分别为人体必须氨基酸和条件必需氨基酸,过表达的ATP 硫酰化酶(ATPS)能够促进大豆中这2 种氨基酸的合成,从而提升大豆蛋白品质[47]。磷酸烯醇丙酮酸羧激酶(PEPC)活性在高蛋白品种中高于低蛋白品种,可作为大豆育种中的一种生物标志物[48]。Zhang 等[49]在qOC-8-1(qWSPC-8-1)株系中鉴定出1 个对油脂和水溶性蛋白的合成起反作用的Glyma.08G107800基因,该基因的表达与大豆的油类合成呈负相关,与水溶性蛋白的合成呈正相关,Glyma.08G107800编码一种双功能酶:天冬氨酸激酶/高丝氨酸脱氢酶(AK-HSDH),该酶对天冬氨酸家族(赖氨酸、苏氨酸、甲硫氨酸和异亮氨酸)的氨基酸合成非常重要。
种子蛋白质的合成与积累除了由其合成代谢和组分构成等因素决定,还受到多种转录因子的共同调控。目前,对于控制大豆种子蛋白合成的转录因子鉴定较少,已报道的相关转录因子多集中于拟南芥中。拟南芥中鉴定的FUS3、LEC2 和ABI3(ABA insentive3)是植物特异的B3 类转录因子,能够以直接调控或间接调控的方式影响储藏蛋白基因At2S3的表达,其中,FUS3 和LEC2 存在部分功能冗余[50–51]。核因子LEC1 是植物种子发育的核心调控因子,参与种子发育的多个生物学进程。Kagaya等[52]研究表明,LEC1 通过控制FUS3 和ABI3 协同调控种子储藏蛋白基因CRC和At2S3的表达,从而影响种子储藏蛋白的积累。Chern 等[53]在菜豆(Phaseolus vulgaris)中鉴定出bZIP 类转录因子ROM1 是一种DNA 结合位点依赖的抑制因子。ROM1 与转录激活因子PvALF 拮抗,抑制发育过程中种子蛋白合成基因的转录。此外,转录因子与激素之间相互作用同样影响着种子储藏蛋白的积累。赤霉素信号负调控因子RGA-LIKE3 作为ABI3 的共激活因子,在种子成熟阶段促进储藏蛋白的生物合成[54];茉莉酸信号转导中的抑制蛋白JAZ 能够直接靶标bHLH 类转录因子MYC2、MYC3 和MYC4,协同调控种子储藏蛋白的积累[55]。近期研究表明,在第20 号染色体上存在1 个与大豆种子蛋白含量呈负相关的基因Glyma.20G85100,其编码的蛋白中CCT结构域存在1 个转座子的插入,从而影响该基因的表达,同时用RNAi 干扰抑制Glyma.20G85100基因,能够显著提升种子蛋白含量,具体机制仍有待研究[56](表2)。

表2 种子蛋白含量的主要调控因子Table 2 Major regulatory factors that control seed proteins
4 大豆种子蛋白、油脂的调控位点
大豆种子的油脂、蛋白含量是由多基因控制且易受环境影响的数量性状。Lee 等[57]利用限制性片段长度多态性(RFLP)标记对(“Young”×PI416937)和(PI97100דCoker237”)进行分析,在第15 (E 连锁群)、18 (G 连锁群)、12 (H 连锁群)和9 (K 连锁群)号染色体上存在多种蛋白和油脂相关的共同标记。Liang 等[58]利用群体(Jindou23×Huibuzhi)分别检测出6 个蛋白和11 个油脂数量性状座位(quantitative trait locus,QTL)。从2 个群体(Magellan×PI 438489B)和(Magellan×PI 567516C)分别获得216 和156 个重组自交系。Pathan 等[59]在大豆中分别确定了7 个蛋白和6 个油脂QTL,其中,第5 和6 号染色体上存在蛋白质和油脂共同的QTL。Li 等[60]则对长江、淮河流域的大豆种子进行多位点全基因组关联分析,鉴定出10 个新的与油脂或蛋白相关的QTLs,还分别鉴定出55 个与种子蛋白和51 个与油脂相关的候选基因。Zhang 等[49]对211 份大豆种质进行种子组成的全基因组关联研究(genome-wide association studies,GWAS)分析,分别鉴定出蛋白质(3 个)、脂肪(4 个)和水溶性蛋白质(5 个)相关的QTL,其中,qOC-8-1(qWSPC-8-1)对油脂和水溶性蛋白的合成起反比作用。GqOil20是1 个环境稳定的QTL,与油脂含量显著相关,占多种环境下种子油脂总表型变异量的23.70%[24]。
通过对不同群体进行QTL 和GWAS 分析,已报道的与大豆蛋白和油脂相关的QTLs 分别有248 和327 个,分布于20 条染色体上(http://www.soybase.org,2022-07) (表3)。大豆种子蛋白相关的QTLs 位于第6、9、18 和20 号染色体上较多,其中,第20号染色体上的蛋白相关的QTL 与GWAS QTL 数目均最多,最高可解释65%的表型变异[61]。大豆种子油脂相关的QTLs 位于第5、15、18、19 和20 号染色体上较多,其中,第15、18 和19 号染色体上的QTL 数目均为 23 个。
5 讨论和展望
大豆是世界粮食安全的重要组成部分,大豆蛋白优质[62],能够提供人体所不能合成的必需氨基酸,同时大豆油脂中饱和脂肪酸较少,胆固醇含量低,有益于人体健康[63–64]。长期以来,大豆已成为人类食品和饲料中不可替代的植物蛋白和食用油来源。面对世界人口激增、耕地面积短缺和多变的自然气候,培育出能够适应未来环境变化的优质大豆品种已成为迫切需要。提高大豆油脂和蛋白质含量更是一直以来的育种目标。
为培育高产优质的大豆品种,通常需要将多个优良品质性状聚合,但大豆油脂和蛋白质含量受多个数量位点的控制,且调控途径相互串扰,传统育种技术很难克服这个限制。有研究认为大豆蛋白与油脂之间存在一定的底物竞争,两者合成所需的原料均来源于糖类物质的分解[65–66]。打破蛋白与油脂间的负相关性以及维持多性状的平衡关系是实现大豆品质改良的关键。目前,油脂蛋白合成的调控研究主要为合成途径中关键酶[67–69]及一些转录因子[70–71],仍有大量的种子蛋白油脂含量的位点需要克隆和功能验证,从而进一步完善大豆蛋白和油脂基因的合成代谢网络。
现代栽培大豆由野生大豆驯化而来,长期的驯化历程使植物朝着更有益于人类社会生产的方向发展,但这也导致植物大量优异农艺性状丢失,遗传多样性降低。如何在避免其他优异农艺性状丢失的同时选择目标性状,有研究者认为从头驯化可能是另一种途径[72–74]。野生大豆具有高蛋白低油脂的性状,经过千年的驯化,如今的栽培大豆油脂含量高,蛋白含量低,抗逆性差。通过对野生大豆从头驯化或再驯化,针对性的选择高油脂和蛋白含量等位基因,清除有害变异,减少或避免其他优异基因的丢失,也可能会实现大豆高产优质的育种目标[75]。
现代分子生物学、基因组学的快速发展,促进了蛋白和油脂含量调控基因的挖掘和调控网络的解析。同时,随着基因定向编辑[76–77]、大豆快速遗传转化[78–79]等技术的进一步突破,也会加速大豆品质性状的精准改良,最终实现大豆产量和品质的共同提高。
猜你喜欢
食品工业(2023年12期)2023-12-12 12:56:16
今日农业(2022年16期)2022-11-09 23:18:44
中国化肥信息(2022年5期)2022-08-30 01:58:26
今日农业(2021年20期)2021-11-26 01:23:56
今日农业(2021年14期)2021-10-14 08:35:34
中国生殖健康(2020年5期)2021-01-18 03:00:06
疯狂英语·新读写(2020年3期)2020-06-06 09:06:14
食品与机械(2018年11期)2019-01-03 06:15:34
中国生殖健康(2018年5期)2018-11-06 07:15:56
中国粮油学报(2018年12期)2018-01-26 13:32:04
