
铜绿假单胞菌T3SS和T6SS的基因分布特性及与耐药的相关性
2022-12-16 01:14周蓓蓓王凌波张秀彩陈栎江王忠永周铁丽
温州医科大学学报 2022年12期
周蓓蓓,王凌波,张秀彩,陈栎江,王忠永,周铁丽
1.温州医科大学附属第一医院 医学检验中心 浙江省检验诊断及转化研究重点实验室,浙江 温州 325015;2.浙江大学医学院附属儿童医院 实验检验中心,浙江 杭州 310052
铜绿假单胞菌(Pseudomonas aeruginosa)是一种能引起多部位感染的人类常见机会致病菌,可以导致肺部、尿路、烧伤创面和糖尿病足等多种感染[1]。研究显示,由铜绿假单胞菌导致的感染在院内感染中呈不断上升的趋势,且多重耐药铜绿假单胞菌日益增加,限制了抗菌药物的治疗选择[2-3]。 铜绿假单胞菌具有多种促进感染发生发展的毒力因素,其中分泌系统是重要的毒力因素之一,细菌可以通过分泌系统获取营养成分,向外界环境分泌毒素蛋白,或者直接作用于真核生物的作用靶点发挥致病性[4]。较多研究表明[5-6],III型分泌系统(type III secretion system, T3SS)在临床上常与严重感染相关,铜绿假单胞菌可通过T3SS将分泌蛋白注入宿主细胞;VI型分泌系统(type VI secretion system, T6SS)是铜绿假单胞菌的另一分泌系统,可编码3个T6SS分泌系统(H1-、H2-和H3-T6SS)[7]。目前有较多国内外报道对临床菌株T3SS毒力因子的流行和作用进行探究,但T6SS毒力因子的流行情况及特性仍未完全明确,并且最近发现的T6SS效应蛋白PldA、PldB具有较强的毒力效应,不仅可以竞争杀菌还可内化至非吞噬细胞[8],因此对于T6SS相关因子携带率的检测和探究极为重要。另外BOULANT等[9]发现T3SS及T6SS相关因子在引起人类不同感染的铜绿假单胞菌中的携带具有一定的差异,而呼吸道和血液作为铜绿假单胞菌分离的两大标本来源,探究T3SS和T6SS效应基因与不同标本来源菌株之间的关系具有一定的临床意义。
细菌耐药性和毒力特征是影响铜绿假单胞菌感染临床预后的关键因素,随着多重耐药铜绿假单胞菌在临床中的快速出现,迫切需要新的方法来治疗由铜绿假单胞菌引起的各种严重感染。毒力抑制策略是一种有潜力的策略,而了解临床菌株中编码毒力因子基因的分布情况是关键。本研究旨在通过检测痰液及血液来源标本中T3SS和T6SS效应蛋白相关基因的分布特性,并分析其与细菌耐药性的关系,为临床控制铜绿假单胞菌所致感染以及药物靶向治疗提供理论依据。
1 材料和方法
1.1 菌株来源 收集2016年至2018年间分离自温州医科大学附属第一医院住院患者痰液和血液标本的铜绿假单胞菌共92株(剔除同一患者相同来源的重复菌株),其中来源于痰液标本61株,血液标本31株。菌株分离:将血液样本接种至血培养瓶中,血培养报阳后接种于血平板上,37 ℃培养18~24 h 后观察菌落形态;将痰液标本接种于血平板、嗜血平板及科玛嘉平板,37 ℃培养18~24 h后观察菌落形态;将疑似菌落由基质辅助激光解析/电离飞行时间质谱(MALDI-TOF MS)鉴定系统进行鉴定,确定为铜绿假单胞菌,菌株保存在30%甘油肉汤保菌管中,并存于-80 ℃超低温冰箱中。本研究已获得本院伦理委员会批准(伦理批号:KY2022-R139)。
1.2 仪器和试剂 VITEK MS质谱鉴定仪(法国生物梅里埃公司,VITEK MS PLUS);PCR扩增仪(美国Thermo公司,VertiTMDx 96 Well Thermal Cycler)。Muller-Hintom(MH)琼脂(英国OXOID有限公司);细菌基因组提取试剂盒(北京BioFlux公司);引物合成(上海华大生物科技有限公司);PCR反应试剂(大连Takara公司);凝胶成像分析系统(美国BIO-RAD公司)。
1.3 VITEK MS质谱鉴定仪菌株鉴定 将待测微生物与基质液进行混合加在靶板上,待干后形成样本基质结晶体,经激光辐射使样本的蛋白进入飞行时间检测器,检测器通过检测蛋白飞行时间的不同来建立微生物蛋白质量指纹图,进而与数据库中的信息比对来对微生物种或菌株进行区分和鉴定。大肠埃希菌ATCC 8739作为实验的校准菌株,鉴定可信度达到99.9%则证明鉴定正确。
1.4 聚合酶链反应(PCR) 参照细菌基因组提取试剂盒说明书方法提取基因组模板,使用特异性引物进行PCR扩增。PCR引物序列参照文献[6-7]合成,引物序列见表1。在PCR扩增仪中设置反应条件如下:预变性95 ℃ 10 min;变性95 ℃ 1 min;退火30 s(温度视基因而不同,见表1);延伸72 ℃ 1 min,35个循环;再延伸72 ℃ 5 min。PCR扩增产物经1%琼脂糖凝胶电泳,GelRed染色后于凝胶成像系统观察结果。所有PCR阳性扩增产物均送至上海华大基因科技有限公司进行测序确认。
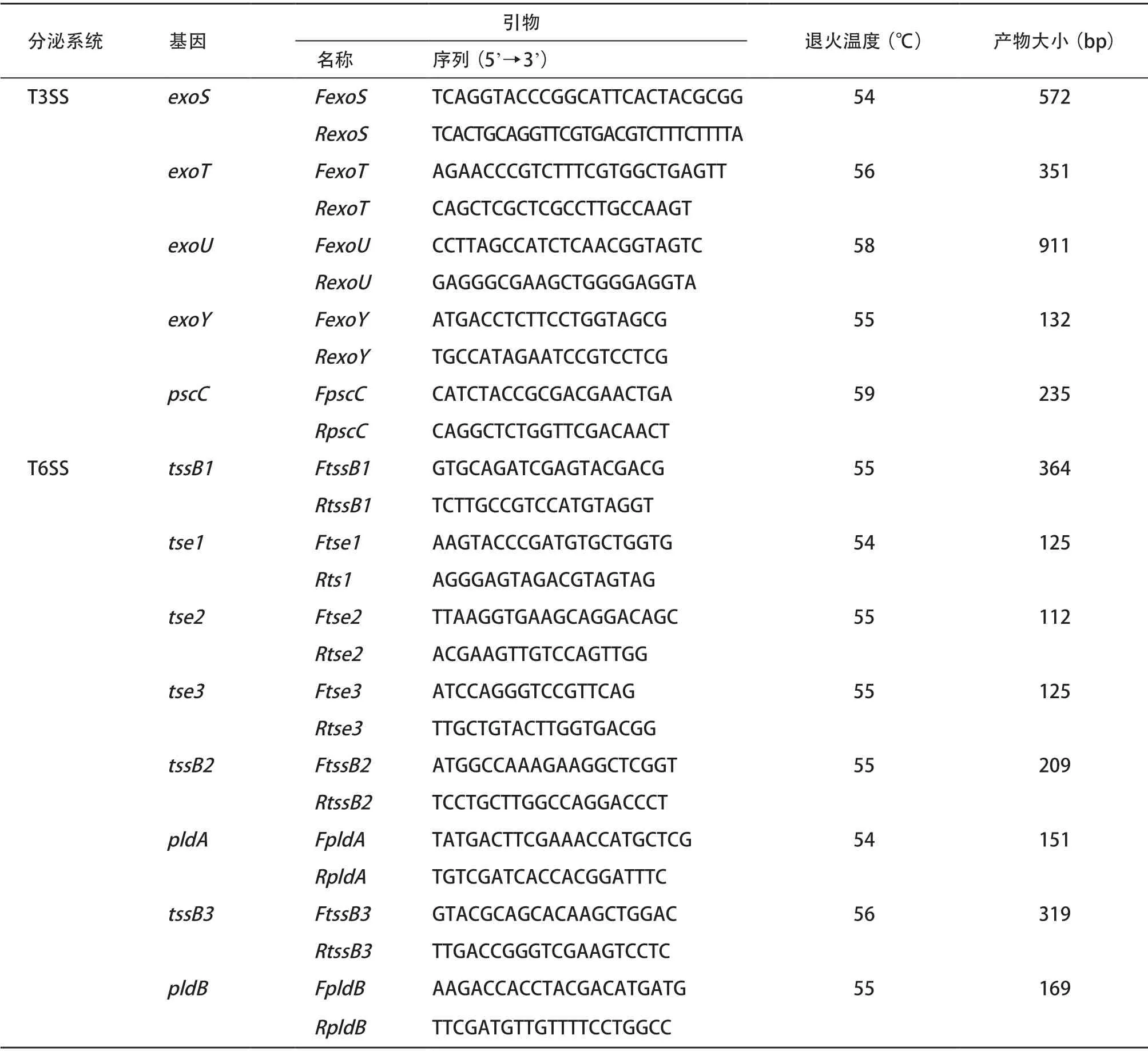
表1 T3SS和T6SS效应蛋白基因引物序列
1.5 药物敏感性试验 采用琼脂稀释法检测菌株对常用抗菌药物亚胺培南、环丙沙星、头孢他啶、阿米卡星的敏感性,结果判读参照美国临床和实验室标准化协会(Clinical and Labortory Standards Institute, CLSI)2020年标准判断[10]。
1.6 统计学处理方法 采用SPSS26.0统计软件进行处理,计数资料以频数和率表示,组间比较采用χ2检验或Fisher确切概率法,利用Spearman相关系数分析部分菌株中T3SS和T6SS效应蛋白编码基因之间的相关性。P<0.05为差异有统计学意义。
2 结果
2.1 不同来源T3SS和T6SS效应蛋白基因检测结果 92株临床分离铜绿假单胞菌中T3SS效应蛋白编码基因exoT及T6SS效应蛋白编码基因tssB1、tse2、tssB2、pldB均高达90%以上;其中痰液标本来源菌株T3SS效应蛋白基因exoU的携带率显著高于血液来源菌株(P<0.05),而T3SS及T6SS其他相关效应蛋白基因的携带率在两种不同的标本来源的菌株中差异无统计学意义(P>0.05),见表2。
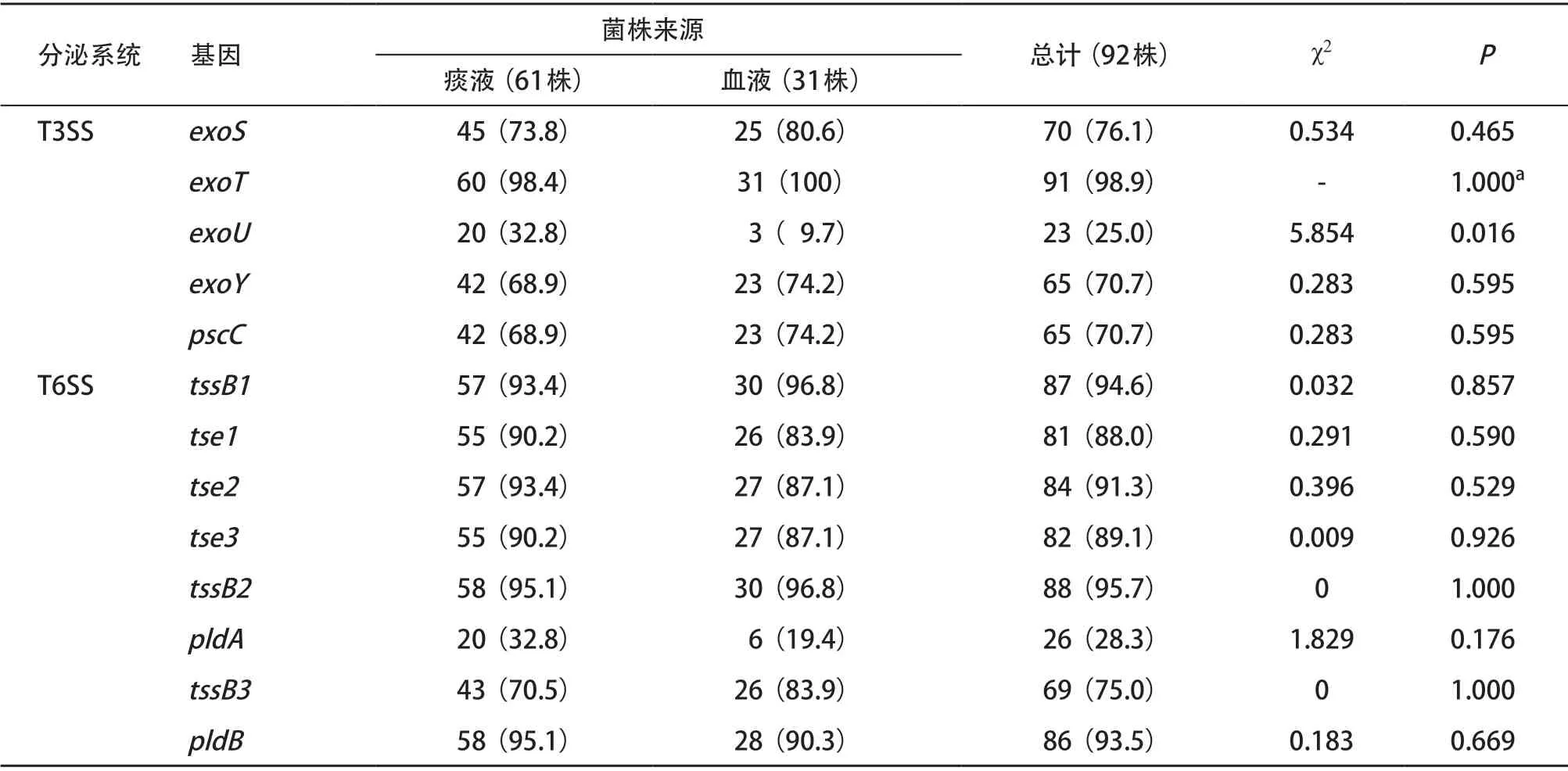
表2 不同来源铜绿假单胞菌T3SS和T6SS效应蛋白基因携带率比较[株(%)]
2.2 不同来源的铜绿假单胞菌对临床常用抗菌药物的耐药率 痰液及血液来源菌株对临床常用抗菌药物亚胺培南、环丙沙星、头孢他啶以及阿米卡星的耐药情况比较结果显示,痰液来源菌株对亚胺培南的耐药率显著高于血液来源菌株(49.2%vs.25.8%),差异有统计学意义(P<0.05),见表3。

表3 不同来源铜绿假单胞菌对常用抗菌药物的耐药率[株(%)]
2.3 T3SS和T6SS效应蛋白编码基因的相关性分析exoU为T3SS的发挥致病作用的主要毒力因子,有文献报道T6SS蛋白编码基因pldA和T3SS蛋白编码基因exoU的携带与非囊性纤维化患者预后恶化有一定的关联[11],为了检测T6SS蛋白编码基因是否与T3SS蛋白编码基因的携带具有相关性,我们对毒力基因exoU和pldA进行了Spearman相关性分析,结果显示pldA和exoU之间呈正相关(r=0.474,P<0.01)。
2.4 T3SS和T6SS效应蛋白编码基因与菌株耐药性的关系 携带不同T3SS及T6SS基因的铜绿假单胞菌对亚胺培南、环丙沙星、头孢他啶、阿米卡星的耐药性分析结果显示,与其他种类的抗菌药物相比,携带T3SS和T6SS效应蛋白编码基因的菌株对碳青霉烯类抗菌药物亚胺培南具有较高的耐药率;但携带不同T3SS及T6SS效应编码基因的菌株对同类抗菌药物的耐药性差异无统计学意义(P>0.05)。见表4、表5。
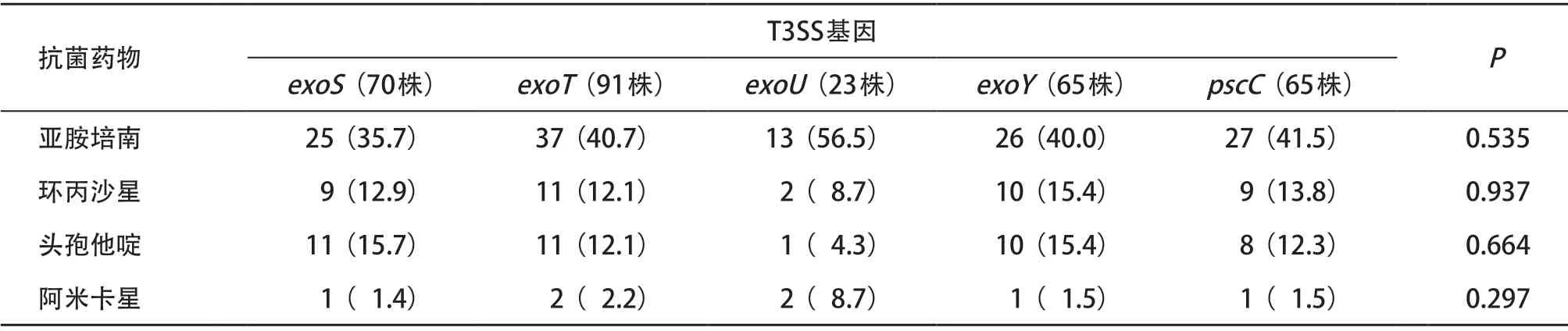
表4 携带T3SS基因铜绿假单胞菌对常用抗菌药物耐药率比较[株(%)]
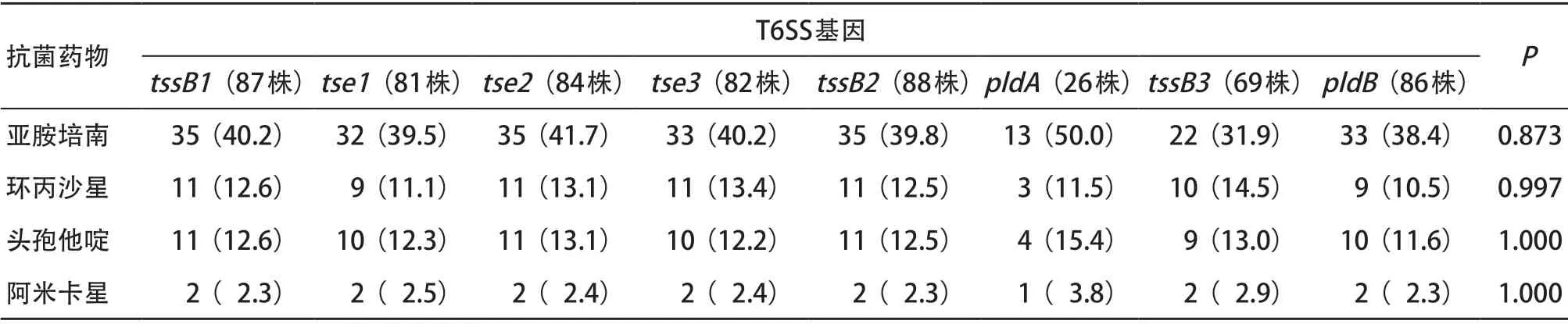
表5 携带T6SS基因铜绿假单胞菌对常用抗菌药物耐药率比较[株(%)]
3 讨论
铜绿假单胞菌是一种引起社区和医院获得性感染的临床常见条件致病菌[12-14]。铜绿假单胞菌可通过分泌系统分泌多种效应蛋白发挥毒性作用,其中T3SS是铜绿假单胞菌与外界环境交流的重要途径之一[5-6]。张秀彩等[15]的研究显示铜绿假单胞菌T3SS毒力基因exoT和exoY的携带率较高,毒力基因与铜绿假单胞菌耐药性相关。T6SS是近年来发现的分泌系统,铜绿假单胞菌可通过T6SS向邻近的病原体传递毒素和蛋白效应因子从而发挥其生存优势,同时T6SS作为铜绿假单胞菌的毒力因子,还能促进其生物被膜的形成,对铜绿假单胞菌的致病性及耐药性发挥重要作用[16]。了解T6SS各毒力因子编码基因的流行分布特性,将有助于铜绿假单胞所致感染的控制。目前国内尚未见评估T6SS多个蛋白编码基因流行率的相关报道。由痰液分离铜绿假单胞菌所引起的呼吸道感染常持续时间久较难清除,同样由铜绿假单胞菌所引起血流感染的患者的治疗具有一定的挑战性,因此了解T3SS和T6SS效应基因与不同标本来源菌株之间的关系值得进一步探究。
本研究对92株铜绿假单胞菌临床分离株中T3SS基因及T6SS基因的流行分布情况进行检测分析,结果显示T3SS效应蛋白编码基因exoT以及T6SS效应蛋白编码基因tssB1、tse2、tssB2、pldB的检出率高达90%以上,表明这些基因在铜绿假单胞菌中具有普遍性,这与已有的研究报道[9,12]基本一致。另外本研究发现痰液标本来源菌株exoU的携带率显著高于血液来源菌株(P<0.05)。HORNA等[17]的研究指出T3SS效应蛋白编码基因exoU是导致宿主细胞死亡的高度细胞毒性表型,携带exoU的菌株被认为具有较强的毒力且常与严重感染有关。而铜绿假单胞菌是引起呼吸道感染的重要原因,痰液是铜绿假单胞菌感染的主要标本分离来源,有研究报道[18]毒力强的铜绿假单胞菌更易引起呼吸道感染,这与本研究结果一致。可能与铜绿假单胞菌所引起的呼吸道感染持续时间长、预后较差有关。我们对2016至2018年不同标本来源铜绿假单胞菌的药敏分析显示痰液来源菌株对亚胺培南的耐药率显著高于血液来源菌株(P<0.05),这可能因为呼吸道分离的铜绿假单胞菌的大部分患者常合并有肺部基础疾病,导致对吸入微生物的清除延迟,治疗周期长、易诱导和筛选出耐药菌[19]。同时这提示我们在临床用药时要关注呼吸道来源铜绿假单胞菌的碳青霉烯类抗菌药物的耐药水平,选择合适药物治疗。另一方面有研究报道[8]铜绿假单胞菌中还存在3种进化上完全不同的T6SS基因群(分别为H1-T6SS、H2-T6SS和H3-T6SS),表达tse1、tse2、tse3、pldA、pldB等相关基因。其中由pldA基因编码的磷脂酶A(PldA)是铜绿假单胞菌H2-T6SS的效应蛋白,可向宿主细胞传递毒素。本研究结果显示包括pldA在内的T6SS相关效应蛋白编码基因在痰液标本和血液标本来源的菌株中不存在显著性差异。有研究报道指出黏液性铜绿假单胞菌中pldA和exoU的存在与非囊性纤维化患者恶化的高风险相关[11],Spearman相关性分析结果显示pldA和exoU表达之间呈正相关性(r=0.474,P<0.01),这提示我们可对pldA在严重感染疾病中的致病作用开展进一步研究。关于T3SS和T6SS效应蛋白编码基因与耐药相关性的探究,之前有研究报道携带exoU基因的菌株耐药性更强[20]。TAKATA等[21]的研究指出碳青霉烯类以及氟喹诺酮类耐药菌株中exoU基因的携带率较高;另外BOULANT等[9]的研究也表明pldA基因在耐药菌株中有较高的携带率。本研究结果显示携带不同T3SS和T6SS基因铜绿假单胞菌对碳青霉烯类抗菌药物亚胺培南的耐药率较高,但不同的基因之间不具有显著性差异(P>0.05)。关于携带高毒力基因exoU或pldA或其他T3SS及T6SS相关基因菌株是否在介导抗菌药物耐药性方面起着相关作用仍需要在未来进行更加广泛且深入的研究。
综上所述,铜绿假单胞菌对T3SS和T6SS大部分效应蛋白编码基因的携带情况具有一定普遍性,但个别毒力较高的分泌系统编码基因的携带需要引起我们的关注。本研究提示T6SS或可作为治疗铜绿假单胞菌感染性疾病的新靶标,从而针对性地控制T6SS相关效应蛋白编码基因携带可能引起的铜绿假单胞菌相关感染疾病,另外在临床治疗中要及时关注痰液来源菌株的毒力特性及耐药性,合理安排用药方案。
猜你喜欢
中国现代医药杂志(2020年10期)2020-12-14
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
天然产物研究与开发(2019年1期)2019-03-01
中成药(2017年9期)2017-12-19
中国医药指南(2017年3期)2017-11-13
中华老年口腔医学杂志(2016年2期)2017-01-15
中国畜牧兽医文摘(2015年9期)2015-12-29
特产研究(2014年4期)2014-04-10
食品工业科技(2014年23期)2014-03-11
