
蓝莓AP2/ERF基因家族鉴定及其在休眠解除过程中的表达分析研究
2022-12-09 07:16马芮李瑞雪郑煜聆李永强
浙江农业科学 2022年12期
马芮,李瑞雪,郑煜聆,李永强,2*
(1.浙江师范大学 化学与生命科学学院,浙江 金华 321004;2.浙江省特色经济植物生物技术研究重点实验室,浙江 金华 321004)
APETALA2/ethylene response factor(AP2/ERF)转录因子家族作为植物中最大的转录因子家族之一,参与植物体内各种信号传导和生长发育调节[1-3]。AP2/ERF家族每个成员包含至少一个由60~70个氨基酸组成的AP2结构域,包括AP2、ERF和RAV亚家族,根据AP2结构域的数目和序列的相似程度将AP2/ERF超家族分为AP2家族、ERF家族和RAV家族[4]。其中AP2家族具有两个AP2/ERF保守结构域,而ERF家族成员包含一个AP2/ERF保守结构域。AP2结构域首次在拟南芥(Arabidopsisthaliana)中被发现AP2蛋白中的重复基序,能够参与花的发育[5]。RAV家族蛋白包含一个B3结构域和一个AP2/ERF结构域[6]。研究表明,ERF转录因子在植物的生长发育以及多种逆境胁迫应答反应中发挥着重要调控作用[7-8]。在乙烯信号转导级联反应中一个重要的转录因子家族是乙烯响应因子(ERFs)。他们作为AP2/ERF超家族的成员,由EILs基因激活并进一步表达,部分ERFs激活GCC-box靶基因,激活功能基因进行各种代谢反应。前人研究表明,从烟草(Nicotianatabacum)中分离出的含有保守ERF结构域的乙烯响应元件结合蛋白(ERF),它可以与乙烯响应元件GCC-box结合,调控乙烯相关基因的表达[9]。
目前,在番茄中已有多个ERFs被鉴定出来,部分可以正向调控果实成熟,而也有部分乙烯响应因子对成熟调控起到负调控作用[10]。研究表明,AP2/ERF超家族成员广泛参与植物乙烯信号途径和花芽萌发以及果实成熟,除此之外还参与植物发育过程中的胁迫、代谢、衰老等。AP2/ERF能够响应各种生物和非生物胁迫,进而调节植物的生长发育,例如低温[11]、干旱、高温[12-13]、盐胁迫等。AP2家族中的基因被证实可以决定玉米穗的分生组织[14],AP2/ERF转录因子通过转录和翻译等多种机制参与调控生长、发育、激素和应激反应等多种生物学过程的基因表达[15-16]。有研究证明,过表达DREB2C时会导致植物种子推迟萌发[17]。Sl-ERF2转基因番茄种子比野生型提早萌发,提高了植株对乙烯的敏感性[18]。ERFs基因广泛存在于高等植物中,可以通过与DNA直接结合,达到调控下游基因转录的目的,也可以调控各种信号转导途径[19-21]。当山毛榉(Faguslongipetiolata)处于休眠状态时,ERF1在种胚中的表达量较低,但休眠解除时,其表达量显著升高[22]。
蓝莓(Vacciniumspp.)是杜鹃花科(Ericaceae)越橘属(Vaccinium)植物,不仅具有较高的营养价值,还具有较高的经济价值,在提高免疫力、增强心肺功能等方面作用显著,被誉为“浆果之王”[23]。由于AP2/ERF转录因子在植物生长发育过程中起到重要作用,本研究借助四倍体蓝莓品种Draper的基因组数据,对蓝莓ERF基因进行鉴定,共得到160个家族成员。进行了多序列比对,保守基序、基因结构、启动子分析,通过转录组数据分析蓝莓AP2/ERF基因家族成员在花芽休眠解除过程中的表达情况。研究结果为进一步研究ERF基因在果树芽休眠调控中的功能奠定基础,为蓝莓产业的发展提供理论基础。
1 材料与方法
1.1 序列数据获取
通过ArabidopsisInformation Resource(TAIR)数据库(http://www.arabidopsis.org),获取拟南芥AP2/ERF基因家族成员,下载蛋白质序列作为参考序列,从越橘基因组数据库(GENOME DATABASE FOR VACCINIUM,GDV)中获取蓝莓蛋白数据库[24]。以拟南芥ERF蛋白序列作参考,使用TBtools软件对蛋白数据库进行BlastP比对[25]。
1.2 蓝莓AP2/ERF基因家族成员的鉴定
选择从上述程序获得的独特序列作为候选AP2/ERF家族蛋白序列。通过CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)排除一些不包含AP2结构域的蛋白序列,通过MEME将筛选出的蓝莓ERF蛋白序列进行保守基序分析,确定蓝莓AP2/ERF基因家族成员。
1.3 蓝莓ERF蛋白的理化性质
利用ProtParam在线软件预测蓝莓中ERF蛋白的氨基酸长度、分子量(MW)、理论等电点(pI)、亲水性(GRAVY)。应用WoLF PSORT软件分析ERF蛋白序列,进行亚细胞定位预测[26]。
1.4 多序列比对及进化分析
通过MEGA5.0分析软件对筛选得到的160个蓝莓ERF全长蛋白序列进行多序列比对并构建系统发育树,方法为:邻接法(Neighbor-Joining method)Bootstrap 1 000次重复[27],借助在线工具EvolView(https://www.evolgenius.info/evolview/#/)绘制进化树图[28]。
1.5 保守基序分析
利用MEME(http://meme-suite.org/tools/meme)在线工具进行蛋白保守结构域预测分析,将保守基序(motif)大小设定为默认参数,输出结构域最大数为5,motif宽度最长为60,其余为默认[29]。
1.6 染色体定位及共线性分析
根据蓝莓基因组信息将160个VcERF基因定位到染色体上。通过运行BlastP来检索蓝莓基因组中的同源基因对(E< 10-5,前五个匹配项)。然后将同源对用作MCScanX的输入文件,分析基因复制的类型[30-31]。采用Circos软件作图[32]。
1.7 基于转录组的VcERFs表达模式分析
根据蓝莓花芽休眠时期的转录组表达数据,获得VcERFs在花芽内休眠解除、生态休眠解除的表达量数据,将FPKM值(fragments per kilobases per millionreads)进行log2a标准化处理,使用TBtools软件的“HeatMap”功能,构建蓝莓ERF基因的表达热图[25]。
1.8 启动子顺式作用元件分析
根据蓝莓ERF基因各转录组表达情况,筛选可能有重要调控作用的蓝莓ERF候选基因。通过蓝莓基因组文件提取VcERFs编码区上游2 000 bp的启动子序列,使用PlantCARE(http://bioinfoatics.psb.ugent.be/webtools/plantcare/html/)网站分析相关蓝莓ERF基因启动子的顺式作用元件,将得到的顺式作用元件进行筛选和统计,用热图展示顺式作用元件的类型及数量[33]。
2 结果与分析
2.1 蓝莓ERF鉴定及蛋白理化性质
通过序列比对及结构域分析,最终筛选获得160个蓝莓ERF蛋白(表1)。使用ExPasy网站在线工具对VcERF蛋白的理化特性进行分析,结果显示,ERF蛋白的氨基酸数量均在154~786个,分子量17.6~86.5 ku,等电点4.71~10.75。不稳定系数变化范围在32.58~74.54,ERF蛋白大部分处于不稳定形态,亲水性值在-1.000至-0.312,说明均为亲水蛋白。亚细胞定位预测分析发现,VcERF蛋白定位在细胞的各个部位,主要定位在细胞核,少数成员定位在细胞质、叶绿体、线粒体和细胞基质。

表1 蓝莓ERF蛋白基本信息
2.2 蓝莓AP2/ERF系统发育进化树
为更深入地了解蓝莓AP2/ERF转录因子同源进化关系,通过MEGA5.0软件构建了160个蓝莓ERF蛋白成员的系统进化树,系统进化树分枝显示(图1),AP2/ERF家族成员被分为3个亚族,蓝莓ERF分布在AP2家族、RAV家族和ERF家族中,AP2家族含有54个ERF蛋白,ERF家族所含的蓝莓成员最多,共82个,RAV家族中有24个ERF蛋白,此类成员含有一个AP2结构域和一个B3结构域。

图1 蓝莓AP2/ERF基因家族成员系统进化树分析
2.3 蓝莓ERF蛋白结构和保守基序分析
为更深入地了解VcERFs中基序(motif)组成的多样性,利用MEME预测蓝莓ERF蛋白质保守基序,共得到5个保守基序(图2),结果表明,所有VcERF蛋白均存在高度保守的AP2结构域(motif 1),motif 1作为DNA的结合模块,对ERF蛋白的功能至关重要。根据保守基序的分布特点,可将蓝莓ERF蛋白分为3组,Ⅰ组成员最多,含有82个ERF蛋白成员,Ⅱ组含有54个,Ⅲ组含有24个。从图2中可知,每条ERF蛋白序列所含有的保守基序的数量以及类型都存在或多或少的差异,其在分布上也存在差异,可能表明每个基序的功能有所差异,其中Ⅲ组成员含有的保守域数量最多,Ⅱ组成员除特有的AP2结构域外,还含有一个特殊的B3结构域(motif 5)。
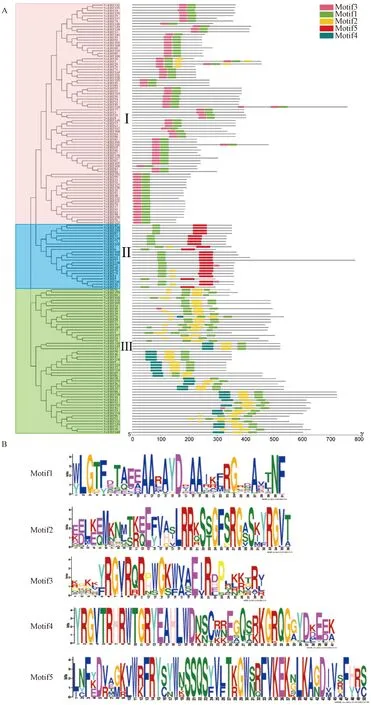
A—蓝莓VcERF进化关系及保守基序;B—蓝莓VcERF Motif序列。
2.4 ERF基因的共线性分析
为了进一步探讨蓝莓AP2/ERF基因家族的起源和可能的进化机制,将蓝莓自身蛋白比对的结果输入MCScanX,输出同源配对文件,分析VcERFs的组内同源配对情况发现,VcERF的同源配对非常复杂,除VcERF5、VcERF6、VcERF15、VcERF21、VcERF23、VcERF26、VcERF27、VcERF38、VcERF39、VcERF50、VcERF51、VcERF52、VcERF58等30个基因没有同源配对外,其他基因家族成员均存在线性关系,见图3中红色连线。蓝莓AP2/ERF基因家族共有45条染色体上存在共线性基因,其中17、27号染色体上共线性基因最多,均有7对共线性基因,其次21、26号染色体上均有6对共线性基因。

图3 蓝莓ERF基因在染色体上的分布和共线性
2.5 基于转录组数据分析VcERF基因的表达特性
基于已有的蓝莓转录组数据分析AP2/ERF基因家族成员在花芽休眠解除下的表达模式(图4),相同亚家族VcERF在休眠解除过程中的表达模式并不相似,暗示其可能在花芽休眠解除过程中发挥不同的作用。ERF家族成员VcERF51、VcERF115、VcERF119、VcERF74、VcERF95、VcERF14、VcERF29、VcERF137、VcERF48、VcERF49、VcERF148、VcERF103在休眠解除过程中表达量较高;VcERF146、VcERF50、VcERF99、VcERF122、VcERF133、VcERF82、VcERF81、VcERF105、VcERF117在内休眠初期表达量水平很高,在内休眠解除过程中下调;其中VcERF81、VcERF105、VcERF117、VcERF65、VcERF106在生态休眠解除过程中的表达量升高,最终也没有恢复到内休眠初期的表达量水平。

a—ERF家族;b—AP2家族;c—RAV家族。
AP2家族成员VcERF109、VcERF114、VcERF33、VcERF140、VcERF70、VcERF145、VcERF157在休眠解除过程中表达量较高;VcERF112、VcERF53的表达量在内休眠解除过程中下调。
第Ⅲ类RAV家族成员VcERF5、VcERF43、VcERF153、VcERF87、VcERF125、VcERF73、VcERF142在内休眠及生态休眠解除过程中几乎不表达;VcERF30、VcERF138、VcERF72、VcERF141随着内休眠的解除表达量上升;VcERF30、VcERF141的表达量在生态休眠解除过程中先下降后上升。
2.6 启动子顺式作用元件分析
为进一步分析蓝莓AP2/ERF家族基因在启动子区域的调控元件特点,对10个ERF基因上游2 000 bp片段进行启动子顺式作用元件分析。结果显示(图5),蓝莓ERF启动子区域含有大量光响应元件和激素响应元件,包括光反应(Box 4、ATCT-motif、MRE、SP1、G-box)等,脱落酸(ABRE)、赤霉素(P-box、TATC-box、GARE-motif)、生长素(TGA-element)、茉莉酸甲酯(CGTCA-motif、TGACG-motif)、水杨酸(TCA-element)等,表明蓝莓ERF基因可能在光响应和激素响应中起重要作用。除此之外,蓝莓ERF基因还含有胁迫响应元件,如低温(LTR)、防御应激(TC-rich repeats)。
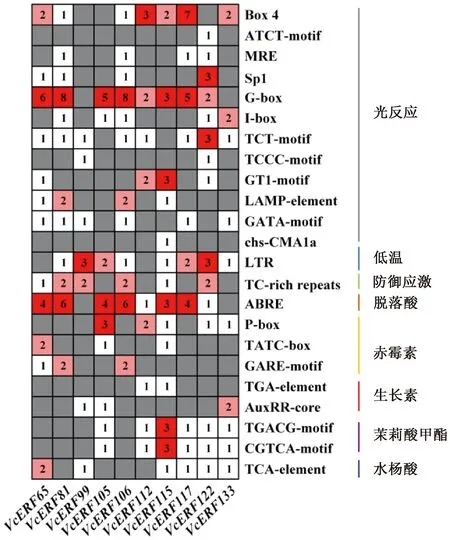
图5 蓝莓ERF基因启动子的顺式作用元件分析
3 讨论
本研究中采用生物信息学方法从蓝莓基因组数据库中鉴定出160个ERF基因,分布于蓝莓48条染色体中的45条上,亚细胞定位预测显示ERF基因主要定位于细胞核。通过生信分析发现,最长的VcERF基因编码786个氨基酸,最短的编码154个氨基酸,分子量分布在17.6~86.5 ku,对VcERF基因的结构分析表明,虽然VcERF基因的长度存在较大差异,但总体上同一亚类的基因具有相似的基因结构,这说明VcERF基因在进化上是保守的。为了进一步研究蓝莓与拟南芥中AP2/ERF家族的同源进化关系,利用邻接法构建AP2/ERF基因家族系统发育树,并根据其分类特点将这些基因分为3组。拟南芥中的一些AP2/ERF基因家族成员功能已经得到验证,根据进化关系中基因的同源性,推测与拟南芥位于同一组的蓝莓ERF基因也具有与其相同或相类似的功能。蓝莓基因组中之所以存在如此丰富的AP2/ERF基因家族成员,一方面可能是由于其属于多倍体物种,本研究用于AP2/ERF基因家族生物信息学预测的基因组来源于四倍体蓝莓品种Draper,多倍体化会影响基因家族的分布和大小;另一方面可能与基因组加倍事件有关,蓝莓基因组经历了至少1次全基因组重复[24]和大量串联重复,串联重复和片段重复是基因家族扩张和进化的重要因素。系统发育分析也表明,许多蓝莓AP2/ERF家族成员具有高度相似性,而一些VcERF基因的相邻染色体位置为串联复制提供了进一步的支持。
通过对ERF基因在蓝莓休眠及解除过程中的转录水平进行研究,发现多数VcERF基因在内休眠及生态休眠解除过程中均能表达。其中VcERF51、VcERF115、VcERF119、VcERF74、VcERF95、VcERF14、VcERF29等基因在内休眠及生态休眠解除过程中表达量较高;VcERF116、VcERF118、VcERF123、VcERF82、VcERF81等基因随着内休眠的解除表达量下降;不同的基因在不同暖温处理时长下表达存在差异。启动子顺式作用元件分析结果显示,10个ERF基因启动子区域存在光响应、激素响应(脱落酸、赤霉素、生长素、茉莉酸甲酯、水杨酸)和胁迫响应元件(低温、防御应激),说明蓝莓ERF可能通过光、激素和胁迫信号调控蓝莓花芽的生长发育和休眠。本研究可为蓝莓ERF的研究提供一定的理论基础。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
南方医科大学学报(2022年3期)2022-04-13
河北果树(2021年4期)2021-12-02
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
实用肿瘤学杂志(2020年4期)2020-12-08
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
今日农业(2019年14期)2019-01-04
商周刊(2017年5期)2017-08-22
作文周刊·小学一年级版(2017年26期)2017-08-10
