
低蛋白饲料添加蛋氨酸和赖氨酸对大口黑鲈生长性能和抗氧化能力的影响
2022-12-03 06:27:10桂聪邓琦琦杨慧君莫爱杰沈志刚袁勇超
华中农业大学学报(自然科学版) 2022年6期
桂聪,邓琦琦,杨慧君,莫爱杰,沈志刚,袁勇超,2
1.华中农业大学水产学院/农业动物遗传育种与繁殖教育部重点实验室/农业农村部淡水生物繁育重点实验室,武汉 430070;2.长江经济带大宗水生生物产业绿色发展教育部工程研究中心/湖北省名优鱼育种与健康养殖工程技术研究中心,武汉 430070
近年来,氮排放引发的水环境污染随水产养殖规模和集约化程度的不断扩大而日趋严重。水产养殖对象对饲料蛋白质的高效利用是充分发挥生长性能并降低氮排放的关键。降低日粮中蛋白水平不仅可以降低成本,还可以提高饲料中蛋白质利用效率以及降低氨氮排放[1],但是随着饲料中蛋白质含量的降低,会引起氨基酸比例失调等情况,从而影响养殖对象正常生长[2]。因此,在不影响养殖鱼类正常生长的基础上,降低饲料蛋白水平、提高蛋白质的利用、减少氨氮排放等问题已成为热门话题。功能性氨基酸对改善低蛋白日粮的适口性和提高低蛋白日粮的采食量以及耐受性尤为重要。在降低饲料蛋白水平或鱼粉替代的基础上添加功能性氨基酸如限制性氨基酸[3]赖氨酸和蛋氨酸能维持鱼类的生长性能[4]、提高鱼类对低蛋白日粮蛋白质的利用效率[5]。如在低蛋白饲料中补充必需氨基酸不仅能促进大口黑鲈(Micropterus salmoides)[1]和 虹 鳟(Oncorhynchus mykiss)[6]的生长,而且对草鱼(Ctenopharyngodon idella)幼鱼饲料系数、摄食率、体成分没有显著性影响[7]。
大口黑鲈(Micropterus salmoides)属于典型的肉食性鱼类,对饲料中蛋白水平要求较高,其蛋白最适需求量的问题仍存在争议,提高饲料中蛋白水平能有效促进大口黑鲈生长,但会出现蛋白质利用效率下降的现象[8],容易造成浪费、增加成本。而低蛋白饲料会造成大口黑鲈的生长性能显著性降低以及免疫力下降[9]。因此,本试验研究低蛋白饲料中添加必需氨基酸对大口黑鲈幼鱼生长、生产性能及其抗氧化能力的影响,旨在为大口黑鲈优质配合饲料的研制提供参考依据。
1 材料与方法
1.1 试验饲料配制
饲料原料由武汉澳华科技有限公司提供,试验饲料配方如表1所示。其中维生素预混料组成成分:维生素A 16 000 IU/kg;维生素D38 000 IU/kg;其他成分(mg/kg):维生素B117.60;维生素B248.00;维生素B630.00;维生素B120.25;维生素E 156;维生素C 800;维生素K314.70;烟酸胺79.00;泛酸钙73.50;叶酸6.50;生物素0.64。矿物质预混料组 成 成 分(mg/kg):镁(MgSO4·H2O)52.50;锌(ZnSO4·5H2O)34.40;铜(CuSO4·5H2O)2.10;锰(MnSO4·75H2O)6.10;碘(Ca(IO3)2)1.60;铁(FeSO4·7H2O)21.10;硒(Na2SeO3)0.19;钴(CoCl2·5H2O)0.24。设置46%(对照)、41%(LP1)和36%(LP2)3 个饲料蛋白质梯度。在41%和36%蛋白饲料的基础上补充相应的晶体赖氨酸与蛋氨酸,使赖氨酸与蛋氨酸的含量与对照组中含量保持一致,分别记作LP1+AA 和LP2+AA 组。依据饲料配方表准确称取各组饲料原料过0.425 mm孔径筛网后充分混匀,加入蒸馏水约300 g/kg,通过球团机(霍巴特混合机,型号A200),将其压制成3 mm 直径的颗粒饲料,45 ℃烘干,封于自封袋并于−20 ℃冰箱中保存。

表1 各组饲料配方及营养成分(风干质量)Table 1 Feed formula and nutrients in different group(air dry basis)
1.2 养殖试验
规格相近、健康无病的大口黑鲈500尾选购自湖北黄优源渔业发展有限公司,于华中农业大学水产养殖基地塑料养殖桶(直径120 cm,高130 cm)中进行养殖,试验过程中养殖桶是静水模式,每次喂食前使用虹吸管清理粪便、残饵等污染物,换水40%~50%,维持良好的养殖水质。暂养2 周后,从中随机选取450 尾初始体质量(31.52±0.62)g 的试验鱼,每个养殖桶放30 尾,每组饲料设置3 个重复。于每天08:00 和18:00 投喂幼鱼体质量3%~6%的试验饲料。试验过程保持曝气,水温维持在25~28 ℃,氨氮为(0.25±0.12)mg/L,pH 为7.2±0.2,饲养试验为期8周。
1.3 样品采集及分析
养殖周期结束后停喂24 h,取出所有试验鱼,使用MS-222 溶液麻醉,记录大口黑鲈的终末体质量、存活数量以及体长等数据。每个试验组随机选取3尾作为全鱼样品放入−80 ℃冰箱冷冻保存,每个养殖桶挑选3 尾试验鱼无菌操作下尾静脉采血后分离内脏和肝脏并称质量计算形体学指标,其余试验鱼采取肝脏、肌肉组织及血样,血样经过离心,分离出血清,放入−80 ℃冰箱保存用于常规营养成分、抗氧化酶及转氨酶活性的检测。
1.4 测定指标与计算方法
1)测定指标。试验结束对大口黑鲈增重率(WGR)、特定生长率(SGR)、饲料系数(FCR)、存活率(SR)、脏体比(VSI)、肝体比(HIS)、肥满度(CF)、蛋白质保留率(PR)、摄食量(FI)等指标进行测定,具体的计算公式参考陈乃松等[1]的方法。
2)测定方法。全鱼和肌肉中的水分测定采用105 ℃烘干法;样品中粗蛋白含量测定采用凯氏定氮法;粗脂肪测定方式使用索氏抽提法,粗灰分测定采用马弗炉550 ℃高温灼烧法;饲料总能量的测定采用氧弹法;饲料中赖氨酸和蛋氨酸的含量用氨基酸自动分析仪(Hitachi,Model 835-50,Hitachi,Tokyo,Ja⁃pan)分析。使用南京建成生物工程研究所的检测试剂盒对血清和肝脏中谷草转氨酶(GOT)活性(C010-2-1)、谷丙转氨酶(GPT)活性(C009-2-1)、超氧化物歧化酶(SOD)活性(A001-3)、过氧化氢酶(CAT)活性(A007-1-1)、总抗氧化能力(T-AOC)(A015-2-1)、丙二醛(MDA)含量(A003-1)、活性氧(ROS)水平(E004-1)进行测定。样品的前期处理以及测定方法参照试剂盒说明书,大口黑鲈肝脏和血清转氨酶和抗氧化指标测定之前选取一定量的肝脏组织,加入预冷生理盐水以体积1∶9 制成匀浆,在4 ℃下3 000 r/min离心机进行离心10 min,取离心后上层清液,按一定比例稀释后用于后续转氨酶和抗氧化指标活性分析。
1.5 试验数据处理
按照指标的公式对所得的数据进行运算整理,使用SPSS Statistics 25 软件进行单因素方差分析(One-way ANOVA),差异显著性分析用Duncan’s法进行多重比较,显著性水平α=0.05,所有数据以“平均值±标准差”表示。
2 结果与分析
2.1 低蛋白饲料添加赖氨酸与蛋氨酸对大口黑鲈生长性能的影响
由表2 可知,LP1+AA 组大口黑鲈的FBW、WGR、SGR、FCR 和PR 与对照组之间不存在显著差异(P>0.05);其他各组的FBW、WGR 和SGR 与对照组相比均显著性降低,FCR和PR显著性升高(P<0.05)。添加赖氨酸与蛋氨酸的LP1+AA 和LP2+AA 组较未添加氨基酸的LP1 和LP2 组的FBW、WGR、SGR 有一定的升高,FCR 和PR 相对降低,低蛋白饲料组大口黑鲈的FI 均低于对照组,添加赖氨酸与蛋氨酸的试验组与未添加的处理组相比有所升高,但均无显著性差异(P>0.05)。各组大口黑鲈的SR均为100%(P>0.05)。
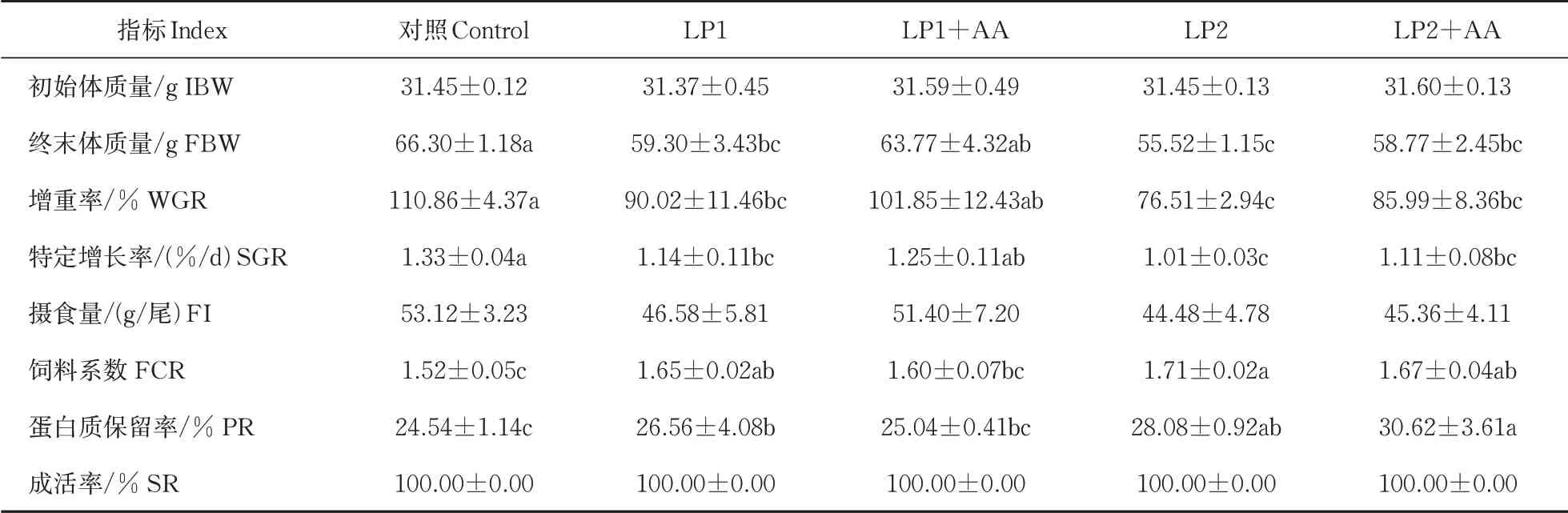
表2 低蛋白饲料中添加赖氨酸与蛋氨酸后大口黑鲈生长性能的变化(n=3)Table 2 Changes of growth performance of largemouth bass supplemented with lysine and methionine in low protein diet(n=3)
2.2 低蛋白饲料添加赖氨酸与蛋氨酸对大口黑鲈体组成和形体学指标的影响
由表3 可知,与对照组相比,大口黑鲈肌肉中粗蛋白含量除LP1+AA 组没有显著性差异外(P>0.05),其余各组均显著性降低(P<0.05);降低饲料蛋白水平全鱼中粗蛋白显著性低于对照组(P>0.05)。添加赖氨酸与蛋氨酸的LP1+AA 和LP2+AA 组与对应的LP1 和LP2 组相比肌肉中粗蛋白的含量没有显著性变化(P>0.05),但全鱼中粗蛋白含量升高,LP2+AA 组显著性高于LP2 组(P<0.05),大口黑鲈肌肉和全鱼其他营养成分均未出现显著影响(P>0.05)。

表3 低蛋白饲料中添加赖氨酸与蛋氨酸后大口黑鲈肌肉与全鱼营养成分的变化Table 3 Changes of muscle and whole fish nutrient composition of largemouth bass supplemented with lysine and methionine in low protein diet %
由表4可得,降低饲料蛋白显著降低了大口黑鲈幼鱼的肝体比(HSI)(P<0.05),但41%低蛋白水平添加赖氨酸与蛋氨酸后大口黑鲈形体学指标与对照组相比没有显著性差异(P>0.05)。肥满度(CF)和脏体比(VSI)均无显著性差异。
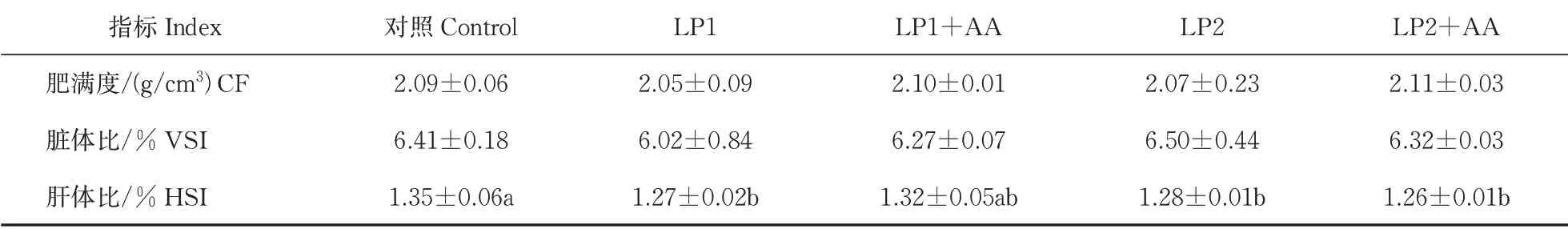
表4 低蛋白饲料中添加赖氨酸与蛋氨酸后大口黑鲈形体学指标的变化Table 4 Changes of physique indices of largemouth bass supplemented with lysine and methionine in low protein diet
2.3 低蛋白饲料添加赖氨酸与蛋氨酸对大口黑鲈转氨酶活性的影响
由图1 可知,大口黑鲈肝脏中谷草转氨酶(GOT)和谷丙转氨酶(GPT)的活性差异不显著(P>0.05);对照组血清中GPT 的活性显著低于LP2组(P<0.05),与其他试验组无显著性差异。
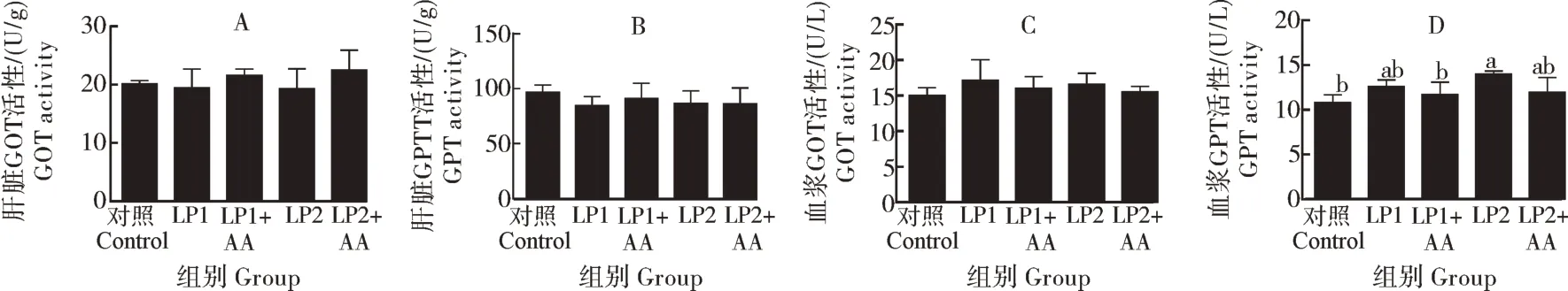
图1 低蛋白饲料中添加赖氨酸与蛋氨酸大口黑鲈幼鱼转氨酶活性的变化Fig.1 Changes of transaminase activity of largemouth bass fed low protein diet supplemented with lysine and methionine
2.4 低蛋白饲料添加赖氨酸与蛋氨酸对大口黑鲈幼鱼肝脏抗氧化能力的影响
由表5 可知,与对照相比,LP1+AA 肝脏中活性氧(ROS)与对照组的比值、总抗氧化能力(T-AOC)、过氧化氢酶(CAT)活性和丙二醛(MDA)含量均无显著性差异(P>0.05);LP2 和LP2+AA 组ROS 均显著性升高(P<0.05);降低日粮中蛋白水平后肝脏中T-AOC显著性降低(P<0.05);LP2 组肝脏中MDA含量和CAT 的活性相比于对照组差异显著(P<0.05),其他各组间差异均不显著;肝脏中超氧化物歧化酶(SOD)的活性在各组间均无显著性差异(P>0.05)。
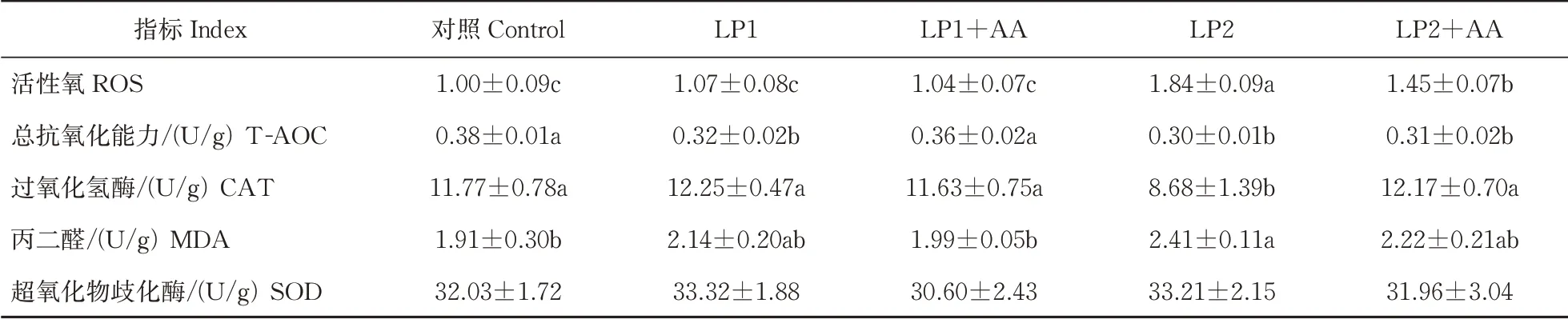
表5 低蛋白饲料添加赖氨酸与蛋氨酸后大口黑鲈肝脏抗氧化能力的变化Table 5 Changes of liver antioxidant capacity of largemouth bass supplemented with lysine and methionine in low protein diet
2.5 低蛋白饲料添加赖氨酸与蛋氨酸对大口黑鲈血清抗氧化能力的影响
由表6 可知,饲料蛋白水平由46%降至36%时,大口黑鲈血清中ROS 的含量发生显著性变化(P<0.05),但降低至41%时却无显著性差异(P>0.05)。饲料蛋白水平由46%降低至41%或36%时血清中CAT 的活性显著性降低(P<0.05),但补充必需氨基酸得到明显改善与对照组之间无显著性差异(P>0.05)。与46%蛋白水平相比,其他各组血清中T-AOC、MDA 和SOD 均无显著性差异。
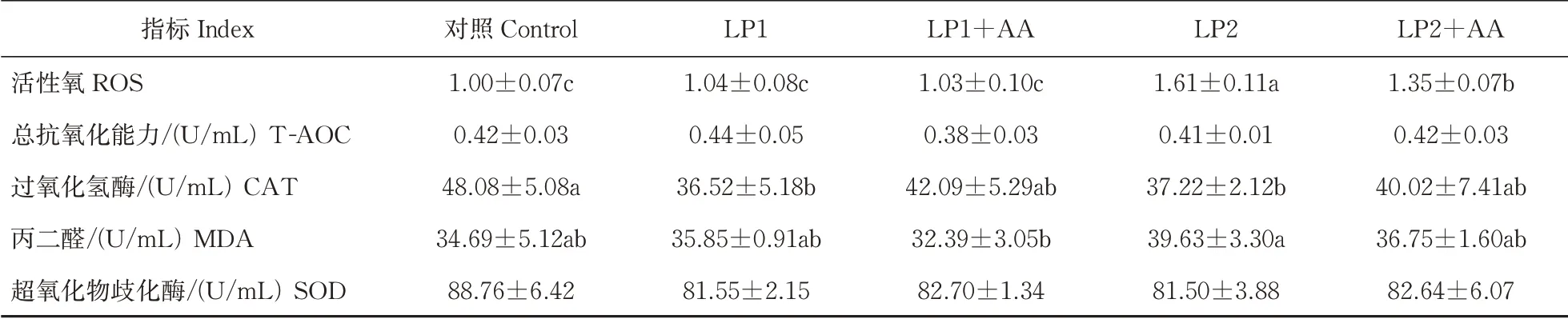
表6 低蛋白饲料中添加赖氨酸与蛋氨酸大口黑鲈血清抗氧化能力的变化Table 6 Changes of serum antioxidant capacity of largemouth bass supplemented with lysine and methionine in low protein diet
3 讨 论
本研究结果显示降低饲料中蛋白质含量会导致大口黑鲈幼鱼生长性能和饲料利用效率显著下降,这与降低饲料蛋白质水平对宝石鲈(Scortum bar⁃coo)[10]的生长也造成类似的影响相一致。但在低蛋白饲料中补充赖氨酸与蛋氨酸可以改善大口黑鲈幼鱼的生长性能,这与虹鳟(Oncorhynchus mykiss)[11]和军曹鱼(Rachycentron canadum)[12]的相关研究结果基本一致。有关研究显示,降低饲料中蛋白质水平再添加适宜的赖氨酸和蛋氨酸等限制性氨基酸,能够维持部分鱼类正常生长[11,13],但是当饲料中蛋白水平降低一定量后,即使补充赖氨酸和蛋氨酸等限制性氨基酸,鱼类的生长性能也会因为其他氨基酸的缺乏而难以得到维持。如陈乃松等[1]的研究中,当饲料中蛋白水平降低至35%时补充必需氨基酸后大口黑鲈生长性能仍显著低于45%蛋白水平组;Yamamoto 等[14]将虹鳟(Oncorhynchus mykiss)饲料蛋白水平由45%降低至30.6%时,晶体氨基酸添加量需达到需求量的1.2 倍才能维持虹鳟正常生长,并且能提高机体对饲料中蛋白质的利用效率。此外,刘梦梅[7]通过分析草鱼(Ctenopharyngodon idellus)生长性能,发现饲料粗蛋白含量为29%的试验组在未添加氨基酸的情况下,优于粗蛋白含量为31%的饲料组及其添加赖氨酸与蛋氨酸的饲料组。这种现象表明饲料中超过草鱼机体需求量的蛋白质和氨基酸会增加试验鱼的代谢负荷,从而使试验鱼的生长性能下降。
本研究结果显示,饲料蛋白质含量的降低会导致全鱼粗蛋白含量下降,通过补充赖氨酸和蛋氨酸后有一定的改善作用。而程小飞等[15]有关湘华鲮(Sinilabeo decorustungting)研究结果显示,饲料中蛋白水平的变化不会引起鱼体粗蛋白含量的显著变化,但会对肌肉粗蛋白的含量造成显著性影响。在本研究中降低饲料蛋白水平对大口黑鲈幼鱼肌肉和鱼体的粗脂肪、粗灰分和水分等营养成分未造成显著影响,而刘梦梅[7]以草鱼为研究对象的相关试验也出现类似的结果。本研究中低蛋白组大口黑鲈HSI显著低于对照组,但41%蛋白水平组添加氨基酸后与对照组相比没有显著性差异。分析试验结果可知补充赖氨酸与蛋氨酸能够有效改善试验鱼鱼体蛋白质的含量和鱼体形体学指标,这在有关虹鳟[11,13]和草鱼[7]的试验中也出现类似的结果。
转氨酶是机体氨基酸代谢过程中的关键酶,正常生理状态下血清中的含量相对较低,但在肝、肾、心肌等组织中的含量会相对较高[16]。当肝脏受到外界刺激受损时,转氨酶含量会超过正常范围,转氨酶会从组织进入到血清,从而导致血清中转氨酶含量上升[17]。本研究中降低饲料蛋白含量对大口黑鲈肝脏造成一定程度的损伤,但平衡氨基酸后血清GPT降低,与对照组无显著性差异,这一结果表明降低饲料蛋白含量基础上添加蛋氨酸与赖氨酸在一定程度上可以减缓肝脏受损程度。郭凯等[18]降低荷斯坦公牛饲料中蛋白水平也出现血清中GPT含量升高的类似情况。当养殖对象肝脏出现损伤或者机体蛋白质代谢能力提高时都会引起血清中GPT 和GOT 活性上升[19]。随着饲料中蛋白质水平的升高,出现血清GPT 活性升高的现象,说明饲料中蛋白质水平过高时,会引起机体代谢负荷变大从而造成肝脏损伤。本试验在降低饲料粗蛋白含量的同时补充氨基酸,可以使大口黑鲈血清GPT含量下降。在以红鳍东方鲀(Takifugu rubripes)[20]为研究对象的试验中,向饲料中添加蛋氨酸与赖氨酸,红鳍东方鲀血清中GPT的活性发生类似的下降情况。
T-AOC 的变化侧面反映机体受到外界刺激时,对自由基进行代谢以及代偿能力的高低。当机体脂质过氧化损伤时提高SOD 和T-AOC 的活性做出应激反应[21]。MDA 含量在当机体发生脂质过氧化后会发生明显变化,含量变化越显著说明脂质氧化程度越深[22]。本研究中,低蛋白饲料组血清CAT 活性和肝脏T-AOC 与对照组相比都降低,MDA 在血清和肝脏中的含量均上升;补充赖氨酸与蛋氨酸的饲料组出现肝脏中T-AOC 升高,血清CAT 活性升高,肝脏和血清MDA 含量均降低等现象。降低鳙(Aris⁃tichthys nobilis)[23]日粮中蛋白含量也出现了机体抗氧化性能下降的情况。本研究中添加氨基酸能改善低蛋白饲料对大口黑鲈幼鱼抗氧化性能的影响、减缓大口黑鲈机体脂质氧化过程的损伤,但是当饲料蛋白水平降低10%并添加氨基酸与对照组相比,肝脏中T-AOC 和ROS 仍然存在显著差异,血清中ROS 水平显著升高。研究结果表明,饲料蛋白含量从46%降低至41%并补充适量的必需氨基酸对大口黑鲈生长性能等没有显著性影响;但饲料粗蛋白含量降低至36%并补充赖氨酸和蛋氨酸,不能有效缓解饲料粗蛋白水平过低对生长性能造成的负面影响。
猜你喜欢
垂钓(2023年11期)2024-01-21 16:07:04
基层中医药(2021年6期)2021-11-02 05:46:10
创新作文(小学版)(2018年10期)2018-08-20 03:11:38
广东饲料(2016年3期)2016-12-01 03:43:13
广东饲料(2016年3期)2016-12-01 03:43:11
江苏农业科学(2016年9期)2016-11-28 15:23:56
动物营养学报(2015年10期)2015-12-01 03:30:54
爆笑show(2015年8期)2015-09-22 17:40:01
应用化工(2014年10期)2014-08-16 13:11:29
中国酿造(2014年9期)2014-03-11 20:21:03
