
黄玛草蛉幼虫对草地贪夜蛾卵和低龄幼虫的捕食能力
2022-11-28 07:36施琳琳李子园林丹敏陆永跃陈科伟
昆虫学报 2022年10期
施琳琳,李子园,林丹敏,陆永跃,*,陈科伟,2,*
(1.华南农业大学植物保护学院,广州 501642;2.广东省生物农药创制与利用重点实验室,广州 501642)
草地贪夜蛾Spodopterafrugiperda是一种原产于美洲热带和亚热带地区的杂食性害虫(Sparks,1979;Todd and Poole,1980)。该虫具有寄主范围广、繁殖力高、迁飞能力强、暴发性为害等特点(Roseetal.,1975;Johnson,1987;Goergenetal.,2016;Montezanoetal.,2018;王磊等,2019),是联合国粮农组织向全球预警的跨国界迁飞性农业重大害虫。我国于2019年1月11日首次在云南省江城县发现该虫幼虫为害,截止到2019年10月8日迁飞扩散至我国26个省(自治区、直辖市)1 518个县(区),累计发生108万ha(姜玉英等,2019),并已在云南、广东、海南、福建、广西、四川和贵州中国南方7省(区)周年繁殖(王磊和陆永跃,2020;姜玉英等,2021),对我国农作物的安全生产构成严重威胁。
生物防治在害虫的可持续控制中起着基础性作用。在草地贪夜蛾入侵我国后,国内学者陆续开展了本地天敌资源的挖掘与评价工作,其中涉及的捕食性天敌有半翅目蠋蝽Armachinensis(唐艺婷等,2019a;王燕等,2019)、益蝽Picromeruslewisi(唐艺婷等,2019b;王燕等,2020)、叉角厉蝽Eocantheconafurcellata(范悦莉等,2019;唐敏等,2019)、黄带犀猎蝽Sycanuscroceouittatus(王亚楠等,2020)、大红犀猎蝽Sycanusfalleni(侯峥嵘等,2020)、东亚小花蝽Oriussauteri(代晓彦等,2019,2020;赵雪晴等,2019;孙贝贝等,2020),脉翅目中华通草蛉Chrysoperlasinica(黄海艺等,2020)、大草蛉Chrysopapallens(徐庆宣等,2019;李萍等,2020;曹雯星等,2020)、丽草蛉Chrysopaformosa(李玉艳等,2021),以及鞘翅目七星瓢虫Coccinellaseptempunctata(孔琳等,2019)、龟纹瓢虫Propylaeajaponica(蒋骏等,2020)、异色瓢虫Harmoniaaxyridis(孔琳等,2019;刘本菊等,2020;赵英杰等,2020)、多异瓢虫Hippodamiavariegata(孔琳等,2019)等。这些研究为利用本地捕食性天敌昆虫防治草地贪夜蛾提供了重要的基础信息。
黄玛草蛉Malladabasalis属脉翅目(Neuroptera)草蛉科(Chrysopidae)玛草蛉属Mallada,国内主要分布于华南及台湾地区(杨星科等,2005;李志刚等,2011;Chengetal.,2012),是农林生态系统常见的一种本地优势天敌。研究表明黄玛草蛉对螺旋粉虱Aleurodicusdispersus(李志刚等,2011)、米蛾Corcyracephalonica(李水泉等,2012)、埃及吹绵蚧Iceryaaegyptiaca(叶静文等,2013)、柑桔粉蚧Planococcuscitri(叶静文等,2012)、神泽氏叶螨Tetranychuskanzawai和柑橘全爪螨Panonychuscitri(Chengetal.,2012)、玉米蚜Rhopalosiphummaidis和甘蓝蚜Brevicorynebrassicae周娟等,2020)等害虫均有较强的捕食作用。但目前尚未见国内外有黄玛草蛉对草地贪夜蛾捕食能力方面的报道。为充分挖掘、利用本地优质天敌资源,本研究采用捕食功能反应方法评价黄玛草蛉对草地贪夜蛾卵及低龄幼虫的捕食能力,为今后利用该天敌防控草地贪夜蛾提供相关理论依据及实践参考。
1 材料与方法
1.1 供试虫源
黄玛草蛉:由广东省科学院动物研究所提供,在室内用米蛾卵作为中间猎物饲养,选取个体大小相接近的2龄及3龄幼虫供实验。饲养条件为:温度26℃±1℃,相对湿度60%~70%,光周期16L∶8D(宁波莱福科技有限公司生产MGC-300H型人工气候箱)。
草地贪夜蛾:草地贪夜蛾幼虫于2019年4月在广州市花都区花山镇红群村玉米地(23°48′N,113°27′E)采获,带回室内采用草地贪夜蛾人工饲料饲养(李子园等,2019),饲养环境条件同黄玛草蛉。
1.2 黄玛草蛉对草地贪夜蛾的捕食功能反应测定
选择黄玛草蛉2和3龄幼虫与草地贪夜蛾卵及1和2龄幼虫进行组合,共设6个实验组合,每个组合的猎物密度设置见表1。实验前将1头经24 h饥饿处理的黄玛草蛉幼虫与相应龄期、密度的草地贪夜蛾幼虫放入同一培养皿(直径9 cm)中,同时在培养皿中投放足量的新鲜人工饲料供草地贪夜蛾幼虫取食,避免其互相残杀,然后用保鲜膜封口,用昆虫针在膜上扎适量透气孔。24 h后检查各处理草地贪夜蛾卵或幼虫被捕食的数量。各处理重复10次。

表1 测试黄玛草蛉幼虫对草地贪夜蛾卵及幼虫捕食功能反应的实验设计Table 1 Experimental design for testing the predatory functional responses of Mallada basalis on the eggs and larvae of Spodoptera frugiperda
1.3 数据处理
1.3.1黄玛草蛉对不同密度草地贪夜蛾卵及幼虫捕食率差异统计分析:捕食量数据采用Bartlett法进行方差齐性检验,如方差齐性则采用单因素方差分析(one-way ANOVA),如方差不齐则采用Kruskal-Wallis法进行非参数检验;捕食率数据经平方根后反正弦函数转换再进行方差齐性及各处理间的差异性检验,检验方法同捕食量数量处理;采用Tukey氏法进行均数间的差异显著性检验。黄玛草蛉同一龄期幼虫对相同密度的草地贪夜蛾1与2龄幼虫的捕食率差异采用独立样本t检验,在本实验中当黄玛草蛉为2龄幼虫时,草地贪夜蛾1与2龄幼虫的相同密度设置有20,40及80头/皿;当黄玛草蛉为3龄幼虫时,草地贪夜蛾1与2龄幼虫的相同密度设置有20,40及80头/皿。数据处理软件为SAS 9.0(SAS,2008)。
1.3.2捕食功能反应类型判断及模型拟合:捕食功能反应类型判断及模型拟合均在“R”软件内运行“frair”包进行数据处理(Pritchardetal.,2017)。目前文献报道的功能反应类型有3种,在6个实验组合中黄玛草蛉2和3龄幼虫的捕食量均随猎物密度的升高而不断增加,最后趋于饱和状态,呈典型的Ⅱ型或Ⅲ型功能反应类型,因而排除Ⅰ型功能反应的可能性,侧重于Ⅱ型或Ⅲ型功能反应模型的拟合。
采用Juliano(2001)提出的Logistic回归方法拟合捕食率的多项式模型(公式1),运行“R”软件“frair”包内的函数“frair_test”进行拟合。
(1)
其中,Ne为被捕食猎物数量,N0为初始猎物数量,Ne/N0为猎物被捕食率,P0为截距,P1,P2与P3分别为N0的线性系数、二次方系数与三次方系数。用P1和P2来确定功能反应的类型,当线性系数为负时(P1<0),捕食者表现出Ⅱ型功能反应,说明猎物消耗的比例随猎物密度的增加而下降;当P1>0且P2<0时,捕食者表现出Ⅲ型功能反应(Juliano,2001)。
我们的预实验结果表明,黄玛草蛉2和3龄幼虫对草地贪夜蛾卵及1和2龄幼虫的捕食行为符合Ⅱ型功能反应。Juliano(2001)提出,Holling圆盘方程(Holling,1959)在实际应用中有一定的局限性,仅适用于猎物密度固定的实验条件,而由于在本研究中被捕食的猎物不再进行补充,猎物的密度是在一个动态变化过程中,因而采用Rogers(1972)提出的随机捕食者方程(Rogers’ random predator equation)(公式2)进行Ⅱ型功能反应模型的拟合。数据在“R”软件“frair”包中采用函数“frair_fit”进行Rogers随机捕食模型拟合。
Ne=N0×(1-e-a×(T-Th×Ne))
(2)
式中Ne为被捕食猎物数量,N0为初始猎物数量,Th为捕食者处理1头猎物的时间,a为瞬时攻击率,T为实验观察总时间(本研究中T为24 h)。在R包“frair”中采用函数“frair_compare”对不同龄期黄玛草蛉幼虫对草地贪夜蛾相同虫态的功能反应参数(瞬时攻击率和处理时间)进行差异显著性检验(Juliano,2001;Pritchardetal.,2017)。观察的捕食量及拟合的功能反应曲线采用R包“ggplot2”绘图。
2 结果
2.1 黄玛草蛉对草地贪夜蛾卵以及1龄和2龄幼虫的捕食量
经方差分析或Kruskal-Wallis法检验表明,黄玛草蛉2龄或3龄幼虫对不同密度草地贪夜蛾卵(2龄:F=32.02,df=5,54,P<0.01;3龄:χ2=48.07,df=5,P<0.01)以及1龄(2龄:χ2=12.75,df=5,P=0.0157;3龄:χ2=38.17,df=5,P<0.01)、2龄幼虫(2龄:χ2=23.24,df=5,P<0.01;3龄:χ2=40.26,df=5,P<0.01)的捕食率之间存在显著差异(表2和3)。在各组合处理中,黄玛草蛉幼虫的捕食率总体随着猎物密度的升高而逐步下降。在实验设置的猎物最高密度时,黄玛草蛉2龄幼虫对猎物的捕食率均低于40%,3龄幼虫对猎物的捕食率均低于50%。
当猎物密度分别为20和40头/皿时,黄玛草蛉2龄幼虫对草地贪夜蛾1龄与2龄幼虫的捕食率介于63.00%~73.50%之间,两猎物密度之间没有显著差异(20头/皿:t=0.16,df=18,P=0.8780;40头/皿:t=0.50,df=18,P=0.6202);而当猎物密度升至80头/皿时,黄玛草蛉2龄幼虫对草地贪夜蛾2龄幼虫的捕食率为39.63%,明显低于其对1龄幼虫的捕食率(66.50%)(t=3.09,df=18,P=0.0110)(表2)。当猎物密度分别为20和80头/皿时,黄玛草蛉3龄幼虫对草地贪夜蛾1龄与2龄幼虫的捕食率介于79.50%~99.00%之间,相同密度不同龄期之间没有显著差异(20头/皿:t=0.33,df=18,P=0.7484;80头/皿:t=0.18,df=18,P=0.8571);而当猎物密度为40头/皿时,黄玛草蛉3龄幼虫对草地贪夜蛾2龄幼虫的捕食率明显低于其对1龄幼虫的捕食率(t=2.59,df=18,P=0.0237)(表3)。
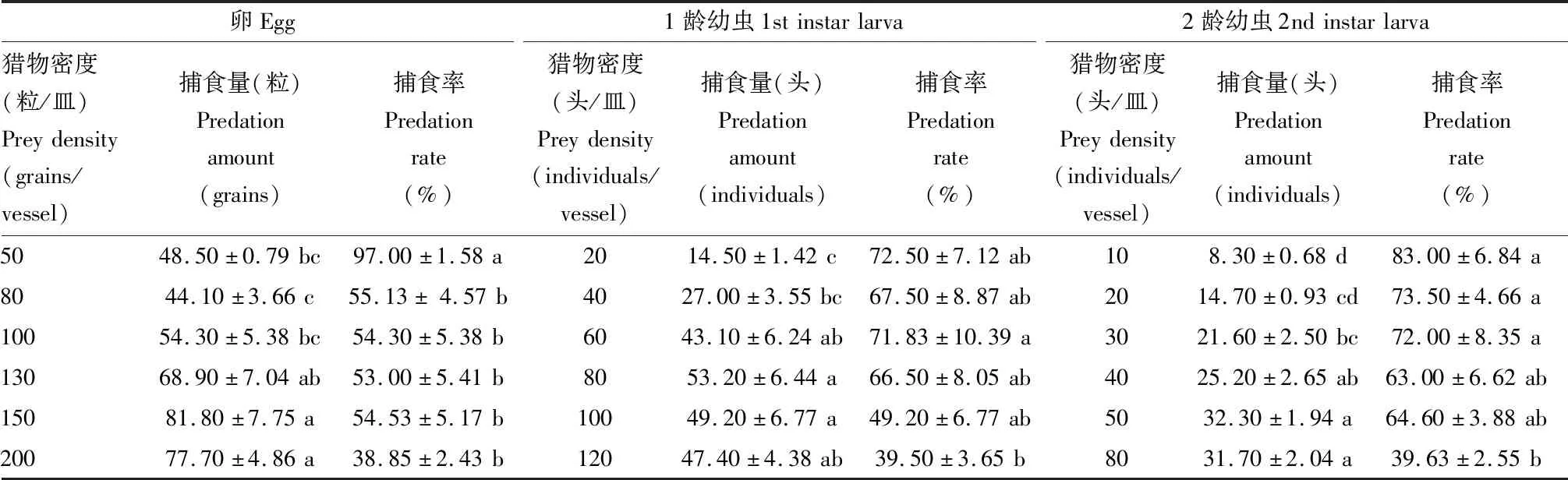
表2 黄玛草蛉2龄幼虫对草地贪夜蛾卵及1和2龄幼虫的捕食量和捕食率Table 2 Predation amounts and predation rates of the 2nd instar larvae of Mallada basalis on the eggs,and the 1st and 2nd instar larvae of Spodoptera frugiperda

表3 黄玛草蛉3龄幼虫对草地贪夜蛾卵及1和2龄幼虫的捕食量和捕食率Table 3 Predation amounts and predation rates of the 3rd instar larvae of Mallada basalis on the eggs, and the 1st and 2nd instar larvae of Spodoptera frugiperda
2.2 黄玛草蛉对草地贪夜蛾卵以及1龄和2龄幼虫的捕食功能反应
在黄玛草蛉2和3龄幼虫与草地贪夜蛾卵以及1和2龄幼虫分别组合的6个实验中,黄玛草蛉对草地贪夜蛾的捕食量均随着猎物密度的增加而增加然后逐渐平缓,拟合的Logistic回归方程的线性系数P1<0,符合Ⅱ型功能反应(表4;图1和2)。
在相同猎物龄期下,黄玛草蛉2龄幼虫对草地贪夜蛾卵以及1和2龄幼虫的瞬时攻击率a均显著低于黄玛草蛉3龄幼虫(草地贪夜蛾卵:Z=-7.3032,P<0.01;草地贪夜蛾1龄幼虫:Z=-8.7769,P<0.01;草地贪夜蛾2龄幼虫:Z=-8.5768,P<0.01);除对草地贪夜蛾1龄幼虫的处理时间Th无显著差异(Z=-0.1111,P=0.9115)外,黄玛草蛉2龄幼虫对猎物其他虫态的处理时间均显著长于黄玛草蛉3龄幼虫(草地贪夜蛾卵:Z=15.3689,P<0.01;草地贪夜蛾2龄幼虫:Z=5.9535,P<0.01)(表5)。随着草地贪夜蛾龄期的增长,黄玛草蛉的瞬时攻击率下降,但黄玛草蛉幼虫对草地贪夜蛾卵的瞬时攻击率均高于对1和2龄幼虫,且对卵的处理时间均低于对1和2龄幼虫。从理论日最大捕食量T/Th值来看,黄玛草蛉3龄幼虫捕食草地贪夜蛾卵及1和2龄幼虫的T/Th值依次为235.294,77.419和76.923,均高于黄玛草蛉2龄幼虫(T/Th值依次为85.106,77.170和46.242),说明前者的捕食能力强于后者。

表4 黄玛草蛉幼虫对草地贪夜蛾捕食作用的Logistic回归方程拟合的最大似然估计值Table 4 Maximum likelihood estimates from Logistic regression analysis of Spodoptera frugiperda consumed by Mallada basalis larvae

图1 不同猎物密度下黄玛草蛉2龄幼虫对草地贪夜蛾卵(A)以及1龄(B)与2龄(C)幼虫的捕食量Fig.1 Numbers of eggs (A),and the 1st (B) and 2nd (C) instar larvae of Spodoptera frugiperda consumed by the 2nd instar larvae of Mallada basalis at different prey densities图中曲线是由Rogers随机捕食者模型方程拟合所得,圆点表示观察值;图2同。Curves were fitted by using Rogers’ random predator equation,the dots indicating the observed values.The same for Fig.2.
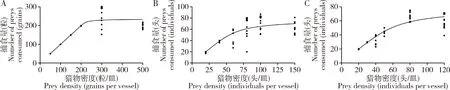
图2 不同猎物密度下黄玛草蛉3龄幼虫对草地贪夜蛾卵(A)以及1龄(B)与2龄(C) 幼虫的捕食量Fig.2 Numbers of eggs (A),and the 1st (B) and 2nd (C) instar larvae of Spodoptera frugiperda consumed by the 3rd instar larvae of Mallada basalis at different prey densities

表5 黄玛草蛉幼虫对草地贪夜蛾的捕食功能反应Table 5 Predatory functional responses of Mallada basalis on Spodoptera frugiperda
3 结论与讨论
功能反应是评价天敌捕食能力的经典方法,通常无脊椎动物(例如昆虫)对猎物的捕食率随猎物密度的升高而逐步下降,呈负加速现象,表现为Ⅱ型功能反应;而脊椎动物的捕食率通常呈先上升后下降的趋势,表现为Ⅲ型功能反应(Holling,1959;徐汝梅,1987)。本研究结果表明,黄玛草蛉2和3龄幼虫对草地贪夜蛾的卵及1和2龄幼虫的捕食行为均表现为Ⅱ型功能反应。类似的结果在黄玛草蛉对其他害虫捕食能力的研究中也有报道,如黄玛草蛉1,2和3龄幼虫均对米蛾卵表现为Ⅱ型功能反应(李水泉等,2012),其2和3龄幼虫对埃及吹绵蚧若虫(叶静文等,2013)、玉米蚜和甘蓝蚜(周娟等,2020)、木瓜粉蚧若虫和雌成虫(李子园等,2021)以及扶桑绵粉蚧Phenacoccussolenopsis(Zhouetal.,2021)亦表现为Ⅱ型功能反应。而其他草蛉则表现出不同的捕食功能反应类型,例如普通草蛉Chrysoperlacarnea1龄幼虫对木瓜粉蚧Paracoccusmarginatus表现为Ⅱ型功能反应,而2和3龄幼虫则表现为Ⅲ型功能反应(Sajjadetal.,2018);适通草蛉Chrysoperlacongrua3龄幼虫对棉铃虫Helicoverpaarmigera卵表现为Ⅲ型功能反应,而亚非玛草蛉Malladadesjardinsi对同样的猎物则表现为Ⅱ型功能反应(Kabissaetal.,1996)。Hassell等(1977)认为除Ⅱ型功能反应外,Ⅲ型功能反应也普遍存在于昆虫各类群中,以上研究结果表明捕食者的功能反应类型与物种种类及其龄期有关。
从黄玛草蛉2和3龄幼虫对草地贪夜蛾各虫态的捕食量来看,黄玛草蛉3龄幼虫对草地贪夜蛾卵及1和2龄幼虫的实际最大捕食量分别为246.80粒、70.30头以及67.50头,而黄玛草蛉2龄幼虫的捕食量分别为81.80粒、53.20头以及32.30头。从反映捕食能力指标理论日最大捕食量来看,黄玛草蛉3龄幼虫也明显高于2龄幼虫,说明前者的捕食能力强于后者。这与黄玛草蛉捕食木瓜粉蚧(李子园等,2019)与扶桑绵粉蚧(Zhouetal.,2021)时的情况相似,类似的结果也见于大草蛉捕食豆大蓟马Megalurothripsusitatus以及豆蚜Aphiscraccivora(唐良德等,2017)、中华通草蛉Chrysoperlasinica捕食黑刺粉虱Aleurocanspinfetus(唐天成等,2018)等。从同一龄期黄玛草蛉幼虫对相同密度草地贪夜蛾1和2龄幼虫的捕食率来看,在较低猎物密度时(如20或40头/皿),对草地贪夜蛾不同龄期幼虫的捕食率没有明显差异,但随着猎物密度的升高,对草地贪夜蛾1龄幼虫的捕食率明显高于2龄幼虫,说明其对低龄的猎物具有更强的捕食能力,这与大草蛉(李萍等,2020)及中华通草蛉(黄海艺等,2020)的情况类似。与其他种类草蛉捕食草地贪夜蛾卵及低龄幼虫(其中实验密度有所差异)的情况相比,在高密度猎物条件下,除黄玛草蛉2和3龄幼虫对草地贪夜蛾1龄幼虫的捕食量略低于大草蛉外,其他相同龄期条件下黄玛草蛉对同一龄期草地贪夜蛾的捕食量均高于大草蛉、丽草蛉以及中华通草蛉(表6)。总的来说,黄玛草蛉幼虫对草地贪夜蛾卵及低龄幼虫展示出良好的捕食能力。

表6 4种草蛉对草地贪夜蛾卵及低龄幼虫的捕食量Table 6 Numbers of the eggs and early instar larvae of Spodoptera frugiperda consumed by four lacewing species
本研究是在室内采用培养皿及设定的猎物密度进行的,这种在局部空间内集中提供猎物的实验方式可能会放大捕食者的捕食能力。而在自然条件下,因猎物常常分散在寄主(植株)的多个部位,捕食者搜寻猎物的范围、时间会增加,这就导致了捕食者的捕食能力或单位时间内捕食量的下降,常与室内进行捕食功能反应的研究结果有较大区别(Mahzoumetal.,2020)。除此之外,天敌在田间的捕食能力还受到多种环境因素的影响,例如当田间目标害虫的密度很高时,捕食者种群扩散的概率会降低,增加了其种群内部及多种捕食者集团内的捕食竞争,从而降低了捕食者个体的捕食效能(Lester and Harmsen,2002)。另外,田间应用的复杂性还在于猎物种类的多样性,众多种类猎物的存在可能会降低天敌对目标害虫的控制作用(Benhadi-Marínetal.,2019),但却可能有利于天敌的持续生存(Messelinketal.,2016)。由于单一的捕食者、高密度的猎物以及稳定的环境条件,在室内开展捕食功能反应实验存在一定的局限性,有研究人员曾对实验条件进行优化,从而更科学合理地评价天敌的捕食能力,例如通过增加供试天敌(王甦等,2012;Messelinketal.,2016)及猎物的种类(Messelinketal.,2016;Benhadi-Marínetal.,2019)、添加猎物的寄主植物(Desurmont and Weston,2008;张燕南等,2018)、设置不同的温度或湿度(Zhuetal.,2019)等以模拟田间环境。进一步的研究将评价猎物种类的多样性、天敌的种间与种内竞争、温度与湿度及光周期、作物类型及生育期等多个生物与非生物因素对黄玛草蛉实际控害效果的影响,以期为黄玛草蛉的田间应用提供更为全面的理论依据。
猜你喜欢
湖南城市学院学报(自然科学版)(2022年6期)2022-11-14
数学物理学报(2021年6期)2021-12-21
南通职业大学学报(2021年3期)2021-11-17
云南农业(2021年11期)2021-11-12
百科探秘·海底世界(2020年11期)2020-12-31
今日农业(2020年13期)2020-12-15
矿业科学学报(2020年5期)2020-09-18
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
太原师范学院学报(自然科学版)(2018年2期)2018-08-17
