
灰葡萄孢致病基因BcPDR1与Gα亚基基因的关系研究
2022-11-21 12:09:42魏雅迪曹丽萍刘英姿刘晓颖曹宏哲邢继红董金皋
河北农业大学学报 2022年5期
魏雅迪,曹丽萍,刘英姿,刘晓颖,曹宏哲,张 康,邢继红,董金皋
(1. 河北农业大学 真菌毒素与植物分子病理学实验室/河北省植物生理与分子病理学重点实验室,河北 保定 071000;2. 黑龙江省农业科学院 齐齐哈尔分院,黑龙江 齐齐哈尔 161000;3. 河北省承德市园林管理中心,河北 承德 067000)
灰葡萄孢(Botrytis cinerea)是一种引起植物灰霉病的重要病原真菌,它可以侵染蔬菜、水果等上千种寄主植物[1]。灰葡萄孢已成为研究植物病原真菌的模式病原菌,其生长发育和致病相关基因的功能与机制研究,不仅为深入研究灰葡萄孢生长发育和致病力的分子机理奠定基础,也为其他植物病原真菌的相关研究提供重要参考[2]。
异三聚体G 蛋白是普遍存在于真核生物中,参与多种重要的生理过程[3]。丝状真菌G 蛋白于1993 年首次报道,已明确其在生长、无性和有性发育中是必不可少的[4];其中,G 蛋白α 亚基参与了营养感知、信息素反应以及致病等过程的调控[5]。Gα亚基在稻瘟病菌(Magnaporthe grusea)的生长、繁殖以及致病过程中都具有重要的作用[6]。灰葡萄孢Gα亚基位于cAMP 信号通路上游,参与病菌的菌丝生长、分生孢子发育、分生孢子萌发以及致病过程[7]。Gα亚基基因BcBCG1调控病菌的菌落形态和侵染能力,BcBCG2、BcBCG3基因的突变体可以对寄主进行侵染,但是与野生型相比侵染速度缓慢[8-9],且BcBCG3基因的突变体分生孢子的发育和分生孢子萌发均具有特异性缺陷[6]。本实验室前期研究利用RNAi 技术获得了灰葡萄孢BcBCG2和BcBCG3基因的RNAi 沉默突变体,通过对突变体的表型和致病力进行分析,明确了BcBCG2和BcBCG3正调控病菌的菌丝生长、分生孢子的产量、致病力和穿透能力[10]。
灰葡萄孢BcPDR1基因为本实验室前期研究获得,利用基因敲除和互补回复技术明确了BcPDR1基因正调控病菌的菌丝生长、分生孢子和菌核的发育、菌丝侵染结构的形成以及致病力[11-12]。本研究利用qRT-PCR 技术分析BcPDR1基因突变对BcBCG2、BcBCG3基因表达的影响,以及BcBCG2、BcBCG3基因突变对BcPDR1基因表达的影响,确定BcPDR1基因与Gα 亚基基因之间的调控关系;在BcPDR1基因敲除突变体ΔBcpdr1的背景下,构建Gα 亚基基因中过表达菌株ΔBcpdr1/BcBcBCG2-OE 和ΔBcpdr1/BcBCG3-OE, 通 过 分析过表达菌株的表型和致病力,明确BcPDR1基因与BcBCG2、BcBCG3基因之间的关系,从而确定BcPDR1基因与病菌cAMP 信号通路之间的关系,为阐明灰葡萄孢BcPDR1基因调控病菌生长发育和致病力的分子机制提供依据。
1 材料和方法
1.1 试验材料
灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA 插入突变体BCt89、敲除突变体ΔBcpdr1和 回 复 菌 株CE[13],BcBCG2、BcBCG3基 因 的RNAi 菌株,均由河北省植物生理与分子病理学重点实验室/河北农业大学真菌毒素与植物分子病理学实验室保存及提供。
1.2 qRT-PCR 分析
利用qRT-PCR 技术,分析灰葡萄孢野生型菌株BC22,BcPDR1基因的缺失突变体BCt89和ΔBcpdr1和互补回复菌株CE 中的BcBCG2、BcBCG3基因的表达水平以及BcBCG2、BcBCG3基因的RNAi 沉默突变体中BcPDR1基因的表达水平。以灰葡萄孢Tubulin基因为内参基因,用BcBCG2、BcBCG3和BcPDR1基因的特异性引物(见表1),以各菌株的cDNA 为模板,进行qRT-PCR 分析。
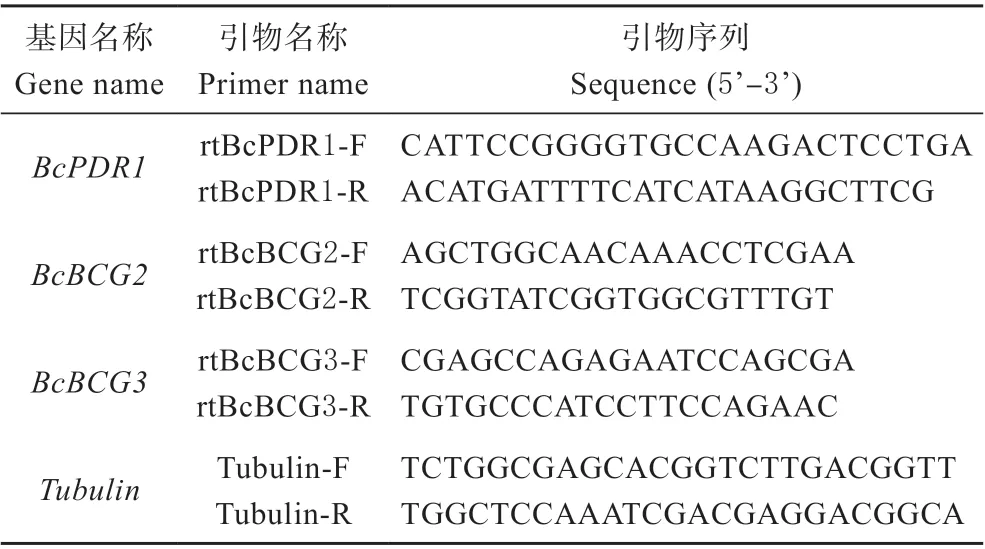
表1 qRT-PCR 引物设计Table 1 Primers for qRT-PCR
1.3 Gα 亚基基因过表达菌株的构建
1.3.1 Gα亚基基因过表达载体的构建 用BcBCG2、
BcBCG3基因的特异性引物和灰葡萄孢野生型BC22菌株的cDNA 为模板,PCR 扩增BcBCG2、BcBCG3基因的编码区序列,电泳分离、纯化后进行克隆、测序。将测序正确的BcBCG2、BcBCG3基因序列与基因过表达终载体pEarly gate 103 连接,菌落PCR 和测序鉴定后,获得BcBCG2、BcBCG3基因的过表达载体pEarly gate 103-BcBCG2、pEarly gate 103-BcBCG3。
1.3.2 Gα 亚基基因过表达转化子的获得与鉴定 利用农杆菌介导的转化方法[14],将pEarly gate 103-BcBCG2、pEarly gate 103-BcBCG3分 别 转化BcPDR1基因敲除突变体ΔBcpdr1中,获得转化子后在含有抗性的培养基中进行多次筛选,然后利用PCR 和qRT-PCR 技术分别从DNA 水平和转录水平对转化子进行鉴定。
1.4 Gα 亚基基因过表达菌株的表型分析
将灰葡萄孢野生型BC22 和突变体ΔBcpdr1、ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BcBCG3-OE 分 别接种在PDA 培养基上,20 ℃黑暗条件下培养7 d,对其菌落形态、菌核产生等表型进行观察;同时,收集各菌株的分生孢子,用血球计数板统计各菌株的分生孢子数量;制备各菌株的菌丝悬浮液,显微镜下观察其菌丝形态;分别将各菌株的菌盘(Φ=8.0 mm)接种在新的PDA 培养基上,20 ℃黑暗培养,每天定时测定各菌株的菌落直径,进而分析菌株的生长速率[15]
1.5 Gα 亚基基因过表达菌株的致病力分析
将灰葡萄孢野生型BC22 和突变体ΔBcpdr1、ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BcBCG3-OE 定量(菌盘Φ=8.0 mm)接种到离体的番茄果实上,检测各菌株的致病力,测量其病斑面积。
2 结果与分析
2.1BcPDR1基因对Gα 亚基基因表达的影响
利用qRT-PCR 技术,分析BcPDR1基因突变体Gα亚基基因BcBCG2、BcBCG3的表达情况。结果发现,BcPDR1基因缺失突变体BCt89 和ΔBcpdr1中BcBCG2、BcBCG3基因的表达水平显著高于野生菌株BC22 和回复菌株CE(见图1)。表明BcPDR1基因缺失影响了BcBCG2、BcBCG3基因的表达,也说明BcPDR1基因负调控BcBCG2、BcBCG3基因的表达。

图1BcPDR1基因突变体中BcBCG2和BcBCG3基因的表达分析Fig.1 Expression levels ofBcBCG2andBcBCG3inBcPDR1mutants
2.2 Gα 亚基基因对BcPDR1基因表达的影响
利用qRT-PCR 技术,分析Gα亚基基因BcBCG2、BcBCG3的突变对BcPDR1基因表达的影响。结果发现,BcBCG2、BcBCG3的RNAi 沉默突变体中BcPDR1基因的表达水平显著低于野生型菌株BC22(图2)。表明BcBCG2、BcBCG3基因突变影响了BcPDR1基因的表达,也说明BcBCG2、BcBCG3基因正调控BcPDR1基因的表达。

图2BcBCG2和BcBCG3基因突变体中BcPDR1基因的表达分析Fig. 2 Expression levels of theBcPDR1in RNAi mutants ofBcBCG2andBcBCG3genes
2.3BcBCG2、BcBCG3过表达菌株的构建
利用Gateway 技术,成功构建了BcBCG2和BCBCG3基因的过表达载体。利用农杆菌介导的遗传转化方法,将目的基因表达载体转入到突变体ΔBcpdr1中,通过抗性筛选获得转化子,在对转化子进行PCR 验证后进行qRT-PCR 分析。结果发现,BcBCG2、BcBCG3基因的转化子中BcBCG2和BcBCG3基因的表达水平显著高于突变体ΔBcpdr1(图3)。表明了BcBCG2和BCBCG3基因的过表达菌株ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BCBCG3-OE 构建成功。

图3BcBCG2和BcBCG3过表达菌株的基因表达分析Fig.3 Gene expression analysis inBcBCG2andBcBCG3overexpressing strains
2.4BcBCG2、BcBCG3过表达菌株的表型分析
对ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BCBCG3-OE 菌株的菌落形态、菌丝形态进行观察(见图4),发现过表达菌株ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BCBCG3-OE 的菌落灰褐色,产生大量菌核,菌丝灰黑色、分隔明显,与野生型BC22 的表型非常一致,而显著区别于BcPDR1基因的敲除突变体ΔBcpdr1。对过表达菌株的菌丝生长速率和分生孢子产量进行分析,发现ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BCBCG3-OE 菌株的生长速率和分生孢子数量均与野生型较为接近,其生长速率与野生型没有明显差别,其产孢量略低于野生型;过表达菌株的生长速率和分生孢子数量均显著区别于突变体ΔBcpdr1。表明BcBCG2、BcBCG3基因的过表达使突变体ΔBcpdr1的表型得到了恢复,也说明BcPDR1基因与BcBCG2、BcBCG3基因之间密切相关。

图4BcBCG2和BcBCG3过表达菌株的表型分析Fig. 4 Phenotypic analysis of over-expression isolates ofBcBCG2andBcBCG3
2.5BcBCG2、BcBCG3过表达菌株的致病力分析
以灰葡萄孢野生型BC22菌株和BcPDR1基因的敲除突变体ΔBcpdr1为对照,对ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BcBCG3-OE 菌 株 的 致 病力进行分析,发现接种野生型BC22 和过表达菌株的部位均能产生明显的病斑,且接种过表达菌株的病斑面积与野生型基本一致,而接种突变体ΔBcpdr1的部位并未产生明显的病斑(图5)。表明BcBCG2、BCBCG3基因的过表达均能使突变体ΔBcpdr1的致病力得到了恢复,进一步说明了BcPDR1基因与BcBCG2、BcBCG3基因之间密切相关。

图5BcBCG2和BcBCG3过表达菌株的致病力Fig. 5 Pathogenicity analysis of over-expression isolates ofBcBCG2andBcBCG3
3 结论与讨论
灰葡萄孢cAMP 信号途径在病菌的生长、发育和致病过程中起着重要作用。异源三聚体G-蛋白Gα 亚基由BcBCG1[7]、BcBCG2[16]、BcBCG3[17]基因编码,位于灰葡萄孢cAMP 信号途径上游,参与调控病菌的形态建成、分生孢子萌发和侵入生长[18]。其中,BcBCG1基因的缺失突变体ΔBcBCG1不能产生2 次扩展病斑,外源cAMP 的添加可以使ΔBcBCG1突变体的菌落形态恢复到野生型;BcBCG2、BcBCG3基因的缺失突变体ΔBcBCG2、ΔBcBCG3可以侵染寄主并能继续扩展,但是扩展速度较野生型缓慢[8-9]。ΔBcBCG3可以使分生孢子萌发受损并引起延迟发病。实验前期研究明确了灰葡萄孢BcPDR1参与病菌的生长、发育和致病力的调控[18],但是灰葡萄孢BcPDR1与cAMP信号途径之间的关系并未明确[20]。因此,本研究首先利用qRT-PCR 技术,分析了BcPDR1突变体中BcBCG2、BcBCG3基因表达水平和BcBCG2和BcBCG3突变体中BcPDR1基因表达水平,结果确定了BcPDR1基因负调控BcBCG2、BcBCG3基因的表达,BcBCG2、BcBCG3基因正调控BcPDR1的表达,表明了BcPDR1基因与病菌的cAMP 信号途径密切相关。为进一步明确BcPDR1基因与BcBCG2和BcBCG3基因之间的关系,试验成功构建了BcBCG2和BCBCG3基因的过表达菌株ΔBcpdr1/BcBCG2-OE、ΔBcpdr1/BcBCG3-OE,然后对过表达菌株进行表型和致病力分析,确定了BcBCG2和BcBCG3基因的过表达能够使BcPDR1基因的敲除突变体ΔBcpdr1的表型和致病力得到恢复,表明了BcPDR1基因与BcBCG2和BcBCG3基因之间密切相关,同时说明了BcPDR1基因与病菌cAMP 信号途径之间具有相关性。但是BcPDR1基因与病菌cAMP 信号途径之间的调控关系及机制尚需深入研究,为阐明灰葡萄孢生长发育和致病力调控的分子机理奠定基础。
猜你喜欢
中国森林病虫(2021年6期)2021-12-20 08:45:42
世界最新医学信息文摘(2020年68期)2020-12-25 11:55:27
中国森林病虫(2018年4期)2018-09-19 12:13:08
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:13
塔里木大学学报(2015年1期)2015-04-25 02:38:44
山东医药(2015年40期)2015-02-28 14:28:45
大豆科技(2014年5期)2014-03-23 02:46:18
中国医学科学院学报(2013年6期)2013-03-11 20:26:01
