
环境因子和储藏时期对长枝木霉PT6蛋白培养滤液杀线活性评价
2022-11-18 03:53刘永红毛维兴陈瑞莲马耀杰张树武徐秉良
草业科学 2022年10期
刘永红,毛维兴,陈瑞莲,马耀杰,张树武,徐秉良
(甘肃农业大学植物保护学院 / 甘肃省农作物病虫害生物防治工程实验室, 甘肃 兰州 730070)
禾谷孢囊线虫(Heterodera avenae)自1908 年在英国首次发现后,目前已在印度、中国和澳大利亚等40 多个国家均有发生[1]。据估计,全世界每年因禾谷孢囊线虫病造成的经济损失达数百亿美元[2]。我国华北、华中、华东和西北地区16 个省(市)小麦(Triticum aestivum)产区均有禾谷孢囊线虫病发生,危害小麦面积约400 万hm2[3]。已有研究表明,禾谷孢囊线虫除危害小麦、大麦(Hordeum vulgare)、燕麦(Avena sativa)和青稞(Hordeum vulgare)外[4],还对禾本科牧草黑麦草(Lolium perenne)、鹅观草(Roegneria kanoji)、苇状羊茅(Festuca elatior)、球茎草芦(Phalaris tuberosa)和鸭茅(Dactylis glomerata)具有危害[5]。特别是禾本科牧草作为禾谷孢囊线虫的中间寄主或桥梁寄主存在于麦类作物田间,使得在农牧交错带对禾谷孢囊线虫的防治提出了更高的要求[6]。目前,尽管化学杀线剂在防治禾谷孢囊线虫方面取得了良好的效果,但存在对人畜有害、污染环境以及影响土壤中有益微生物等缺陷[7]。因此,生物防治禾谷孢囊线虫受到了人们的广泛关注[8]。
木霉菌(Trichodermaspp.) 作为重要的农业微生物资源,不仅在植物病原菌和植物寄生线虫的生物防治中发挥重要的作用,而且还在促进植物生长、诱导植物产生抗病性等方面具有显著的效果[9]。古丽君等[10]研究发现,长枝木霉(Trichoderma longibrachiatum)孢子悬浮液对6 种牧草种子的发芽率,幼苗根长、芽长及干重均有明显的促进作用且能够诱导和提高其抗性。章武等[11]研究发现4 株木霉菌对14 种草坪草病原菌具有不同程度的抑制效果,最高抑制效果可达100%,表明木霉菌在牧草病害防治中具有重要作用。
植物寄生线虫体壁是由真皮层分泌产生的非细胞结构,由角蛋白、胶原蛋白和纤维类反向交联组成,主要成分是蛋白质;线虫的卵壳由蛋白质和几丁质层组成,其主要成分是几丁质;体壁和卵壳是线虫阻止生防真菌侵染的重要屏障。因此,蛋白酶和几丁质酶等水解酶类在真菌毒杀线虫过程中发挥着重要作用[12]。有研究表明,深绿木霉(T.atroviride)、哈茨木霉(T. harzianum)发酵滤液均能显著降低南方根结线虫(Meloidogyne incognita)卵孵化率,对2 龄幼虫具有较高的致死作用,同时其发酵液对线虫卵和幼虫体壁有消解作用[13]。另外,木霉菌不仅可以寄生线虫卵和幼虫,而且可以产生大量具有杀线活性的胞外酶,主要有丝氨酸蛋白酶、几丁质酶和蛋白酶等,对植物寄生线虫有较好毒杀效果[14]。Sharon 等[15]报道哈茨木霉菌高效表达丝氨酸蛋白酶基因prb1的转化子对南方根结线虫(M. incognita)毒杀效果有增强作用,表明蛋白酶是木霉菌防治植物线虫过程中的毒力因子。Chen 等[16]从拟康氏木霉(T. pseudokoningii) SMF2中分离到1 个线虫体壁降解丝氨酸蛋白酶SprT,该酶可以抑制根结线虫卵的孵化并且对2 龄幼虫也具有较高的致死率。目前关于蛋白酶和几丁质酶等水解酶类对植物寄生性线虫的防治机制及杀线活性已有相关研究。然而,有关木霉杀线蛋白稳定性研究鲜有报道。
前期植物病毒学和分子生物学课题组研究发现,长枝木霉T6 菌株孢子悬浮液不仅对小麦[17]和多种禾本科牧草具有促生作用[10],而且还可使禾谷孢囊线虫孢囊、卵和2 龄幼虫体壁降解,严重抑制孢囊和卵的孵化,导致2 龄幼虫死亡[8],但有关其胞外蛋白酶对2 龄幼虫致死活性,以及其胞外蛋白酶在运输、储藏、保存和施用过程中是否会受到外界环境条件(高温、过酸过碱、金属离子和氧化还原剂等)的干扰等方面仍尚不清楚。鉴于此,本试验以提取的PT6 蛋白培养滤液和禾谷孢囊线虫2 龄幼虫作为研究对象,通过评价不同条件处理下PT6 蛋白培养滤液对禾谷孢囊线虫2 龄幼虫的致死活性,以期明确PT6 蛋白培养滤液的杀线稳定性,为杀线蛋白制剂能够应用于生产实践提供一定的理论基础和技术支撑。
1 材料与方法
1.1 试验材料
供试蛋白滤液:PT6 蛋白培养滤液提取和保存于甘肃农业大学植物保护学院植物病毒学和分子生物学实验室。
供试线虫:由甘肃农业大学植物保护学院植物病毒学和分子生物学实验室提供。
供试药品:酪蛋白、几丁质、葡萄糖、甘露醇和卵黄蛋白等为国产分析纯。
1.2 试验方法
1.2.1 禾谷孢囊线虫2 龄幼虫分离
参考张树武等[8]方法。采用“漂浮法”分离孢囊。将孢囊收集后消毒处理置于线虫分离器内,低温(4 ℃) 刺激1 个月,然后置于培养箱中培养3 周左右(15 ℃) 获得禾谷孢囊线虫2 龄幼虫。最后收集2 龄线虫经无菌水配制成200 条·mL-1的线虫悬浮液,4 ℃保存备用。
1.2.2 pH 对PT6 蛋白培养滤液杀线活性影响
pH 用氢氧化钠溶液和盐酸溶液调节,分别将PT6 蛋白培养滤液pH 调为2、4、6、8、10 和12,并置于4 ℃下静置24 h 后重新调至中性。然后测定其杀线活性,并以原始PT6 蛋白培养滤液(pH 为7)作为对照,每个处理和对照均重复3 次。杀线活性测定参考张树武等[8]方法。利用96 孔细胞培养板进行试验,在处理和对照孔加入10 μL 线虫悬浮液和90 μL PT6 蛋白培养滤液,每个处理和对照均为16 个重复。然后,25 ℃恒温培养箱黑暗培养,统计24、48、72 h 处理后线虫死亡数并计算其死亡率。
1.2.3 温度对PT6 蛋白培养滤液杀线活性影响
将提取保存的PT6 蛋白培养滤液分别置于30、40、50、60、70、80、90 和100 ℃恒温水浴锅处理30 min,然后置于冰上使其迅速冷却,试验以未经处理的PT6 蛋白培养滤液为对照,3 次重复。室温冷却后参考 1.2.2 方法测定其杀线活性。
1.2.4 金属离子对PT6 蛋白培养滤液杀线活性影响
利 用 无 菌 水 配 制0.1 mol·L-1的K+、Na+、Cu2+、Zn2+、Mn2+、Fe3+、Mg2+离子溶液,并分别与PT6 蛋白培养滤液按1 ∶ 9 的体积比混合,以单一PT6 培养滤液作为对照,每个处理和对照均重复3 次。待充分混合后,分别置于4 ℃下放置24 h 后参考1.2.2 方法测定其杀线活性。
1.2.5 氧化还原剂对PT6 蛋白培养滤液杀线活性影响
氧化剂和还原剂溶液使用双氧水与亚硫酸钠配置,氧化剂和还原剂浓度配制成0.01、0.02、0.04、0.06、0.08 和0.1 mol·L-1。然后将不同浓度氧化剂和还原剂分别与PT6 蛋白培养滤液按1 ∶ 9 体积比混合。待充分混合后,4 ℃静置24 h 后参考1.2.2 方法测定其杀线活性。以未添加氧化剂和还原剂的培养滤液作为对照,每个处理和对照均重复3 次。
1.2.6 PT6 蛋白培养滤液储藏稳定性测定
将提取的PT6 蛋白培养滤液 (50 mL) 置于无菌三角瓶并室温下避光保存,并分别于保存10、30、60、90 d 后参照1.2.2 方法测定其杀线活性。试验以新制备的PT6 蛋白培养滤液作为对照,每个对照和处理均重复3 次。
1.3 数据处理
采用Excel 2016 进行数据统计分析并作图,SPSS 19.0 软件进行方差分析和Duncan 新复极差法用以多重比较(P< 0.05)。
2 结果与分析
2.1 pH 和温度对PT6 蛋白培养滤液杀线活性的影响
在pH 4~10 时,72 h 线虫死亡率均在80%以上,与对照相比无显著差异(P> 0.05)。pH 超过10时,PT6 蛋白培养滤液杀线活性开始降低。pH 为2时,72 h 线虫死亡率为69.10%;pH 为12 时,72 h 线虫死亡率仅为66.55%,均显著低于对照(P< 0.05) (图1)。

图1 pH 对PT6 蛋白培养滤液杀线活性的影响Figure 1 Effect of pH on the nematicidal activity of PT6 protein culture filtrate
温度在30~60 ℃时,PT6 蛋白培养滤液对禾谷孢囊线虫2 龄幼虫毒杀作用和对照相比无显著差异(P> 0.05)。当温度超过60 ℃,其对线虫杀线活性开始降低。与对照相比,70~100 ℃温度下线虫死亡率显著降低(P< 0.05)。温度为100 ℃时,24、48、72 h 线虫死亡率仅为17.50%、24.64%和28.51% (图2)。

图2 温度对 PT6 蛋白培养滤液杀线活性的影响Figure 2 Effect of temperature on the nematicidal activity of PT6 protein culture filtrate
2.2 金属离子和氧化还原剂对PT6 蛋白培养滤液杀线活性的影响
不同金属离子对PT6 蛋白培养滤液杀线活性影响不同,Zn2+和K+对PT6 蛋白培养滤液杀线活性具有明显的增强作用(图3)。与对照相比,Zn2+和K+处理线虫48、72 h 后线虫死亡率显著提高(P< 0.05),72 h其处理线虫死亡率高达91.45%和91.28%,而Cu2+对PT6 蛋白培养滤液杀线活性具有显著抑制作用(P<0.05),处理72 h 时线虫死亡率仅49.36%;Fe3+、Mg2+、Mn2+和Na+处理48 h 和72 h 对PT6 蛋白培养滤液杀线活性与对照比无显著影响(P> 0.05),处理72 h后线虫死亡率分别为80.62%、82.03%、81.26%和82.68%,具有良好稳定性。

图3 金属离子对PT6 蛋白培养滤液杀线活性的影响Figure 3 Effect of metal ions on the nematicidal activity of PT6 protein culture filtrate
较高浓度氧化剂可使PT6 蛋白培养滤液杀线活性降低,而较低浓度氧化剂对PT6 蛋白培养滤液杀线活性无显著影响(P> 0.05) (图4)。氧化剂浓度低于0.10 mol·L-1时,线虫死亡率与对照相比差异不显著(P> 0.05);氧化剂浓度为0.10 mol·L-1时,PT6 蛋白培养滤液杀线活性显著降低(P< 0.05),24、48 和72 h处理后,线虫死亡率分别为41.37%、53.42%和64.29%。

图4 氧化剂对 PT6 蛋白培养滤液杀线活性的影响Figure 4 Effect of oxidant on the nematicidal activity of PT6 protein culture filtrate
与对照相比,当还原剂浓度较低时对PT6 蛋白培养滤液杀线活性无显著影响(P> 0.05),而还原剂浓度较高时会使PT6 蛋白培养滤液杀线活性显著降低(P< 0.05) (图5)。当还原剂浓度低于0.08 mol·L-1时,线虫死亡率与对照均无显著差异(P> 0.05);当还原剂浓度为0.08 和0.10 mol·L-1时,线虫72 h 死亡率显著低于对照(P< 0.05),72 h 后死亡率分别为74.62%和72.96%。
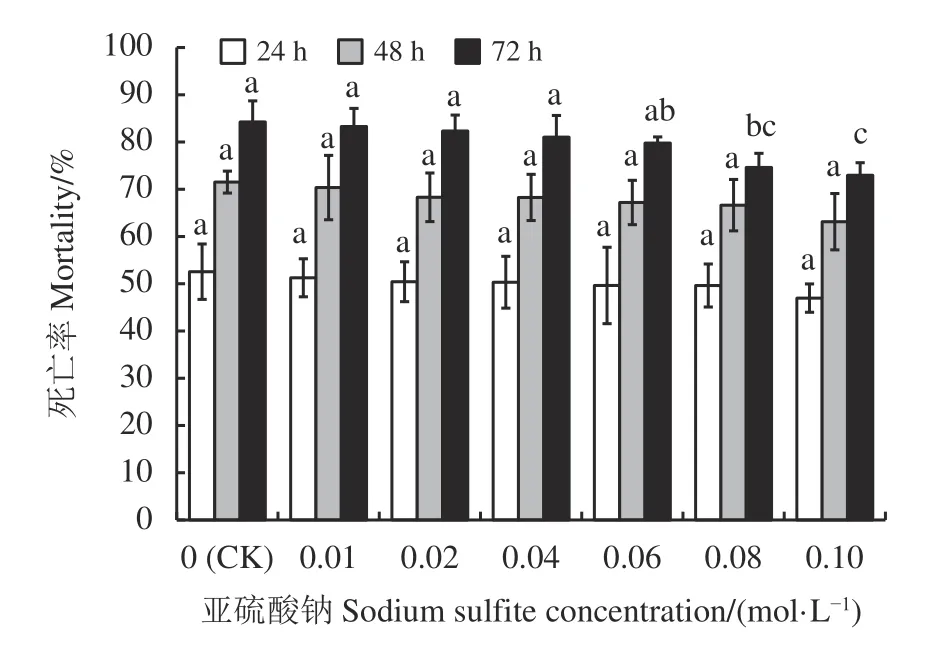
图5 还原剂对 PT6 蛋白培养滤液杀线活性的影响Figure 5 Effect of reducing agent on the nematicidal activity of PT6 protein culture filtrate
2.3 储藏时期对PT6 蛋白培养滤液杀线活性的影响
与对照相比,PT6 蛋白培养滤液储藏时期为10~60 d 时,对蛋白培养滤液杀线活性无显著影响(P> 0.05),72 h 死亡率都在80%以上。但在处理90 d时,PT6 蛋白培养滤液处理线虫48 和72 h 死亡率均显著低于对照(P< 0.05),分别为64.81% 和77.24%(图6)。

图6 储藏时期对PT6 蛋白培养滤液杀线活性的影响Figure 6 Effect of storage time on the nematicidal activity of PT6 protein culture filtrate
3 讨论与结论
Hran 等[18]发现木霉菌的杀线活性蛋白易受外界环境条件的干扰,常具有不稳定性。本研究表明长枝木霉PT6 蛋白培养滤液对禾谷孢囊线虫2 龄幼虫具有较强的致死作用,当pH 4~10,温度30~60 ℃,0.1 mol·L-1的Fe3+、Mg2+、Mn2+和Na+,氧化剂浓度低于0.1 mol·L-1、还 原 剂 低 于0.08 mol·L-1时,PT6 蛋白培养滤液对其杀线活性无显著影响,并且储藏0~60 d 均具有良好的稳定性,Zn2+和K+对其杀线活性还具有一定的增效作用。同时,本研究也表明,长枝木霉PT6 蛋白培养滤液pH 8、温度30 ℃、0.1 mol·L-1Zn2+处理后对禾谷孢囊线虫的毒杀效果最好。
胞外蛋白的温度和酸碱稳定性对于其商业应用有重要的作用[19]。张晓云等[20]研究表明,枯草芽孢杆菌CAB-1 抑菌蛋白在100 ℃下处理30 min 活性变化差异不显著,而121 ℃处理30 min 对番茄灰霉病菌灰葡萄孢(Botrytis cinerea)抑菌活性与对照相比下降了72%,在pH 3~12 时抑菌活性均大于75%。本研究结果表明,长枝木霉PT6 蛋白培养滤液在pH 4~10、温度30~60 ℃时对禾谷孢囊线虫2 龄幼虫均有较高的毒杀作用,但过酸过碱和高温条件会使其杀线活性降低,这可能是因为蛋白结构被破坏,导致蛋白变性、钝化失活。Dunaevsky 等[21]研究发现哈茨木霉分泌的胰蛋白酶类丝氨酸蛋白酶在pH 6.0~11.0、温度45~50 ℃保持初始酶活性不变;Aissaoui 等[22]从哈茨木霉中分离纯化的丝氨酸蛋白酶在pH 5.0~10.0 均能保持初始酶活性的62%以上,在温度30~50 ℃仍具有超过85%的剩余酶活性,与本研究结果较为相似。金属离子也是影响蛋白酶活性的因素之一,胡燕梅等[23]研究表明Mg2+、Fe2+、Ca2+、Ba2+和Mn2+等对盾壳霉几丁质酶活性有激活作用,而Zn2+和Cu2+却对盾壳霉几丁质酶活性发挥有抑制作用。本研究表明Zn2+和K+对PT6 蛋白培养滤液的杀线活性有促进作用和Cu2+对蛋白培养滤液杀线活性具有抑制作用,Mg2+、Fe3+、Mn2+、Na+对PT6 蛋白培养滤液杀线活性无显著影响,其原因可能是与杀线蛋白结合的特殊受体也能与某些金属离子相结合,而金属离子和受体结合会减少受体与杀线蛋白的结合,最终影响杀线蛋白的杀线活性。吴晓冰等[24]发现绿色木霉(T. viride) T-YY 菌株黄色素对较高浓度的氧化剂可以适应,但对还原剂的稳定性较差,而本研究发现蛋白培养滤液在较低浓度的氧化还原剂胁迫下具有良好的杀线稳定性,但随着浓度增加其稳定性降低。褚福红和于新[25]研究结果表明氧化剂、还原剂在一定浓度范围内,绿色木霉代谢物对芒果炭疽菌(Colletotrichum gloeosporiodes)抑菌活性影响不显著,与本研究结果基本一致。张建华等[26]通过对长枝木霉T6 菌株发酵液抑菌稳定性研究表明,当发酵液在4 ℃和25 ℃环境下放置60 d,其抑菌活性和对照相比无显著差异,与本研究结果基本一致,说明PT6 蛋白滤液具有较好的储藏稳定性。
本研究表明,长枝木霉PT6 蛋白培养滤液具有较好的稳定性,如耐酸碱、高温,对金属离子不敏感,储藏效果良好等特性,但关于其他因素对PT6 蛋白培养滤液杀线稳定性的影响及不同环境因子影响PT6 蛋白培养滤液稳定性机制还有待进一步研究。
猜你喜欢
海洋与湖沼(2022年6期)2022-11-25
陕西教育·高教版(2022年6期)2022-06-17
暨南大学学报(自然科学与医学版)(2022年3期)2022-06-16
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与机械(2019年1期)2019-03-30
源流(2018年7期)2018-12-03
青春美文CUTE(2017年2期)2017-11-14
山东农业大学学报(自然科学版)(2017年3期)2017-06-24
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
海洋学报(2017年4期)2017-04-13
