
复合益生菌发酵玉米-豆粕型饲料的品质评价及抑菌性能研究
2022-11-11 06:53丁亚伟徐艳辉王雅丽王海玉郝庆红段春辉郭云霞纪守坤刘月琴张英杰
家畜生态学报 2022年11期
丁亚伟,刘 月, 徐艳辉,王雅丽,王海玉,郝庆红,4,段春辉*,郭云霞,,4*,纪守坤,严 慧,刘月琴,张英杰
(1.河北农业大学 动物科技学院,河北 保定 071001;2.河北省畜牧兽医研究所,河北 保定 071001;3.河北农业大学 生命科学学院,河北 保定 071001;4.农业微生物河北省工程中心,河北 保定 071001)
发酵饲料可降解饲料中大分子营养物质[1],利于幼龄动物肠道的消化吸收。益生菌在发酵过程中还产生有机酸和风味物质,酸化的饲料可防止饲料霉变,抑制有害菌生长,进而减弱大肠杆菌致肠毒症的能力[2],发酵过程中产生的有机酸可以激活与蛋白质和碳水化合物有关消化酶,提高饲料营养利用率[3],最终达到增强免疫力促进生长的目的[4]。芽孢杆菌是最理想的发酵菌种,可形成芽孢,属于兼性厌氧菌,生命力顽强,具有较高的蛋白酶、淀粉酶和脂肪酶活性,产生活性抑菌物质,且独特的夺氧机制为厌氧菌提供生长条件,同时还产生B族维生素和维生素C等。生产中常用的芽孢杆菌有地衣芽孢杆菌和枯草芽孢杆菌。乳酸菌因其发酵可产生乳酸提供酸性环境,可抑制有害菌的增殖,在生产中也被广泛应用。近年来发酵饲料作为优质的替抗产品在单胃动物中研究较多,在幼龄反刍动物中研究较少。本试验以玉米、豆粕、麸皮为发酵底物,以前期筛选的抗腹泻地衣芽孢杆菌(Y5-39)[5]、乳酸杆菌(RSG-1)、枯草芽孢杆菌(B-1)为发酵菌种,研究饲料经复合益生菌发酵后营养物质的变化及对腹泻病原菌的抑制作用,为益生菌发酵料的应用提供理论基础。
1 材料与方法
1.1 试验时间及地点
本试验于2021年3月在河北农业大学动物科技学院完成。
1.2 菌种及培养基
发酵菌种:枯草芽孢杆菌B-1(CGMCC22064),地衣芽孢杆菌Y5-39(CGMCC22062),乳酸杆菌RSG-1(CGMCC22061)。腹泻病原菌:大肠杆菌(E.coli)、沙门氏菌(Salmonella)和痢疾杆菌(ShigellaCastellani)。所有菌种均由河北农业大学生命科学学院制药工程系实验室分离并保存。根据Design-Expert软件中Mixture模型,设计不同菌种分组,以酸溶蛋白为响应值,最终得到乳酸杆菌、枯草芽孢杆菌和地衣芽孢杆菌1∶1∶4复合。
琼脂培养基(NA):琼脂20 g,牛肉膏3 g,氯化钠5 g,蛋白胨10 g、1 L蒸馏水,pH 7.2~7.4;发酵培养基(NB):牛肉膏3 g,氯化钠5 g,蛋白胨10 g,1 L蒸馏水,pH7.2~7.4;肉汤培养基(MRS):蛋白胨10 g、牛肉粉5 g、酵母粉5 g、葡萄糖20 g、吐温80 1 mL、磷酸氢二钾2 g、乙酸钠5 g、柠檬酸三铵2 g、硫酸镁0.2 g、硫酸锰0.05 g、琼脂粉15 g、1 L蒸馏水,pH7.2~7.4。使用前121 ℃灭菌20 min。
1.3 试验设计
本试验中饲料原料(衡水某畜牧有限公司提供)按处理方式分别为对照组和发酵组,对照组不做处理,发酵组用复合益生菌接种,接种量10%、含水量45 %,装瓶室温(30 ℃,发酵瓶容积为350~400 g)密闭发酵14 d。发酵结束后分别检测两组营养物质含量、酸溶蛋白、pH、消化酶、挥发性脂肪酸、抗原蛋白、霉菌毒素等指标,并进行抑菌试验,检测抑菌效果。
1.4 指标测定与方法
1.4.1 酸溶蛋白的测定 取烘干后对照组和试验组饲料各3 g于50 mL离心管中,加入25 mL 15%三氯乙酸静置沉淀4 h,离心取5 mL上清,后按照GB/T6432《饲料中粗蛋白测定方法》[6]测定酸溶蛋白含量。
1.4.2 常规营养成分测定 取烘干后两组的饲料各50 g样品,105 ℃烘干20 min,65 ℃烘48 h后,粉碎过60目筛,干物质(DM)、粗蛋白(CP)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、粗脂肪(EE)含量参照《饲料分析及饲料质量检测技术》[7]测定。
1.4.3 消化酶含量与抗原蛋白测定 饲料中淀粉酶采用α-淀粉酶(α-amylase,α-AL)试剂盒测定,蛋白酶采用酸性蛋白酶(Acid protease, ACP)试剂盒检测,脂肪酶采用脂肪酶(Lipase,LPS)试剂盒检测,纤维素酶采用纤维素酶(Cellulase,CL)试剂盒检测。酶含量检测试剂盒均购自索莱宝试剂公司。大豆球蛋白、β-伴大豆球蛋白采用大豆球蛋白和β-伴大豆球蛋白试剂盒(龙科方舟生物工程有限公司)测定,操作步骤参照试剂盒说明书进行。
1.4.4 毒素的测定 黄曲霉毒素B1、玉米赤霉烯酮、呕吐毒素测定方法参照GB/13078-2017《饲料卫生标准》进行测定。
1.4.5 pH与有机酸测定 样品与蒸馏水按1∶10放于锥形瓶中,37 ℃、180 r/min振荡30 min得到饲料浸提液,用pH计(PSH-3D,梅特勒-托利多)测定pH。参照孙蕊[8]等的方法,称取适量样品(1 g)于10 mL容量瓶,加流动相(0.01 mol/L磷酸二氢钾,pH=2.55)定容至刻度,超声30 min,利用高效液相色谱法(HPLC)进行样品中乳酸、乙酸、丙酸、丁酸、戊酸含量的测定,色谱柱(C18柱,250 mm×4.6 mm×5 um),流速(0.5 mL/min),柱温(35 ℃)进样量(10 μL),检测波长(210 nm)。有机酸计算公式:
式中:X为样品中有机酸的含量,mg/kg;m为样品称样量,g;V为样品定容体积,mL;C为各有机酸在标曲上计算的浓度,μg/mL。
1.4.6 微生物含量的检测 芽孢杆菌参照GB/T26428-2010《饲用微生物制剂中枯草芽孢杆菌的检测》、乳酸杆菌参照GB4789.35-2016《食品微生物学检验 乳酸菌检验》。其中芽孢杆菌计数选用NA培养基,乳酸杆菌计数选用MRS培养基。
1.4.7 抑菌试验E.coli、沙门氏菌、痢疾杆菌接种到NB中,于37 ℃、180r/min培养24 h。按1∶100(病原菌:培养基)在NA中分别接种大肠杆菌、沙门氏菌、痢疾杆菌菌悬液,于灭菌培养皿中倾注15~20 mL的NA,制成病原菌平板。取未烘干的对照组和发酵组各10 g样品,分别加入100 mL正丁醇在37 ℃,45k Hz下超声波震荡浸提1 h,3 000 r/min离心10 min,收集上清液备用,用平板对峙试验在病原菌平板上打孔,加入60 μL浸提液,每个样品9个平行,37 ℃培养箱中培养24 h,记录抑菌圈直径大小。
1.5 数据统计与分析
数据基础整理采用WPS软件,采用SPSS软件中T-text检验进行数据统计分析,所有数据均以“平均值±标准差”表示,以P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 复合益生菌发酵饲料对常规营养成分的影响
由表1可知,与对照组相比,饲料经复合益生菌发酵后粗蛋白含量极显著增加了11.27%(P<0.01),酸溶蛋白含量极显著增加了498.06%(P<0.01),干物质、粗灰分、粗脂肪、中性洗涤纤维、酸性洗涤纤维与对照组无显著差异(P>0.05)。
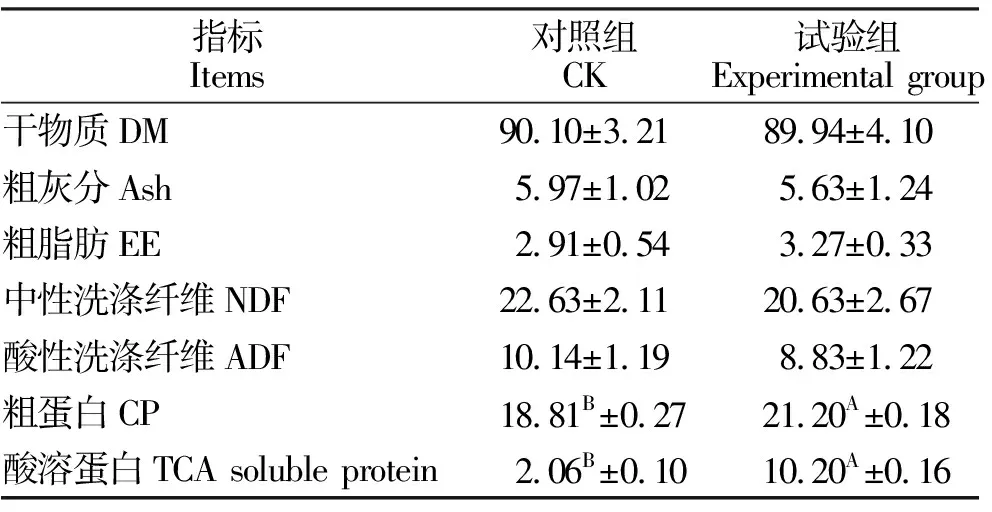
表1 饲料发酵前后营养成分分析(干物质基础)
2.2 复合益生菌发酵饲料对抗原蛋白及消化酶的影响
复合益生菌发酵饲料对抗原蛋白及消化酶的影响见表2。由表2可知,饲料经复合益生菌发酵后极显著降低了大豆抗原蛋白、β-伴大豆球蛋白含量(P<0.01),分别下降了70.85%和66.59%,淀粉酶、纤维素酶和脂肪酶极显著增加(P<0.01),酸性蛋白酶显著提高(P<0.05)。

表2 复合益生菌发酵饲料对抗原蛋白及消化酶的影响
2.3 复合益生菌发酵饲料对霉菌毒素含量的影响
复合益生菌发酵饲料对霉菌毒素含量的影响见表3。由表3可知,与对照组相比,复合益生菌发酵饲料中黄曲霉毒素B1和呕吐毒素含量分别降低44.41%(P<0.01)和79.04%(P<0.01),玉米赤霉烯酮降低了16.89%(P<0.05)。
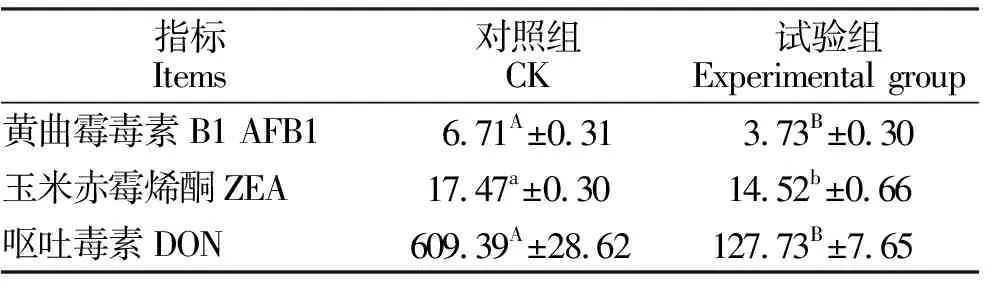
表3 复合益生菌发酵饲料对霉菌毒素含量的影响
2.4 复合益生菌发酵饲料的抑菌性能分析
2.4.1 复合益生菌发酵饲料对pH和挥发性脂肪酸的影响 复合益生菌发酵饲料对pH和挥发性脂肪酸的影响见表4。由表4可知,复合益生菌发酵饲料后pH显著下降(P<0.01),乳酸、乙酸及总挥发性脂肪酸含量极显著增加(P<0.01),丁酸和戊酸显著降低(P<0.01),丙酸无显著性差异(P>0.05)。

表4 复合益生菌发酵饲料对pH和挥发性脂肪酸的影响
2.4.2 复合益生菌发酵饲料后活菌数量和抑菌性能分析 复合益生菌发酵饲料后活菌数量和抑菌性能分析见表5。由表5可知,饲料经复合益生菌发酵后极显著增加乳酸菌与芽孢杆菌的菌落数(P<0.01)。采用正丁醇对发酵料的抑菌物质提取后结果显著,发酵饲料对大肠杆菌、沙门氏菌、痢疾杆菌的抑菌圈直径大小分别比未发酵饲料增加了60.00%(P<0.01)、31.58%(P<0.01)、42.86%(P<0.01)。

表5 复合益生菌发酵饲料后活菌数量和抑菌性能分析
3 讨 论
3.1 复合益生菌发酵饲料对营养成分的影响
酸溶蛋白是评价发酵饲料品质的主要指标之一,主要由肽和游离氨基酸等低分子量的蛋白水解物组成,具有溶于酸性溶液(三氯乙酸)的特性,可以一定程度上反映抗原蛋白和其他蛋白质抗营养因子被水解的程度,同时也反映小肽含量的高低[9]。王梅等[10]研究表明,益生菌发酵豆粕组中粗蛋白含量比对照组提高了6.5%,酸溶蛋白含量是对照组的3倍。本研究发现,益生菌发酵后的饲料粗蛋白显著增加,可能是由于益生菌发酵过程中微生物的大量增殖,提高了微生物菌体蛋白含量;酸溶蛋白含量的显著增加可能是发酵过程中产生的有机酸激活与蛋白质有关的消化酶,促进大分子蛋白降解成小肽利于动物消化吸收[3]。本试验中试验组DM含量低于对照组,与王洋等[11]研究相似,可能是因为饲料在发酵过程中微生物的增殖会消耗发酵底物成分,造成DM的损失。研究表明,乳酸菌发酵饲料对NDF和ADF无影响[12]。本试验结果表明,发酵后NDF、ADF与对照组均无显著性影响,可能是与饲料中含有DM有关,试验组NDF、ADF含量下降可能是发酵过程中芽孢杆菌产生的纤维素酶可以破坏纤维间的氢键,释放细胞内容物形成单糖[13]。进过益生菌发酵饲料可以提升CP、酸溶蛋白含量,降低不易吸收NDF、ADF含量,提升了饲料品质。
3.2 复合益生菌发酵饲料对抗原蛋白影响
肠道作为机体最大的营养器官,渗透性吸收是肠道固有的形态,指肠粘膜分子会把一些小分子物质以被动扩散吸收,大分子物质不被吸收的能力[14]。大豆球蛋白和β-大豆球蛋白是豆粕中主要的抗营养因子。日粮中大豆抗原蛋白含量高会破坏幼龄反刍动物肠道上皮完整性,抑制上皮细胞增殖,提高肠道上皮细胞炎性因子的分泌,引起过敏反应,导致腹泻[15]。陈中平等[16]研究表明,米曲霉发酵过程中产生的活性蛋白酶可将豆粕大分子蛋白亚基降解为小分子蛋白,有效降解豆粕的抗营养因子和致敏化合物。本试验结果显示,益生菌发酵后,酸性蛋白酶活性显著提升,使大豆球蛋白和β-伴大豆球蛋白降解,这与酸溶蛋白含量上升结果一致,说明经过益生菌发酵后的饲料更有利于动物肠道的消化吸收。
3.3 复合益生菌发酵对饲料中霉菌毒素的影响
霉菌毒素是一种有毒的次级代谢产物。黄曲霉毒素是世界公认的毒性最强的物质,主要与肝损伤有关,主要抑制DNA合成。呕吐毒素主体是脱氧雪腐镰刀菌烯醇(DON),其作用机制主要是抑制蛋白质合成以及细胞膜的通透性,从而破坏机体免疫机能。另外,玉米赤霉烯酮是一种类雌激素,它可以与雌激素竞争结合位点结合,对动物生殖生理造成损伤。原料以及配合饲料中极易受到霉菌毒素的污染,因此,饲料中有效的解毒措施是必不可少的。目前认为一些微生物主要通过降解与吸附作用进行有效的解毒。Petchkongkaew等[17]和Cho等[18]研究表明,芽孢杆菌通过代谢后产生的酶水解作用可降解黄曲霉毒素和玉米赤霉烯酮。Chen等[19]研究表明,乳酸菌和芽孢杆菌作为革兰氏阳性菌,有相同的细胞壁特征,通过细胞膜疏水性和细胞壁碳水化合物组分吸附真菌毒素使毒素含量降低。本研究表明,经过复合益生菌发酵后三种毒素菌显著降低,说明枯草芽孢杆菌B-1、地衣芽孢杆菌Y5-39和乳酸杆菌复合后发酵饲料可有效降低霉菌毒素含量,与以上研究结果一致。
3.4 复合益生菌发酵对饲料抑菌性能的影响
pH和有机酸作为评价发酵品质重要指标,良好的发酵饲料pH会从6.0左右降到4.0以下[20]。本试验结果表明经过益生菌发酵后饲料pH从6.0降低到3.4,与候楠楠等[21]研究结果一致。主要是因为固态发酵初期利用芽孢杆菌的好氧发酵消耗密闭环境的氧气为乳酸菌的厌氧发酵提供条件,乳酸菌可以将饲料中的葡萄糖等碳水化合物转化为乳酸等有机酸,从而降低饲料的pH。另外,乳酸具有调节肠道生态平衡、抑制病原菌、缓解腹泻、促进钙质吸收以及减缓应激的功能[10],说明益生菌发酵饲料可能对促进羔羊健康生长有重要作用。乙酸是良好的酸味剂以及降低环境恶臭的有益物质,丙酸是很好的防腐剂和诱食剂[22]。本研究中,发酵后期主要是乳酸菌起作用,饲料碳水化合物经过同质性乳酸与异质性乳酸发酵产生乳酸和乙酸,因此乙酸和乳酸含量较高,这与王梅等[10]研究结果相似。丁酸是腐败菌等微生物降解蛋白质、葡萄糖和乳酸的产物,丁酸含量增加会伴随着pH的升高及蛋白质的快速降解[23],本研究中益生菌发酵饲料丁酸含量比对照组降低了64.86%,提高了发酵饲料的品质。
动物肠道中主要由致病菌、有益微生物和病原体构成动态平衡[24]。肠道菌群微生物与宿主是共生关系,如果肠道菌群紊乱会造成动物腹泻。肠道主要致病菌有大肠杆菌和沙门氏菌,病原菌最适生长pH为6.0~7.0,发酵过程中会产生抗菌代谢产物、乳酸等有机酸,饲料乳酸菌菌落数提高又可进一步证明乳酸的产生,降低饲料pH,处于酸性环境,抑制有害菌的增长[25]。本研究结果表明,益生菌发酵后芽孢杆菌菌落数明显提高,经过正丁醇提取发酵后的代谢产物抑菌直径显著提高,酸性环境可加速乳酸菌代谢,产生的过氧化氢、乙醛、有机酸等物质,及芽孢杆菌代谢中产生杆菌素等物质,均对革兰氏阴性菌有抑制作用,同时产生的大量有益菌与有害菌形成竞争关系,主要是对饲料营养物质和空间位点的竞争,芽孢杆菌占有绝对优势后,有害菌不满足增殖条件从而达到抑菌作用。因此,益生菌发酵饲料的抑菌功能提高。
4 结 论
本试验条件下,玉米-豆粕型饲料经过复合益生菌发酵后品质显著提高,显著降低饲料中的抗营养因子与霉菌毒素且抑菌效果较好,有望用于动物饲养的一种新型功能性饲料。
猜你喜欢
湖南饲料(2022年6期)2022-12-20
湖南饲料(2021年4期)2021-10-13
当代水产(2021年2期)2021-03-29
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
猪业科学(2018年5期)2018-07-17
幸福(2017年18期)2018-01-03
饲料与畜牧(规模养猪)(2017年9期)2017-10-16
中国乳品工业(2017年1期)2017-02-20
