
CO2升高和施氮互作对木荷幼苗生理特性的影响
2022-11-04 02:07冯永霞尚鹤曹吉鑫倪秀雅陈展
生态环境学报 2022年9期
冯永霞,尚鹤,曹吉鑫,倪秀雅,陈展*
1.中国林业科学研究院森林生态环境与自然保护研究所/国家林业和草原局森林生态环境重点实验室,北京 100091;2.北京市园林绿化科学研究院,北京 100044
随着全球科技飞速发展,工业化进程加快,在人口密度越来越大的背景下,CO2、CH4等温室气体大量排放是导致气候异常、海平面上升等环境问题的重要原因之一(Kunalan et al.,2022)。工业革命前,地球的 CO2摩尔分数约为 280 μmol·mol-1,2021 年 CO2年均摩尔分数为 416.45 μmol·mol-1(https://www.co2.earth/)。IPCC第五次评估报告指出,在不同减排措施下大气 CO2摩尔分数到 2100年会出现 4种情景:430—480、580—720、720—1000、1000 μmol·mol-1以上(IPCC,2014)。因此研究不同路径CO2浓度对植物的影响,尤其是木本植物对高浓度CO2的响应是很有必要的,可以为森林植物应对未来气候变化提供一定的实验依据。
N素是调节植物器官代谢和生长的重要组成成分,也可以调节激素形成与平衡(Sun et al.,2020)。N沉降是全球性的环境问题,中国成为了第三大N沉降区(乔枫等,2018)。有研究表明,在全球范围内大气氮沉降约为 19 Tg·a-1(Schwede et al.,2018)。大气N沉降预计将在未来几十年快速增长(Wang et al.,2017)。
近十几年以来,CO2浓度升高对植物的影响已被广泛报道。大量研究探讨了CO2浓度升高对植物光合作用、叶绿素荧光特征的影响(Rodrigues et al.,2016),而对植物抗氧化系统以及内源激素的研究较少。抗氧化系统是植物防御外界环境胁迫和维持生长发育的基础,CO2浓度升高将影响植物光合作用过程,进而影响到植物的活性氧和抗氧化系统(Yi et al.,2020)。已有研究表明,短时间内CO2浓度升高可以增加光合色素含量(洪凯等,2020),且光合色素中的类胡萝卜素是一种非酶促抗氧化剂(张雪等,2016)。SOD是植物必不可少的一类抗氧化金属酶,它可以快速清除超氧自由基(O2-·),与其反应生成另一种活性氧过氧化氢(H2O2)(王星等,2014)。MDA是常见的膜脂过氧化指标,用来衡量植物细胞的膜脂过氧化程度,其含量越高,对植物细胞膜的毒害作用越强,膜脂过氧化程度就越大(Chandra et al.,2007)。CO2浓度升高能促进植物可溶性糖的积累,减轻膜质过氧化程度,保持相对较高的抗氧化酶活性和细胞结构的稳定性及膜透性,这有利于植物的生长发育(庄明浩等,2013)。内源激素是调控植物体生理机能的重要因素。当植物遇到非生物因素的胁迫时,ABA、IAA、ZR和GA3等植物内源激素在植物的生长和代谢的调节方面发挥着重要的作用(Sofo et al.,2013)。CO2作为绿色植物光合作用的底物,其浓度的变化直接影响到植物的生理生化过程和生长发育。大量研究显示,短时间内增加CO2浓度会提高农作物的产量和品质,具有“施肥效应”(姜帅等,2013)。也有研究表明,长期处于高CO2浓度下的小麦,叶片气孔导度与蒸腾速率的降低幅度以及光合速率的增大幅度都逐渐缩小,即发生了CO2驯化现象(范金杰等,2020)。因此不同植物对CO2浓度升高的响应大不相同。另外N素也是组成植物有机体的基本成分,是植物生长发育过程中必不可少的因素。有研究表明,施氮肥可以促进植物的生长,增强植物代谢和生物量(王满莲等,2017)。适当的氮添加会促进植物生长。氮添加也会对植物叶片色素含量有所增加,缺氮会对黄瓜幼苗产生胁迫作用并诱导植株抗氧化酶系统响应(张雪等,2016)。目前关于CO2浓度升高对植物的影响,大多探讨了 CO2浓度倍增(约 700—800 μmol·mol-1)对植物的影响(何梅等,2020;康红梅等,2020),但无法预测植物对高 CO2摩尔分数(1000 μmol·mol-1)的潜在响应。尽管一些报道研究了较高的 CO2摩尔分数(1000 μmol·mol-1)对作物的影响(张振花等,2018a),但有关IPCC报告预测的4种CO2浓度情景与N施肥对木本植物影响的试验却很少,而关于高 CO2摩尔分数(1000 μmol·mol-1)与N肥对植物生理生态的复合影响的报道更鲜见。因此探究IPCC报告中预测的所有CO2浓度情景及其与N肥对植物的复合影响,对进一步了解植物对未来气候变化响应情况具有重要的生态学意义。
木荷(Schima superba)主要分布于浙江、福建、江西、贵州、湖南、广西、广东、海南等地,不仅可用作木材加工,也被广泛应用于人工林种植,其生态和经济价值较高。本研究利用 OTC设备,根据IPCC第五次评估报告中的预测情景,设置4个CO2熏气浓度:自然大气 CO2摩尔分数(约 400 μmol·mol-1,CK)、550、750、1000 μmol·mol-1,在不同浓度水平下设置两个不同施氮梯度(低氮:0 g·m-2·a-1,高氮:10 g·m-2·a-1),研究对象为一年生的木荷幼年植株,研究CO2浓度升高与施氮对木荷的光合色素以及抗氧化活性等的影响,旨在为气候变化背景下具有高生态及经济价值的树木的适应性栽培提供依据。
1 材料与方法
1.1 研究区概况
该试验在中国科学院千烟洲红壤丘陵生态试验站(26°44'48"N,115°04'13"E)开展,该站位于中国中南部的江西省泰和县灌溪镇,海拔高度约100 m,年均温为17.9 ℃,年均降雨量为1489 mm,属于亚热带季风气候。植被属于中亚热带常绿阔叶林带。主要的土壤类型有红壤、水稻土、潮土、草甸土等。
1.2 试验处理
本试验设置4个CO2处理:设置OTC设备但不熏气作为对照(CK,使其与自然大气 CO2浓度保持一致)、OTC设备分别通入 550、750、1000 μmol·mol-1CO2。每个处理重复3次,共设置12个OTC设备。根据国家监测网的数据,总氮沉降平均约为 4 g·m-2·a-1,且最大值超过 8 g·m-2·a-1(Xu et al.,2015),中国作为世界上氮沉降最高的几个地区之一,过去30年增长了约60%(Liu et al.,2013)。考虑到N沉降的增长趋势,本研究在每个OTC设备内设置的施氮水平为10 g·m-2·a-1(高氮),同时设置不施氮肥(低氮)的处理,共两个N浓度梯度。OTC设备运行系统详见文献(潘鸿等,2022)。
每个 OTC中放入来自于吉安市泰和县当地苗圃的一年生盆栽木荷幼苗 10株,其平均株高为40.32 cm,平均地径约为5.64 mm。在2018年3月初选取生长状况较一致的木荷幼苗移栽到花盆中(直径25 cm,高度18.5 cm),栽培土壤为试验站当地的红壤。试验开始前对红壤进行检测,其土壤有机质质量分数为 (8.63±0.32) g·kg-1,全氮为500 mg·kg-1,速效磷为 (1.58±0.17) mg·kg-1,速效钾为(20.10±0.36) mg·kg-1,pH 值为 4.50±0.02。移栽后的木荷幼苗在自然环境中缓苗 20天后搬入 OTC内。试验时间为2018年6月30日—2018年10月1日,每日对其进行 9 h熏气(北京时间 08:00—17:00)。施氮处理需先计算木荷盆栽的土壤表面积,再称取不同质量的KNO3(>99%)溶于水中配制成不同浓度的KNO3溶液,每个OTC中选择5株木荷幼苗进行施氮处理。2018年6月15日、7月20日、8月24日和9月20日对其施肥,根据盆栽的土壤表面积,称取一定量KNO3配制成6.8 g·L-1的KNO3溶液,每次每盆喷施150 mL该溶液;同时不施氮处理的盆栽喷淋等量自来水作为低氮对照。试验过程中,根据植物的具体需要使用同一量杯施加水分,确保每株木荷幼苗的水分供应充足且等量。
1.3 测定方法
2018年10初试验结束后,从每个处理中随机选择2株植株,每株在主干从上到下3—5叶位选择2片生长状况正常、叶片展开完全的叶,避开叶脉将其叶片剪碎后用锡箔纸包裹,用液氮罐保存运输到实验室进行分析。光合色素的测定参考叶思源(2019),超氧化物歧化酶(SOD)采用氮蓝四唑(NBT)光化还原法测定(李仪曼等,2019)。丙二醛(MDA)含量采用硫代巴比妥酸法测定(于浩,2017)。利用酶联免疫吸附剂测定法测定 IAA、ABA、GA3和ZR的含量。同时从每个处理中选择随机选择2盆木荷植株用于生物量的测定,首先使用自来水将植物清洗,并用植物剪分成3部分(根、茎、叶),在烘箱中烘干到恒重状态后称重,烘箱温度设置为65 ℃。
1.4 数据分析
应用 Microsoft Excel对数据进行初步分析处理;在SPSS 21.0统计软件中,先进行Kolmogorov-Smirnov和 Levene检验,所有数据均满足正态性以及方差齐性。采用多因素方差分析,探讨 CO2浓度、施氮及其交互作用对木荷生理生化的影响。单因素方差分析分别检验低氮和高氮处理下不同CO2浓度的影响,采用独立样本t检验分析同一CO2浓度处理下高氮和低氮处理之间的差异,并采用最小显著差数法(LSD)进行多重检验(P<0.05)。应用Origin 2017进行绘图。
2 结果分析
2.1 CO2浓度升高和施氮互作对木荷幼苗光合色素的影响
多因素方差分析结果表明,CO2浓度升高对植物叶片叶绿素a和类胡萝卜素的含量都没有显著影响,CO2显著影响叶绿素b的含量(P=0.020),而氮肥则显著影响光合色素(P<0.001),CO2和氮肥的交互作用对光合色素无显著影响。单因素方差分析结果表明,除 1000 μmol·mol-1CO2处理,其他CO2处理下,高氮处理下的光合色素含量均显著高于低氮处理(图1)。在CK处理中,高氮与低氮相比叶绿素a含量显著增加了165.63%(P=0.007),叶绿素b含量增加了181.43%(P=0.007)。在550 μmol·mol-1CO2处理下,高氮处理与低氮处理相比叶绿素 a、叶绿素 b含量分别增加了 123.79%(P=0.017)、101.14%(P=0.044)。在 750 μmol·mol-1CO2处理下,高氮处理与低氮处理相比叶绿素a含量显著增加了157.93%(P=0.01),叶绿素b含量分别显著增加了116.66%(P=0.007)。在CK、550、750 μmol·mol-13个CO2处理中,类胡萝卜素含量在高氮水平下平均增加80%左右(P分别为0.013,0.012,0.006)。在 1000 μmol·mol-1的 CO2处理下,高氮处理虽比低氮处理下的色素含量也有所增加,但是增幅不显著。相同N肥处理下,随着CO2浓度的增加,叶绿素 a、类胡萝卜素的含量并没有显著差异。但与不熏气处理相比,1000 μmol·mol-1CO2处理显著降低了叶绿素b的含量。

图1 不同CO2和氮肥水平对木荷幼苗光合色素的影响Figure 1 Different effects of elevated CO2 and N fertilization on photosynthetic pigments in Schima superba seedlings
2.2 CO2浓度升高和施氮互作对木荷幼苗抗氧化系统的影响
多因素方差分析结果表示,增加 CO2浓度、氮添加及其交互作用均对MDA含量无显著影响;CO2浓度和氮肥均对 SOD活力有显著影响,但CO2和氮肥的交互作用对 SOD活力无显著影响(图2)。而单因素方差分析发现在550 μmol·mol-1CO2处理下,高氮处理MDA含量较低氮处理显著增加了 33.77%(P=0.046)。低氮处理下,随着CO2摩尔分数增加到 550、750、1000 μmol·mol-1时,SOD活力分别较 CK提高了 24.59%(P=0.015)、19.44%(P=0.038)、24.98%(P=0.022)。在 550、750 μmol·mol-1CO2处理下,高氮较低氮的SOD的活力下降了31.86%(P=0.011)、19.33%(P=0.018)。
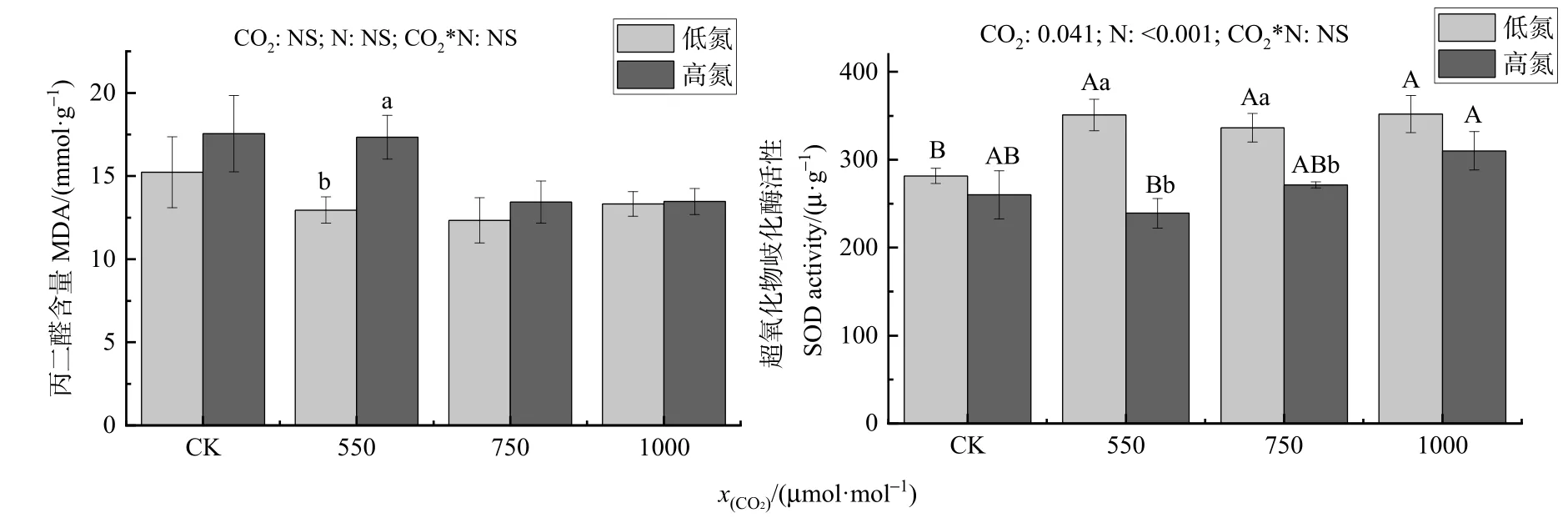
图2 不同CO2和氮肥水平对木荷幼苗丙二醛和超氧化物歧化酶活力的影响Figure 2 Effects of different CO2 and N fertilization on the activities of malondialdehyde and superoxide dismutase in Schima superba seedlings
2.3 CO2浓度升高和施氮互作对木荷幼苗内源激素的影响
多因素方差分析结果显示,增加CO2浓度、施氮以及CO2浓度和氮肥的交互作用都对脱落酸没有显著影响;CO2浓度及其与氮肥的交互作用对生长素有显著影响(P<0.05),而氮肥对IAA无显著影响。CO2浓度及其与氮肥的交互作用对ZR和GA3无显著影响,而施氮对木荷幼苗叶片ZR和GA3含量具有显著影响(P<0.05)(图3)。
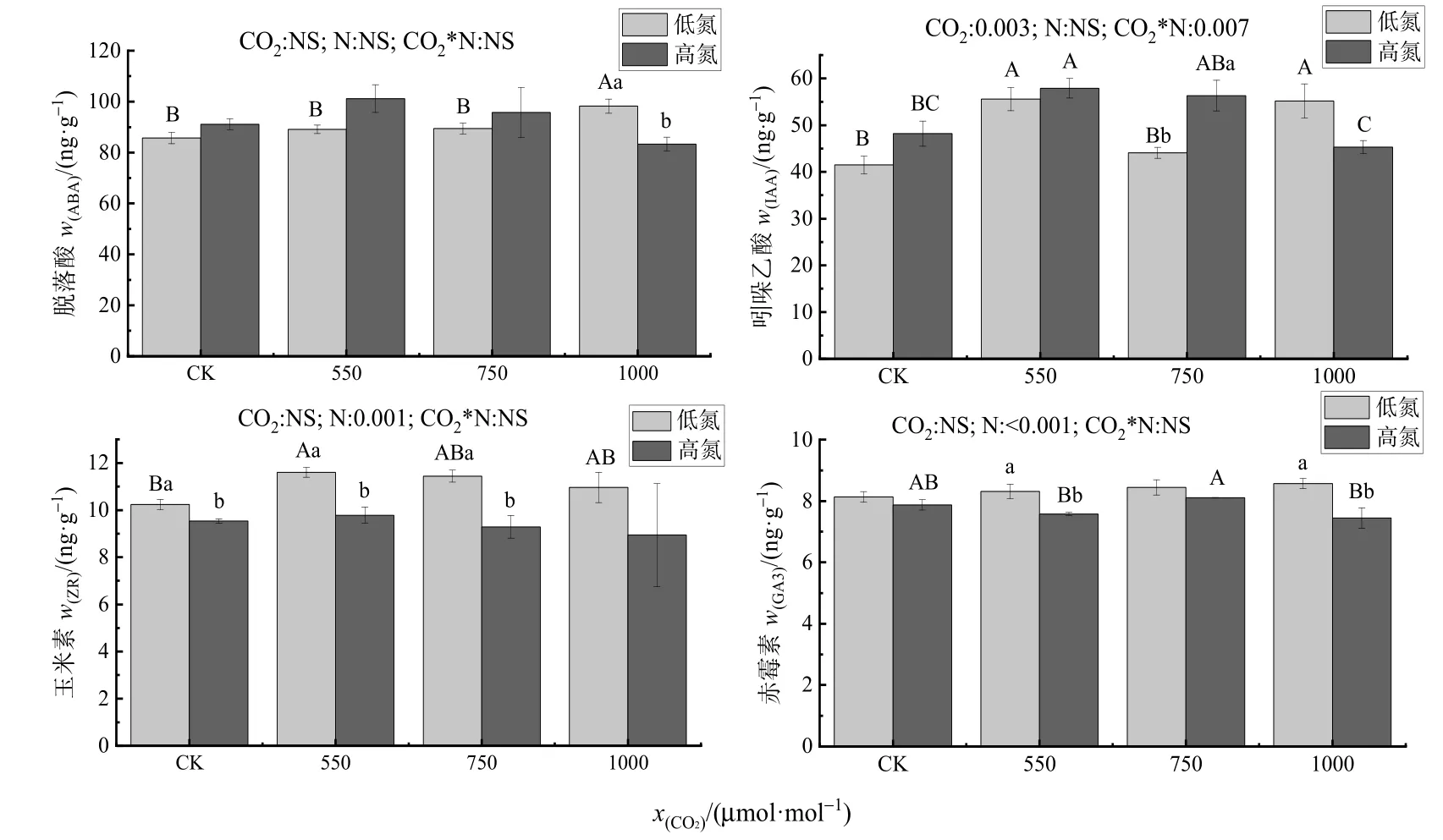
图3 不同CO2和氮肥水平对木荷幼苗内源激素含量的影响Figure 3 Effects of different CO2 and N fertilization on the content of endogenous hormones in Schima superba seedlings
单因素方差分析则表明,在低氮情况下,1000 μmol·mol-1CO2处理较其他 3 种 CO2处理,ABA分别显著增加了14.65%(P=0.004)、10.26%(P=0.021)和9.81%(P=0.025)。低氮处理下,随着 CO2摩尔分数升高至 550、1000 μmol·mol-1,IAA的含量较CK分别增加了34.03%(P=0.016)和 33.04%(P=0.005)。高氮处理下,随着 CO2摩尔分数升高至 550 μmol·mol-1,IAA 的含量较CK也增加了20.16%(P=0.031)。在750 μmol·mol-1CO2处理下,施氮较低氮处理的 IAA含量增加21.73%(P=0.025)。除 1000 μmol·mol-1CO2处理,在其他CO2处理下,与低氮处理相比,高氮显著降低了 ZR 的含量。400 μmol·mol-1CO2处理下,高氮较低氮的 ZR含量降低了 6.82%(P=0.045)。550 μmol·mol-1CO2处理下,高氮较低氮的ZR含量降低了15.70%(P=0.011)。750 μmol·mol-1CO2处理下,高氮较低氮的ZR含量降低了 18.83%(P=0.017)。在 550 μmol·mol-1和1000 μmol·mol-1CO2处理下,高氮较低氮的 GA3含量分别下降了 8.84%(P=0.039)和 13.17%(P=0.040)。
2.4 CO2浓度升高和施氮互作对木荷幼苗植物生长量的影响
多因素方差分析结果表明,CO2对植物总生物量没有显著影响;而施氮对植物的总生物量有显著影响(P<0.001),CO2和施氮的交互作用对植物生物量无显著影响(表1)。单因素方差分析则表明,在低氮处理下,随着CO2浓度升高,木荷幼苗生物量没有明显变化。而在高氮处理下,1000 μmol·mol-1CO2处理下木荷幼苗的总生物量较 CK提高了 52.79%(P=0.044)。在 1000 μmol·mol-1CO2处理下,与低氮处理相比,高氮显著增加了木荷总生物量,高氮比低氮的木荷总生物量增加了106.38%(P=0.003)。
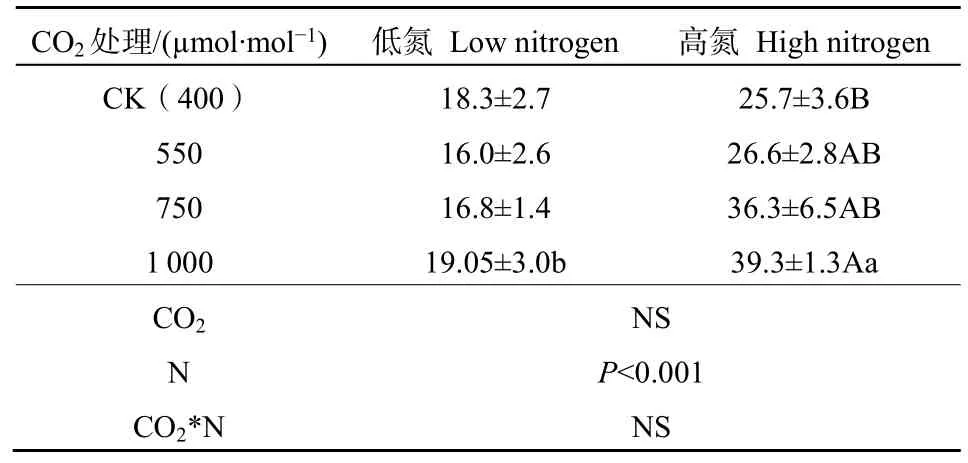
表1 不同CO2和氮肥水平对木荷幼苗总生物量的影响Table 1 Effects of elevated CO2 and N fertilization on plant biomass g·plant-1
3 讨论
3.1 对光合色素的影响
本试验结果表明,随着CO2浓度的升高,其幼苗叶绿素a和类胡萝卜素含量没有显著变化,而在1000 μmol·mol-1的CO2下显著降低了叶绿素b的含量(图1)。而以往的一些研究发现CO2浓度升高会促进植物叶绿素含量的上升(张振花等,2018b)。也有研究表明CO2浓度上升会抑制叶绿素含量,如CO2浓度倍增条件下栓皮栎(Quercus variabilis)的光合色素含量下降(杜启燃等,2014)。而本试验中高浓度CO2下会大大降低木荷的叶绿素b含量,这与王精明(2005)的结果相似。本课题组前期研究也证实叶N含量随着CO2浓度的上升而逐渐下降(Chen et al.,2021)。而叶绿素本身是一类含氮化合物,高浓度CO2将促进植物生长,同时使得与叶片光合色素合成有关的叶N含量下降,进而影响光合色素的合成,使得光合色素含量下降,即稀释效应(陈雨娇等,2020)。同时试验结果表明,木荷幼苗叶片的光合色素含量会随着氮肥水平的增加而显著增加,这与甜菜(Beta vulgarisL.)(张翼飞等,2013)、毛竹(Phyllostachys edulis)(Gao et al.,2016)以及黑麦草(Lolium multiflorumLam.)(Lü et al.,2021)等的研究结果一致。这可能是植物体内光合色素的生成均需要卜琳环与Mg2+结合,而氮元素是这一过程的关键元素(高培军,2013);同时氮是构成植物体内大分子物质如核酸、蛋白质和光合酶等的重要组成成分(Zhang et al.,2022)。因此,植物体内氮素的积累可以使得参与光合色素生成的相关酶浓度和活性有所增加,从而间接导致了植物体内光合色素的积累。但当CO2熏气摩尔分数达到 1000 μmol·mol-1时,氮素也不再对光合色素有所影响。这可能是因为高CO2浓度下,导致植物的气孔导度逐渐关闭,植物对CO2的吸收大量减少,而使得光合作用下降,高CO2浓度下植物的生长发育也受到限制(圣倩倩等,2021),进而植物也不受氮的影响。
3.2 对抗氧化活性的影响
有研究显示,高浓度 CO2下银杏(Ginkgo biloba)叶片中的丙二醛含量将显著下降,这表明高浓度CO2可以减弱叶片的膜脂过氧化能力并对氧化损伤具有防护作用(阮亚男等,2007)。本试验中,在低氮处理下,随着 CO2浓度的上升,MDA含量虽无显著变化但有下降趋势(图 2),这可能是因为胞间CO2浓度上升,CO2同化率增加,使得有关呼吸作用的酶活性降低,呼吸速率降低,产生的活性氧也降低,H2O2产生也受到抑制,因此植物体内MDA含量减少,木荷幼苗在高CO2浓度处理在一定程度上降低影响膜脂过氧化作用(翟晓朦,2015)。植物本身具备一套抗氧化系统,正常生长发育时抗氧化系统处于动态平衡中,而在各种逆境胁迫时,有关抗氧化活性的酶会增加(Denaxa et al.,2020)。在非生物胁迫下,植物会调控抗氧化酶基因的表达来减少活性氧(ROS)造成的伤害(张星雨等,2021)。本试验中,低氮处理下,500、750、1000 μmol·mol-1CO2处理的SOD活力较CK显著增加。而在高氮下,1000 μmol·mol-1CO2处理下SOD活力较 550 μmol·mol-1显著增加(图 2)。这与吴志丹等(2014)的研究结果一致。高浓度CO2条件下木荷幼苗的 SOD活力增强。这可能是由于高浓度CO2能够提高叶绿体PSII内NADP+的利用率,产生大量的 NADPH,促进了卡尔文循环和抗坏血酸-谷胱甘肽循环(ASA-GSH cycle),从而增加抗氧化酶活性(阮亚男等,2007),故高浓度CO2会提高植物自身的抗性和逆境抵抗能力,促进木荷幼苗正常生长。在 550、750 μmol·mol-1CO2处理下,高氮较低氮的 SOD活力显著下降。这可能是因为在氮限制的区域里,施氮会促进了植物生长,而非胁迫,故木荷幼苗的抗氧化酶活性降低(熊露露等,2021)。
3.3 对植物内源激素的影响
本试验中,在低氮处理下,1000 μmol·mol-1CO2处理ABA的含量较其他CO2处理显著增加(图3)。CO2浓度增加会降低叶片气孔孔径和气孔密度,从而导致蒸腾减少。CO2浓度升高通过进入保卫细胞 ABA信号通路而使得气孔导度减小,并使得保卫细胞中ABA生物合成增加(Chater et al.,2015)。有研究表明,施氮处理下植物中 ABA含量明显降低(Ye et al.,2021)。而在 400、550、750 μmol·mol-1CO2处理下,施氮处理下 ABA含量明显有增加的趋势(图 3)。这与李文娆等(2017)的研究施氮下对棉花(Gossypium hirsutumL.)幼苗根源的研究结果一致。这可能是因为 NO3-被大量吸收使得细胞中有机酸合成增加,导致细胞质酸化,进而使保卫细胞中积累大量 ABA(李庆余,2010)。但在1000 μmol·mol-1CO2处理下,高氮较低氮显著降低了ABA的含量。这可能是高浓度的CO2下导致CO2在叶肉细胞中积累,而1-磷酸葡萄糖在相关酶的催化下合成淀粉,从而使细胞液渗透势升高,保卫细胞失水,气孔关闭,导致ABA含量降低。IAA可以改变植物体内的营养物质分配,也可以影响植物生长和细胞伸长、分化等过程。在本试验的高氮处理下,随着 CO2摩尔分数升高至 550 μmol·mol-1时,IAA的含量较CK处理增加了20.16%(P=0.031)(图3),这与Wang et al.(2009)的结果一致。但在 750 μmol·mol-1CO2处理下,IAA 含量没有显著增加,这与Pei et al.(2020)的结果一致。这可能是因为一定程度的CO2浓度升高使得可溶性糖在叶肉细胞中累积从而导致植物的呼吸作用增强,植物呼吸作用产生能量会刺激分生区IAA的合成,所以CO2浓度升高时会促进叶片中生长素的合成(翟开恩等,2015)。在 750μmol·mol-1CO2处理下,高氮较低氮的IAA含量增加21.73%(P=0.025)。徐国伟等(2018)研究表明,施氮有利于根系细胞生长素的合成。这可能是因为施氮能够促进根系分生组织和地上部分的生长,促进IAA的合成(赵平等,2005)。本试验中,除 1000 μmol·mol-1处理外,其他CO2处理下,施氮降低了内源ZR的含量(图3)。这与袁昌洪等(2016)在施氮处理后“乌牛早”嫩芽(Camellia sinensis‘Wuniuzao’)中 IAA、GA3、ZR含量增加的结果不同,但与李洪娜等(2015)的研究表明施氮使得ZR和GA3含量下降的结果一致。GA3可以调节植物的生长和发育,对环境压力变化敏感。翟晓朦等(2016)发现苜蓿(Medicago sativa)在大气 CO2浓度升高后,GA3含量总体上趋于升高的趋势。陈晓娜等(2020)的研究表明氮素对 GA3的促进效果较显著。但在本试验中,CO2浓度对GA3的促进效果并不显著。在550 μmol·mol-1和 1000 μmol·mol-1CO2处理下,高氮处理下较低氮处理下的GA3有所下降(图3),这可能是大量的ABA积累抑制了ZR、GA3的合成和运输(郭文琦等,2010)。同时ZR可以提高SOD等抗氧化酶的活力从而有利于活性氧的清除(于浩,2017),而本研究中施氮降低了ZR的含量,所以也间接降低了SOD活性。
3.4 对植物生物量的影响
CO2浓度的升高能促进植物的枝、茎和节间的生长,增加生物量(曾贞等,2016)。本研究中,高氮条件下,当 CO2摩尔分数升高至 550、750 μmol·mol-1时对木荷幼苗生物量没有显著差异,而当 CO2摩尔分数升高至 1000 μmol·mol-1,木荷幼苗的总生物量则较CK显著增加了52.78%(表1)。这与潘鸿等(2022)发现CO2浓度升高使木荷幼苗生物量增加的结果一致。本研究区土壤全N含量为500 mg·kg-1,相对较低。由表1结果可知,在未施氮肥的情况下,CO2浓度升高对木荷幼苗的生物量没有显著变化,这是因为当可利用的氮含量较低时会限制 CO2对生物量的促进作用(Chen et al.,2021)。而当不考虑任何CO2浓度条件时,施氮显著增加了木荷幼苗的生物量。常宏等(2019)研究表明施氮显著增加了壳斗科(Fagaceae)3种幼树的总生物量。Forsmark et al.(2021)的研究表明人为氮添加会明显增加樟子松(Pinus sylvestrisL.)针叶和细根的生物量。这是因为施氮会提高土壤可利用性氮,植物吸收利用氮肥并促进了植物光合色素的增加以及植物生殖生长,使得植物的生长量也有所增加。
综上所述,虽然CO2升高对木荷幼苗的叶绿素a、类胡萝卜素无显著影响,但对叶绿素 b有显著影响。施氮显著增加了木荷幼苗的光合色素含量,随之其光合能力也逐渐增强,同时促进植物的生长,其生物量也随之增加。随着CO2浓度的上升,MDA含量有无显著变化,而SOD活力上升。这说明木荷幼苗具有一定的程度的CO2胁迫抗性,可以通过增强抗氧化酶的活性来缓解高浓度CO2对植物产生的胁迫,降低膜脂过氧化水平,减轻细胞膜的损伤,以保证木荷幼苗的正常代谢活动。且施氮有效地增加IAA的含量,从而促进生长。
4 结论
(1)N添加显著促进光合色素含量(P<0.001),这有助于促进木荷幼苗光合作用速率,从而提高木荷幼苗生物量。
(2)CO2浓度升高和氮添加的交互作用对MDA含量和SOD活性无显著影响,而CO2浓度升高对SOD活性有提升作用(P=0.041),氮添加对SOD活力有抑制作用(P=0.001)。这表明木荷幼苗在CO2浓度升高时会有一定的自我调节能力,可以通过增强抗氧化酶的活性来缓解高浓度CO2对植物产生的影响,降低膜脂过氧化水平,减轻细胞膜的损伤,以保证木荷幼苗的正常代谢活动。
(3)CO2浓度对 IAA含量有促进作用(P=0.003),施氮对 ZR、GA3的含量有抑制作用(P<0.001)。激素在木荷幼苗应对高浓度CO2时发挥着重要的调节作用,在一定程度CO2浓度下,IAA含量增加会促进木荷幼苗的生长。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
林业科学(2022年5期)2022-08-20
散文诗世界(2022年5期)2022-05-23
中国土壤与肥料(2021年5期)2021-12-02
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
山东农业科学(2017年3期)2017-03-29
江苏农业科学(2016年4期)2016-06-14
农家顾问(2014年9期)2014-10-20
