
2种生境下翠芦莉营养器官解剖比较
2022-11-04 07:36:50朱栗琼宾思晨吕佳峻杨凤兰
江西农业学报 2022年8期
陆 晴,朱栗琼,宾思晨,吕佳峻,杨凤兰
(1.广西大学 林学院,广西 南宁 530004;2.广西农业职业技术大学,广西 南宁 530007)
根、茎和叶都属于植物的营养器官,能够维持植物的生命活动。其中植物主要依靠根和茎吸收、运输水分及营养物质,通过叶完成光合作用,为植物体生产有机物[1]。当植物在特定环境中生长时,其结构会产生一定的变化来适应环境。植物营养器官的解剖结构可以反映植物对环境适应的情况,在一定程度上显示出其对环境的适应策略和能力。影响植物的环境因子主要包括温度、光照、水分、土壤、大气及其他生物有机体。在不同水分的环境中,植物的组织结构特征会发生适应性改变,其结构特征通常可作为评价植物抗旱性能的重要标准[2]。
翠芦莉(Ruellia brittoniana),又名芦莉草、蓝花草,属于爵床科(Acanthaceae)多年生常绿草本植物,其具有高性或者矮性,花期从春到秋,果期为夏秋。对光照要求不严格,全日照或半日照均适宜生长,生长适温为22~30 ℃。翠芦莉具有比较强的抗逆性,其抗旱、耐涝且耐轻盐碱,常用于街道和公园的绿化[3]。
目前关于翠芦莉的研究多在水污染净化[4-5]以及耐涝性[6]、耐旱性[7-8]等方面。翠芦莉作为园林景观营造中观赏价值较高的夏季花卉,其营养器官的解剖结构研究还未见报道。翠芦莉在水、旱两种环境中都长势良好,推测其根茎或叶器官中具有与水生和旱生环境相适应的解剖学构造特征,其与水输送、贮存与散失等有关结构的解剖学特点仍有待进一步深入研究。本研究以水生和旱生2种生境中生长的翠芦莉为对象,进行了解剖学的比较,探讨了翠芦莉营养器官的结构特征与环境变化之间的关系,以期为翠芦莉推广栽培及生态适应性的研究提供理论依据。
1 材料与方法
1.1 材料
翠芦莉采集自南宁市广西大学校园内,南宁市属于湿润的亚热带季风气候,阳光充足,全年雨量充沛,年平均气温在21.6 ℃左右。水生样地土壤含水量大于20%,为沿水下沉式绿地;旱生样地土壤湿度小于15%,土壤板结且受到道路硬质铺装影响。于2019年10月初进行翠芦莉植物样本采集,选择鱼塘旁被水淹的栽植点为水生样地,干旱环境样地选择建筑垃圾填埋地、土层瘠薄的栽植点。在水生和旱生2种生境下各选择6株长势中等的植株,以距植株基部20~30 cm为半径挖掘整株植株。取离顶芽第3~4对的成熟叶片,保留中脉,剪取叶的中间部分,大小约4 mm×5 mm。选取每株植物颜色新鲜的须状根中下部各5条,取离根尖处5~6 cm;茎截取中部均匀段,再将根、茎切成1 cm左右小段;最后将植物材料保存到FAA固定液中。
1.2 制片方法
将完全脱色后的植物材料用不同浓度的乙醇脱水,经过浸蜡后用常规石蜡切片法切片,切片厚度为5 μm;经番红—固绿双重染色,用中性树胶迅速封片[9]。最后用光学显微镜观察并拍照,拍摄视野不少于30个。
叶表皮微观形态采用离析法制片。将样本放入FAA液中固定7 d,取出后使用蒸馏水冲洗干净表面的残留,将其浸泡在H2O2和冰醋酸(体积比为1:1)溶液中,60 ℃水浴加热至叶片变白、产生气泡后,撕取叶表皮,用小毛刷去除叶肉组织。使用1%番红溶液染色并装片,用Nikon E100光学显微镜进行观察并拍照。
1.3 数据分析方法
应用Image J软件测量数据,各指标均以30个视野中随机测量和观察的30个数值的平均值进行统计和计算。所有数据采用SPSS 22.0软件进行分析。
2 结果与分析
2.1 根解剖结构的比较
2.1.1 根解剖结构的形态观察 观察发现,翠芦莉幼根横切结构为初生结构(图1),由表皮、皮层、维管柱三部分组成。其中表皮细胞只有1层,已经死亡并木栓化,位于最外层形成根被。皮层是位于表皮之内维管束之外的多层薄壁细胞,旱生环境中翠芦莉的皮层数为20~22层,薄壁细胞排列较为整齐,形状较规则,多为圆形或长方形,细胞大小不一;水生环境中翠芦莉的皮层数比旱生环境多,为32~34层,薄壁细胞形状较规则,多为圆形,排列没有前者整齐。维管柱是内皮层以内的部分,主要负责运输水分和光合产物,包括初生木质部、初生韧皮部、结合组织、中柱鞘和髓。2种生境中翠芦莉的木质部中的木射线明显,导管口径大小不一,水生环境中翠芦莉的导管比旱生环境多且排列呈线状分布。维管柱中央具有由薄壁细胞组成的髓,水生环境中翠芦莉的髓比旱生环境稍大。
2.1.2 根解剖结构的量化参数比较 经方差分析,2种生境下翠芦莉的皮层厚度、皮层和髓所占根半径比例差异显著,表皮、维管束、髓厚度差异不显著(图2)。与水生环境相比,旱生环境中翠芦莉根的皮层厚度和皮层所占比例显著减小,髓厚度所占比例显著增加。水生环境和旱生环境下翠芦莉根的皮层厚度分别为(905.94±148.13) μm和(740.09±74.25) μm,皮层所占比例分别为69.17%±4%和63.02%±4.35%。水生环境植株在这2个指标上均显著高于旱生环境,且前者的皮层厚度和皮层所占比例分别是后者的1.22倍和1.10倍。2种生境中髓的厚度虽无明显差异,但旱生环境下髓所占比例(17.13%±2.67%)是水生环境下(13.29%±2.28%)的1.29倍,具有显著性差异。
2.2 茎解剖结构的比较
2.2.1 茎初生解剖结构的形态观察 翠芦莉初生茎解剖结构见图3,均由表皮、皮层、中柱三部分组成,其中表皮由1层细胞组成,形状椭圆形且排列整齐而紧密,表皮外附有角质层。皮层包含皮层薄壁组织和厚角组织,靠近表层的厚角组织细胞较小,大小近似表皮细胞。旱生环境下翠芦莉的厚角组织由6~7层细胞构成,皮层薄壁组织由8~9层细胞构成;水生环境下翠芦莉的厚角组织由5~6层细胞构成,皮层薄壁组织由9~10层细胞构成。两者中柱均由维管束、髓和髓射线组成,其中维管束由初生木质部、束间形成层和初生韧皮部组成。髓射线清晰可见,髓位于茎中心部位,由薄壁细胞组成,厚度较大。
2.2.2 茎初生解剖结构的量化参数比较 经方差分析,2种生境下翠芦莉的角质层厚度、表皮厚度、维管束厚度、初生韧皮部厚度、初生木质部厚度和初生木质部与维管束的比例存在显著性差异(图4)。与水生环境相比,旱生环境下翠芦莉的角质层、维管束、初生韧皮部、初生木质部、初生木质部与维管束的比例明显增加。旱生环境下翠芦莉的角质层厚度为(6.57±1.31) μm,为水生环境的1.32倍。旱生环境和水生环境下翠芦莉的维管束厚度分别为(139.22±26.02) μm和(114.38±22.59) μm,初 生 木 质 部 分 别 为(80.87±13.54) μm和(53.19±19.56) μm,初生木质部与维管束的比例分别为58.67%±7.08%和46.02%±10.91%,前者均显著高于后者。虽然旱生环境下的翠芦莉初生韧皮部厚度为(28.29±6.67) μm,比水生环境厚5.48 μm,但两者的初生韧皮部与维管束的比例并无显著差异。旱生环境下翠芦莉的表皮细胞厚度低于水生环境,分别为(12.30±1.87) μm和(14.22±3.31) μm。
2.3 叶解剖结构的比较
2.3.1 叶解剖结构的形态观察 翠芦莉叶结构由表皮、叶肉、叶脉组成(图5)。其中表皮包含上、下表皮,表皮均有双环形气孔器呈不规则分布,并覆有厚薄不等的角质层,上角质层比下角质层厚。表皮细胞为1层,形状为规则的方形,排列紧密。通过对比可知,水生环境下翠芦莉的表皮细胞明显大于旱生环境。叶肉组织发达,为异面叶,明显分化出栅栏组织和海绵组织。叶片的海绵组织由2~3层细胞组成,细胞大小不等,近圆形,含有叶绿体,细胞排列疏松,有间隙;旱生环境下翠芦莉的海绵组织排列比较杂乱,而水生环境下排列得较规则且部分海绵组织发育成发达的储水组织。2种生境下翠芦莉的栅栏组织均为2层,上层长柱状,下层短柱状。但旱生环境中的翠芦莉栅栏组织排列较为疏松;水生环境下的栅栏组织排列较为紧密,且含有较多叶绿体,叶绿体多分布在边缘。二者叶片主脉发达,由角质层、表皮、厚壁组织、薄壁细胞和维管束构成。主脉表皮外被角质层,厚壁组织2~3层,薄壁细胞6~8层。维管束形状较为相似,均为长椭圆形,导管呈径向放射排列,但旱生环境下翠芦莉的维管束比水生环境长。
2.3.2 叶表皮气孔量化参数比较 由表1可知,2种生境中的翠芦莉叶片气孔指标有差异,从气孔指数方面来看,旱生环境下翠芦莉的上、下表皮气孔指数均较大,分别是0.10±0.29和30.57±3.27,显著高于水生环境下的0.04±0.09和28.33±3.39。从气孔密度方面来看,旱生环境下翠芦莉的气孔密度较大,平均每个视野内有255.40±30.22个气孔,显著高于水生环境。从气孔的大小来看,两者气孔长度无显著差异,但旱生环境下翠芦莉的气孔宽度显著高于水生环境。
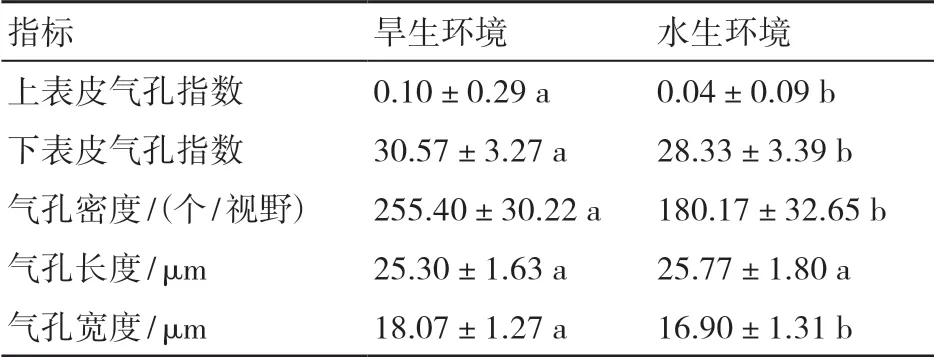
表1 2种生境中翠芦莉叶表皮气孔特征比较
2.3.3 叶片解剖结构量化参数比较 2种生境中的翠芦莉叶片在叶片、上角质层、上下表皮、栅栏组织、海绵组织的厚度、栅海比、叶片结构紧密度、叶片结构疏松度及主脉维管束长度上均存在显著性差异,而下角质层、主脉厚度和主脉维管束宽度无显著差异(表2)。水生环境下翠芦莉叶片厚度是旱生环境的1.08倍,上表皮细胞厚度是后者的1.76倍,下表皮细胞厚度是1.62倍,海绵组织厚度是1.24倍,叶片疏松度是1.14倍。旱生环境下翠芦莉的上角质层厚度是水生环境的1.14倍,栅栏组织厚度是后者的1.51倍,主脉维管束长度是1.49倍。旱生环境下翠芦莉的栅栏组织厚度、栅海比、叶片结构紧密度均大于水生环境,说明旱地环境中翠芦莉叶片的栅栏组织排列更紧密且更发达。

表2 2种生境中翠芦莉叶片解剖结构特征比较
2.3.4 叶片解剖特征可塑性比较 通过计算得到不同生境翠芦莉叶片解剖结构的变异系数和可塑性指数(表3),结果表明:叶片厚度、角质层厚度、上表皮细胞厚度、栅栏组织厚度的变异系数及可塑性指数均表现为旱生环境中的较大。从叶片大部分解剖结构和叶片整体平均水平来看,在旱生环境中,翠芦莉叶片结构变异较大,说明它适应了更加多样化的生态环境。同时,2种生境中叶片的主脉维管束长度的变异系数和可塑性指数都最大,说明主脉维管束长度更易对环境变化做出改变,可作为一个较好的指标用于评价植物的环境适应能力。旱生环境中翠芦莉的叶片结构紧密度变异系数最低,主脉厚度可塑性指数最低;水生环境中翠芦莉叶片厚度的变异系数和可塑性指数最低,可认为是较稳定的系统演化性状。

表3 不同生境翠芦莉叶片解剖结构的变异系数和可塑性指数
3 结论与讨论
植物为了适应干旱环境,会通过减小根皮层数目及皮层厚度的方式来减小水分运输的距离及阻力,从而提高水分的运输速率[10]。翠芦莉的幼根为初生结构,表皮细胞木栓化,保护性强,可防止根内部水分过度散失。维管柱中的髓主要用于储存水分和保护维管组织,因此发达的髓可以帮助提高植株的储水能力。在本研究中,旱生环境下翠芦莉根的皮层厚度显著低于水生环境,髓占维管束的比例显著高于水生环境。由此可说明旱生环境下翠芦莉通过减小皮层厚度来提升水分运输效率,并通过增加髓占维管束比例的方式来储藏水分,从而适应干旱。
植物茎内发达的维管组织、韧皮部、木质部和髓都具有很强的输导作用,从而使水分得到高效运输[11-12]。翠芦莉茎为四棱形,皮层的厚角组织可起到支持作用[13]。角质层作为一个屏障,能够有效地减缓植物体内水分的流失,茎的角质层越厚,抵抗干旱和强光的能力越强[14-15]。翠芦莉在旱生环境中茎的角质层较厚,抵抗干旱能力增强。旱生环境下翠芦莉的茎维管束厚度、初生韧皮部厚度、初生木质部厚度均高于水生环境,说明旱生环境下翠芦莉的茎具有更强的输导能力,水分运输效率较高。
叶片大面积暴露在环境中,是发生光合作用和呼吸作用的主要场所,易受到环境条件的影响而发生叶片结构特征上的变化[16]。因此,可以通过分析叶片的解剖结构来反映植物对不同环境的适应能力。植物体内大量的水分会通过叶片气孔在蒸腾作用中散失,气孔的规格以及密度都会影响蒸腾效率,因此,耐旱植物会表现出植物叶片气孔密度变大的典型特征[17-18]。旱生环境下翠芦莉的气孔密度较大,符合这一特征。同时,旱生环境中翠芦莉的叶片上角质层较厚,能够减少水分散失,增强保水能力。栅栏组织和海绵组织会影响植物抗旱性的强弱,植物抗旱性会随栅栏组织厚度的增加而增强,随海绵组织厚度增加而减弱[19]。发达的栅栏组织通常可以提高水分自输导组织至表皮的运输效率,降低蒸腾耗散;提高光能利用效率,避免高温和光照对植物的伤害[20]。因此,旱生环境下的翠芦莉栅栏组织较厚、叶片紧密度较高,能更有效地利用水分和光能,抗旱性更强。叶片结构疏松度是指海绵组织厚度占叶片厚度的比例,一般来说,疏松度越大,抗旱能力就越弱[21]。在水生环境下,翠芦莉的海绵组织厚度及叶片结构疏松度较大,说明在土壤水分含量较高的水生环境中,翠芦莉抗旱性相对减弱。栅海比大于1的植物通常被认为是典型的抗旱性植物[9]。旱生环境下翠芦莉栅海比大于1,属于抗旱性植物。叶脉结构不仅为叶片提供机械支撑、保障叶片生理机能正常运转,还负责将水分转运至叶片各细胞中。王树森等[22]认为发达的叶脉有利于植物在干旱环境中生存。旱生环境下翠芦莉的叶片主脉维管束结构要比水生环境更发达,从而能够更好地适应缺水环境。综上所述,在旱生环境下,翠芦莉的叶片解剖结构上主要表现为以较大的气孔密度,较厚的角质层厚度、栅栏组织厚度及更大的主脉维管束长度来适应高温、干旱环境。
变异系数法和可塑性指标可用于反映植物对周围环境的自我适应性,变异系数反映物种的潜在适应能力,而可塑性指标则反映物种克服自然环境异质化的能力,指标的数值愈大,表示植物适应自然环境的能力愈强[23]。因此,旱生环境中的翠芦莉叶片结构变异系数较大,即适应了变化性更多样的生境。在所有解剖结构中,主脉维管束长度的变异性和可塑性较高,表明主脉维管束在长度上的调节能力更强,更容易根据环境的变化做出改变,这在翠芦莉叶片适应不同生境中起着非常重要的作用,是翠芦莉能够适应环境的重要结构。2种生境下翠芦莉叶片各解剖结构的变异系数和可塑性指数的不同,说明翠芦莉适应于多样化生境的策略与其叶片解剖结构表现出的变异性与灵活性密切相关。
综上,翠芦莉在水生和旱生环境中生长良好,反映在营养器官结构上均有各自适应不同环境的策略,表明该植物生态幅较宽,同时能形成稳定的景观效益,是本气候区域中一种优秀的绿化植物,值得推广应用。
猜你喜欢
河北农业大学学报(2021年5期)2021-11-10 13:11:28
新疆农业科学(2020年9期)2020-10-13 12:37:42
中国现代医药杂志(2020年3期)2020-05-08 04:33:08
河南农业科学(2019年9期)2019-09-24 09:10:48
中国生物医学工程学报(2019年6期)2019-07-16 07:52:48
中外医疗(2016年15期)2016-12-01 04:25:39
环境科技(2016年4期)2016-11-08 12:18:58
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23 00:59:48
长江蔬菜(2014年1期)2014-03-11 15:09:48
计算机工程(2014年6期)2014-02-28 01:26:26
