
北冰洋太平洋扇区碳循环过程对环境变化响应的研究进展与展望
2022-11-03 08:48:48罗晓凡郑字佳
海洋科学进展 2022年4期
魏 皓,罗晓凡,祁 第,郑字佳,赵 伟
(1.天津大学 海洋科学与技术学院,天津 300072;2.集美大学 极地与海洋研究院,福建 厦门 361000)
人类活动不断排放二氧化碳(CO2),并在大气中积累,使得其浓度迅速攀升,并由此带来了一系列环境问题,如全球变暖和海洋酸化等[1]。在此背景下,提高海水对大气CO2的吸收,从而实现“碳中和”已成为保障人类社会可持续发展的必要措施。极区是全球主要的海洋碳汇区域,而北冰洋海冰覆盖减少可能会吸收更多的大气CO2,是潜在的碳增汇区域[2]。在北冰洋,海表吸收CO2除了受控于开阔海域大气CO2溶解外,其他复杂的碳循环过程,如水体中浮游植物生长固碳、颗粒有机物的沉积埋藏和总碳的深海封存等,均可大大提高海洋的碳汇效率[3]。探究“溶解泵” “生物泵”和“陆架泵”对环境变化的响应是认识北极碳汇过程及其调控机制的关键,是预测北冰洋碳汇的基础。
北冰洋太平洋扇区(图1)是北冰洋最重要的碳汇区域之一。流经白令海峡的太平洋入流水为楚科奇海输入了丰富的营养盐,使其具有很高的初级生产固碳能力[4],对北冰洋总固碳能力的贡献最高可达58%[5]。高初级生产力支撑着复杂的食物网,形成以楚科奇海南部和汉娜浅滩毗连海区两个生物热点区域,向底层输出大量有机碳,启动高生物量的大型底栖生物食物网[6]。具有高营养盐、高溶解无机碳(Dissolved Inorganic Carbon,DIC)和低pH 值特征的太平洋入流水[7-8]携带陆架生态系统产生的有机碳流出陆架后,进入陆坡和海盆次表层以进一步完成碳的深海封存,引起次表层酸化扩展[7],并可在不同垂直过程作用下对表层海-气CO2通量产生影响。
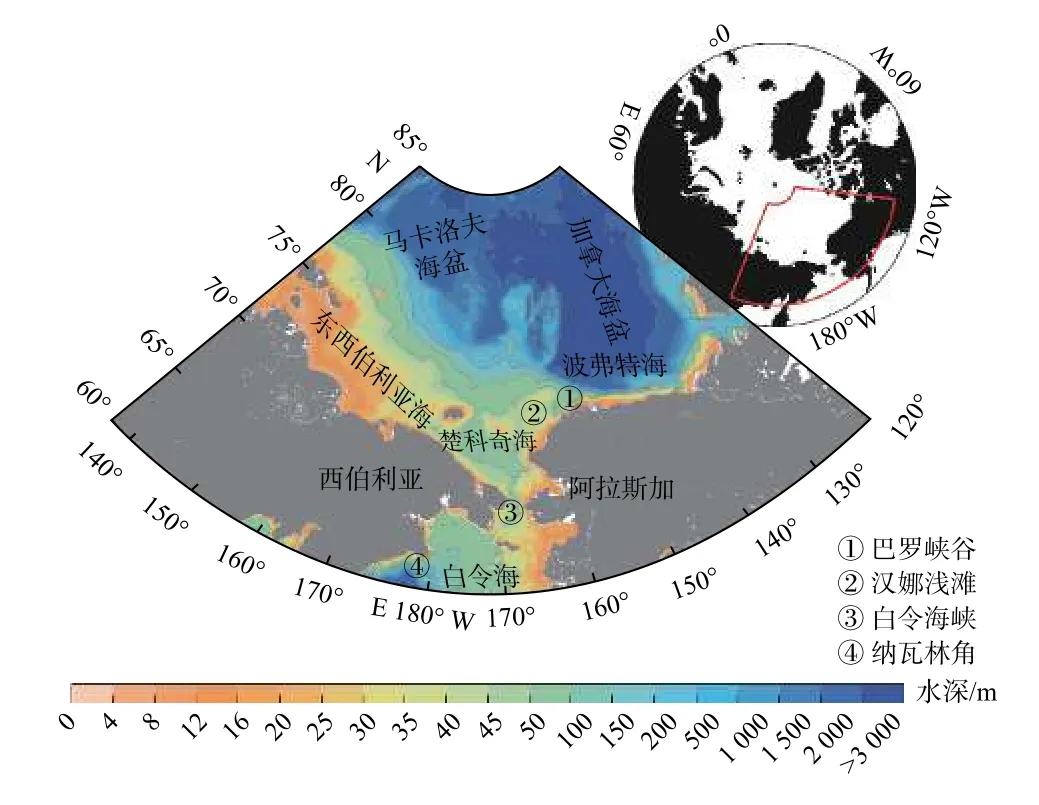
图1 北冰洋太平洋扇区地形及主要海区Fig.1 Topography and major seas in the Pacific Arctic Region
近20 a 来太平洋扇区环境变化剧烈,其海冰覆盖面积下降速率明显高于大西洋扇区[9-10],太平洋入流水通量增加20%[11],波弗特高压增强且西移南下,整个气候系统在2007 年前后发生跃变[12],由此造成了太平洋扇区环流和物质输运的大幅度调整[13]。碳循环过程对环境变化的响应是复杂的,存在很大的不确定性和时空差异,需要基于观测和数值模式的研究给出定量解释。在国家自然科学基金委重点项目和科技部重点研发国际合作计划等支持下,我国学者基于10余a 北极科考和海洋-海冰-生物地化耦合的数值模式开展研究,已跻身国际先进行列:整理并订正了所有中国北极科考数据和近30 a 的国际碳数据集,分析了初级生产和海冰变化对海-气CO2通量的影响,提出环流变化是太平洋扇区海盆区域酸化扩展的决定因素,目前这一观点已被国际公认[7];构建了包含北大西洋、北太平洋和整个北冰洋的海洋-海冰-生态系统-碳循环耦合模式(North Atlantic and Pacific Oceans and the Arctic Ocean-Biogeochemical Model,NAPA-BGC)[14-15],评估结果表明,自由运行(无同化)20 余a 的NAPA-BGC 模式达到国际北冰洋模式比较计划[16-17]相同水平,区域海碳循环模式的建立推动了我国对于北冰洋太平洋扇区碳循环响应机理研究。基于此,本文对国内外相关工作进行概述,并对未来研究需要聚焦的关键科学问题予以展望。
1 基于观测的太平洋扇区夏季海-气CO2 通量变化与影响因素
海-气CO2分压差决定了海洋碳源汇格局和强度,表层海水CO2分压(pCO2)增加(减小)导致碳汇能力的减弱(增强)。在年代际尺度上,世界大洋海表pCO2几乎都在增加,包括中低纬度海区以及南大洋[18-20]。在北冰洋,早期观测研究指出高初级生产力的陆架-陆坡区域表层海水pCO2持续较低,是碳汇增强海区[21]。而海盆无冰区的最新观测发现,由于快速的海-气交换和增温变暖,海表pCO2值接近大气CO2水平,碳汇能力没有显著增加[22-23]。整合北冰洋20 余a 走航观测数据,自然资源部第三海洋研究所碳化学团队给出了北冰洋太平洋扇区夏季海表pCO2的年代际变化及调控机理[24],指出在1994—2017 年,受暖化和海冰消融的影响,加拿大海盆夏季表层海水pCO2的 增加速度是大气的2 倍,因而海-气CO2梯度(ΔpCO2)迅速减小,碳汇减弱并可能在20 a 内降至接近于0;相比之下,楚科奇海陆架、陆坡表层海水pCO2并没有显著增加,该区不断增强的生物吸收作用促使海表pCO2保持在较低水平[25-26],未来随着大气CO2的增加楚科奇海碳汇可能进一步增强(图2),Tu 等[27]用遥感反演产品对整个楚科奇海夏季碳通量研究得到相同结论。
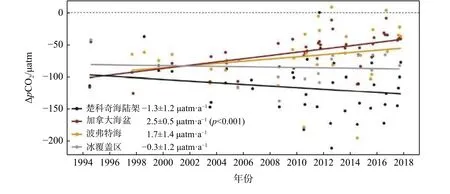
图2 1994—2017 年间楚科奇海陆架、加拿大海盆和波弗特海夏季Δ pCO2的变化趋势Fig.2 Trends of Δ pCO2 in the Chukchi Shelf,Canada Basin and Beaufort Sea during the year of 1994—2017
Ouyang 等[26]基于北极科考2016 和2018 年夏季代表净群落初级生产力(Net Community Productivity,NCP)的海表O2/Ar 和pCO2走航观测数据,进一步细化了不同海冰和营养盐供给条件下NCP 和海-气CO2通量的关系(图3):在海冰覆盖区,由于海冰阻隔了气体交换,海-气CO2通量较小,微弱的初级生产导致了较大的海-气CO2浓度梯度;在冰边缘区,NCP 和CO2通量之间具有显著的线性相关关系,表明在海冰融化初期,海-气CO2通量主要由生物吸收过程所调控;无冰水域,NCP 和CO2通量的耦合因营养盐供应的差异而呈现区域性的变化:在寡营养盐的开阔海盆中,NCP 和CO2通量之间不存在明显的相关性,微弱的NCP 和较低的海-气CO2梯度限制了海盆的碳汇潜力,使之无法成为重要的CO2汇,而营养盐丰富的楚科奇海陆架则具有较强的NCP 和碳汇作用。
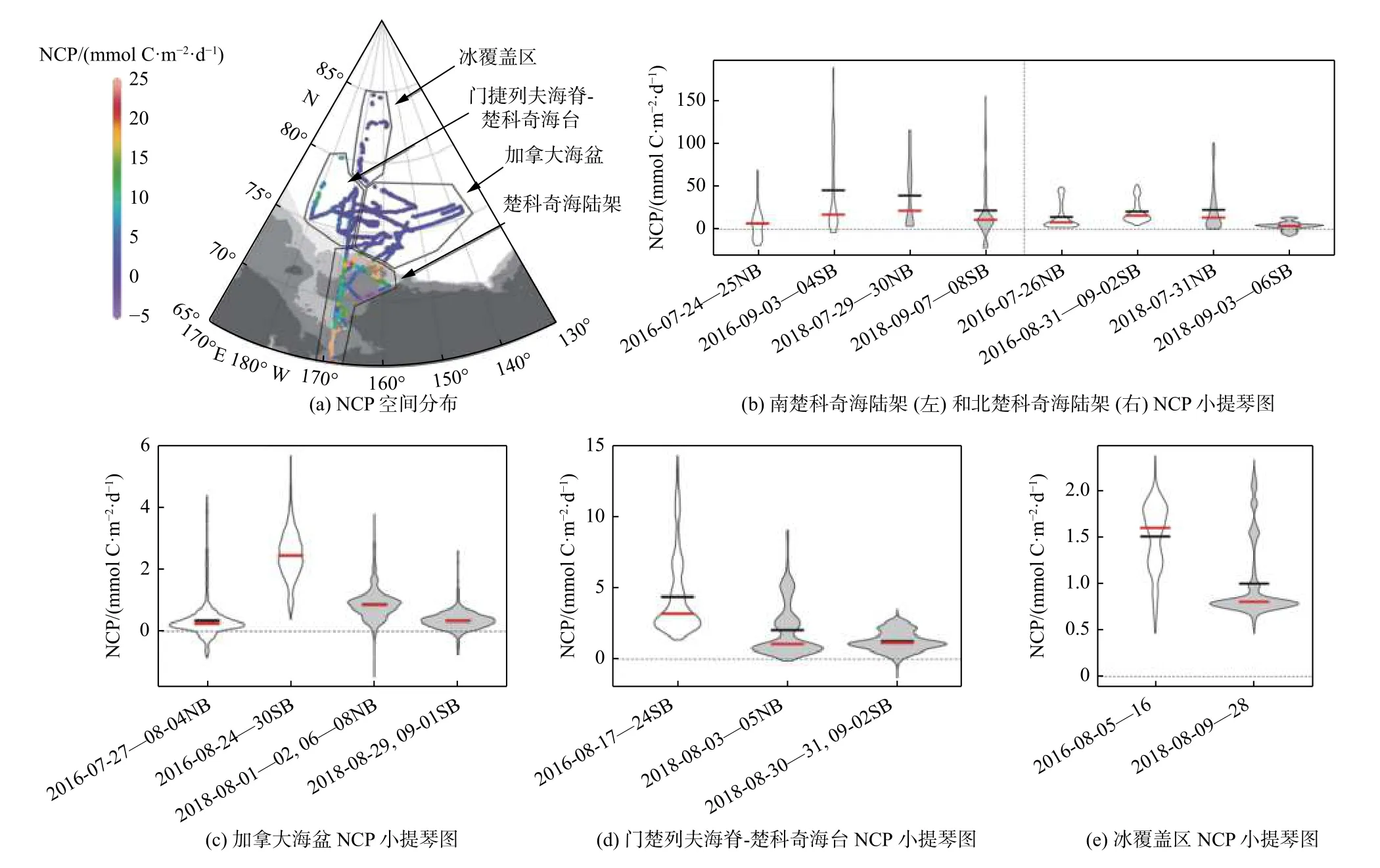
图3 北冰洋太平洋扇区NCP 的估算Fig.3 Estimations of net primary production(NCP)in the Pacific Arctic Region
2 楚科奇海海-气CO2 通量季节、年际变化及控制机制的定量认识
夏季观测数据揭示了北冰洋生物泵作用的重要性,营养盐供给匮乏的海盆区碳汇能力无法随着海冰融化而持续增加。在年均碳汇通量中,夏季占多大份额?碳汇能力在季节、年际和气候状态转换中有什么样的响应?对哪些环境要素变化响应?整个北冰洋的碳循环是一个复杂过程,仅靠夏季表层观测还不足以回答上述问题,区域海-气CO2通量季节、年际变化及控制机制还需要可靠的数值模式及准确的碳收支估算来定量认识。
魏皓等[15]和Zheng 等[15]构建了北冰洋海洋-海冰-生态系统-碳循环全耦合模式(NAPA-BGC),重点解析了楚科奇海陆架海-气CO2通量的季节及年际变化机理。Zheng 等[28]利用现场观测数据对模式模拟的海表DIC 和pCO2进行了严格的评估比较(图4 和图5),结果表明该模式是目前为止已发表模式研究中对楚科奇海碳循环过程模拟最为准确的3 维模式,模式抓住了太平洋入流通过狭窄海峡(白令海峡、巴罗峡谷)补充营养物质对楚科奇海陆架及下游太平洋扇区碳汇的调控作用,这是低分辨率地球系统模式很难反映的过程。
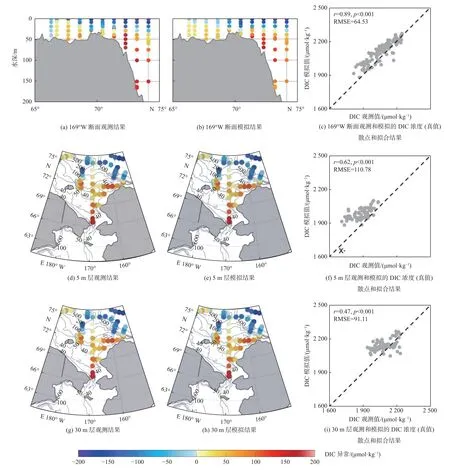
图4 2008 年夏季航次(CHINARE08 和ISSS08)期间楚科奇海169°W 断面、5 m 层和30 m 层观测和模拟的溶解无机碳异常分布Fig.4 Distributions of observed and simulated dissolved inorganic carbon(DIC)anomalies along the 169°W section,at the 5 m and 30 m depth layers in the Chukchi Sea during the summer 2008 cruises(CHINARE08 and ISSS08)
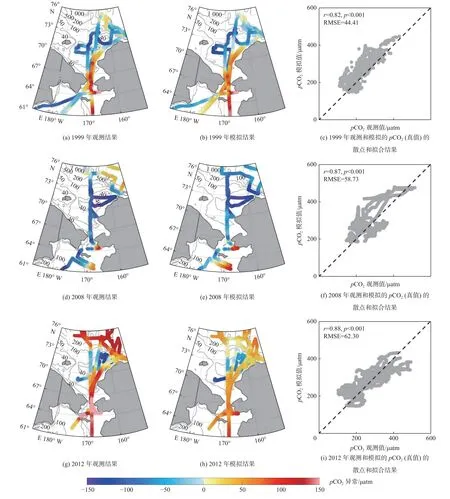
图5 1999 年、2008 年和2012 年夏季观测和模拟的楚科奇海海表CO2 分压( pCO2)异常分布Fig.5 Distributions of observed and simulated partial pressure of CO2( pCO2)anomalies in the summer of 1999,2008,and 2012 in the Chukchi Sea
Zheng 等[28]模式结果表明,楚科奇海陆架全年表现为CO2净汇,其海-气CO2通量的季节变化显著(图6红色实线)。6 月,海冰消退导致开阔水域面积增大,楚科奇海迅速成为1 个较强的碳汇;7—10 月,CO2通量维持在较高水平(<-6.93 mmol·m-2·d-1),尤其是楚科奇海北部区域,模式获得的CO2通量空间分布与观测一致,我国北极考察均在8—9 月进行,此时碳通量约占全年40%;11—12 月,随着海冰快速冻结,海-气CO2通量迅速减小直至为0 并维持到次年5 月。将海-气CO2通量的季节变化影响因子分解为气体交换系数(风速主导)(图6 深红色)、海-气CO2分压差(图6 绿色)和海冰密集度(图6 浅蓝色)的季节变化三个分量,分析了其调控机制。海冰融冻(或开阔水域面积)的季节变化主控了碳通量的季节变化,但在6—7 月的融冰期,初级生产吸收CO2所致海-气CO2分压差的增大对碳汇起到了重要作用,在10—11 月结冰期,秋季大风对于楚科奇海尚存的水面吸收大气CO2有较大的贡献。海冰覆盖的1—5 月,生物降解、群落净呼吸使表层海水pCO2增加,海-气CO2分压差小于年均值,起到减小碳汇的作用。
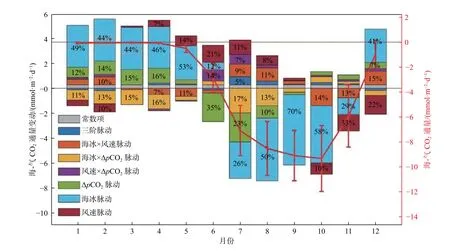
图6 楚科奇海海-气CO2 通量季节变化影响因素Fig.6 Factors affecting the seasonal air-sea CO2 flux variation in the Chukchi Sea
Zheng 等[28]模拟的1998—2015 年多年平均碳通量分布体现了1 个事实:楚科奇海陆架(图7b 黑线框内)是太平洋扇区主要碳汇区,海盆源汇作用极弱、河口附近则为碳源(图7b)。楚科奇海海-气CO2通量多年平均值约为10 Tg C·a-1,自1998 年以来以0.28 Tg C·a-1速率增加(图8a 黑线);初级生产和开阔水域面积的增加主控了楚科奇海融冰期(6—7 月)(图7a 绿线)和开阔水域期(8—9 月)(图7a 蓝线)CO2通量的显著增长趋势,其增速分别为0.08 Tg C·a-1和0.11 Tg C·a-1,这2 个时期(6—9 月)CO2通量的增加趋势决定了楚科奇海全年总CO2通量的长期增长趋势;而结冰期(10—11 月),CO2通量受开阔水域面积和局地风速年间巨变影响,年际波动打破了上升趋势(图7a 红线),它主控了楚科奇海陆架总CO2通量的年际变化(r=0.86,p<0.01)。
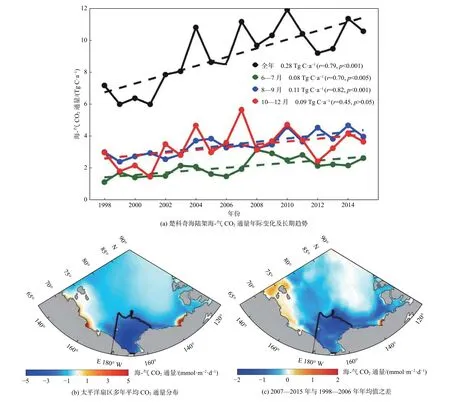
图7 楚科奇海陆架海-气CO2 通量年际变化及长期趋势、太平洋扇区多年平均CO2 通量分布和2007—2015 年与1998—2006 年年均值之差Fig.7 Interannual variability and long-term trend of air-sea CO2 flux over the Chukchi Sea shelf,distribution of multi-year averaged air-sea CO2 flux in the Pacific Arctic Region and changes between 2007—2015 and 1998—2006
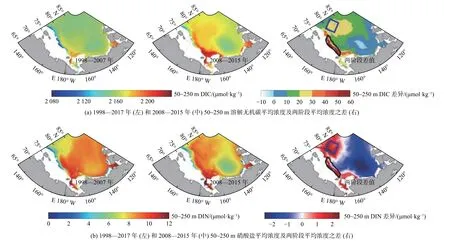
图8 1998—2007 年和2008—2015 年50~250 m 溶解无机碳和硝酸盐平均浓度及两阶段平均浓度之差Fig.8 DIC and nitrate concentration averaged within 50—250 m water column during 1998—2007,2008—2015 and the differences between the two periods
模拟结果展现了太平洋扇区气候转换前(1998—2006 年)和后(2007—2015 年)状态之间显著的碳吸收差异(图7c),表现为强源/汇各自增强,寡营养海盆区对气候变化并不敏感。陆架区初级生产增加对增汇贡献已是定论,陆坡和海盆区除海冰状态变化影响外,营养盐供给变化的贡献还需继续量化。
3 太平洋水在陆坡和海盆区输运变化及其对海洋碳汇作用的影响
高DIC 的太平洋水流经楚科奇海陆架底层时进一步吸收有机物再矿化释放的额外CO2[5,7,30],流出陆架后继续进入陆坡和海盆区,占据北冰洋太平洋扇区次表层[31-32],在海盆内次表层(50~250 m)形成DIC 最大值层[33-34]。Chen[35]基于观测数据研究发现1994 年马卡洛夫海盆次表层没有DIC 最大值层,而2010 年次表层太平洋水占比达到50%、存在DIC 最大值层,推测这可能是环流的改变使得太平洋水扩展至此带来的。Qi等[7]基于150°W 至170°W 多个经向断面多年观测数据研究发现次表层酸性水体也从1990s 到2010 年不断向高纬海盆扩展并且在垂向上有所加深。太平洋水同时也是高营养盐水体,在进入海盆后会在上盐跃层中形成营养盐的最大值层[36]。观测发现马卡洛夫海盆1996 年上盐跃层中不存在营养盐最大值层,2014 年则出现了营养盐最大值层[37]。可见太平洋扇区陆坡和海盆在2007 年前后营养盐供给和DIC 分布均发生了气候态转型,这会引起其碳汇能力怎样的响应呢?
Chu 等[29]利用NAPA-BGC 模式,揭示了太平洋扇区是次表层DIC 和营养盐对北冰洋环境快速变化响应的敏感区,解析了变化格局的主控过程及太平洋水输入在敏感区次表层-表层的界面交换对表层碳汇效率的影响。模式结果表明:太平洋水分布范围在气候转变的前后2 个阶段(1998—2007 年和2008—2015 年)发生很大变化,次表层DIC 和硝酸盐(DIN)浓度高值区在向高纬扩展,最远至85°N 左右,同时也向深处扩展,最深到达300 m 左右;2 个时段次表层(50~250 m)DIC 和DIN 浓度差异均在马卡洛夫海盆和楚科奇-东西伯利亚陆架坡折区最为显著(图8)。波弗特流涡对波弗特高压南下、西移的响应使得楚科奇-东西伯利亚陆坡流增强,同时也有利于物质向陆坡和马卡洛夫海盆输运。然而,环流水平输运带来陆坡区的营养盐供给增加同时发生的表层和次表层,海盆区仅发生在次表层,垂直交换带来截然不同的效果:陆坡区表层初级生产提升、碳汇增加,DIC、DIN 由表层向次表层输运,同时碎屑沉降增加导致次表层有机物再矿化增强,进一步加剧陆坡次表层DIC 浓度增加;马卡洛夫海盆区海表碳汇增加并不明显(图7c),因为其表层营养盐是通过次表层与表层(0~50 m)的垂向向上交换增加的,但同时DIC 也向表层输运,两物质通量的比值(DIC∶DIN)远大于Redfield 碳氮比(RC∶N),对上层海洋吸收CO2能力产生抑制作用,营养盐供给差异在气候态碳汇变化中起了重要作用(图9)。可见海盆区增汇潜力较弱有复杂的物理过程影响,同时可推测只有在寡营养海区增加表层营养盐而不同时增加DIC 的地球工程才能起到增汇作用。
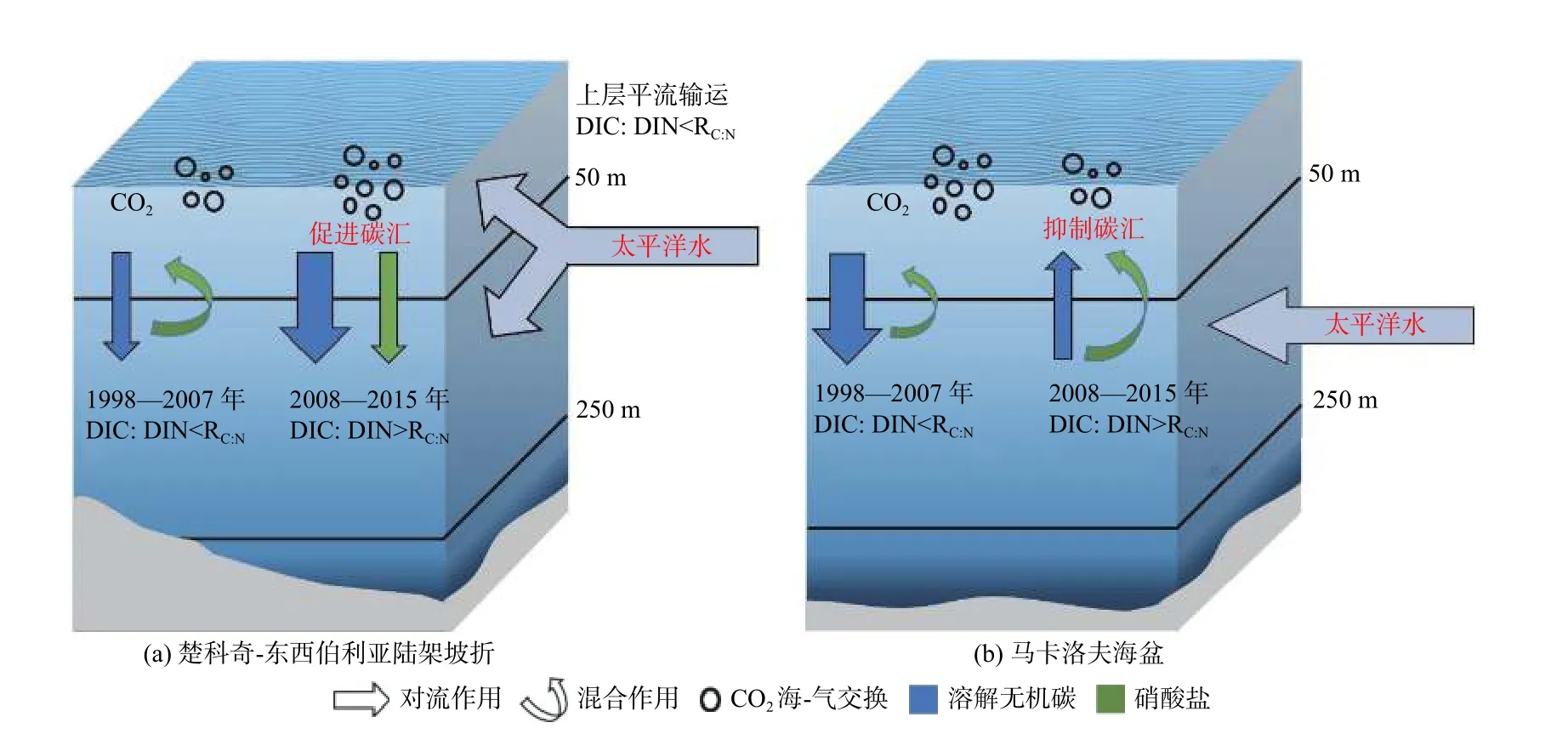
图9 楚科奇-东西伯利亚陆架坡折和马卡洛夫海盆次表层与表层DIC 和DIN 交换概念模型Fig.9 Schematic diagrams of DIC and DIN exchanges between surface and subsurface in the Chukchi-East Siberian Shelfbreak and the Makarov Basin
4 楚科奇海营养盐供给与初级生产维持及年际变化机制
常言道“太平洋水哺育了北冰洋”。NAPA-BGC 模式结果[38]进一步明确了白令海纳瓦林角附近陆架/陆坡交换-安纳德尔流流经海区-白令海峡西侧是营养盐向楚科奇海输运的主要通道,指出纳瓦林角附近陆架/陆坡交换(CN 断面,图10)对硝酸盐向极输运有调控作用,这个交换通量与白令海峡冬(2—3 月)、夏季(7—8 月)硝酸盐通量年际变化的相关关系奠定了营养盐向极输运通量的季节预报基础。对夏季白令海陆架营养盐输运通道的硝酸盐收支估算表明:生物吸收作用对硝酸盐向极补充通量年际变化的影响较小,平流输运作用占优。白令海海洋环流年际差异主要是由阿留申低压位置决定的,随着阿留申低压中心的加深及向西偏移,富含营养盐的安娜德尔流大部分会在安娜德尔湾内发生逆转而无法北上,从而引起白令海峡营养盐通量的急剧减少,这个大气环流异常引起下游楚科奇海整个生态系统到碳汇能力的巨大变化,确实是“太平洋打个喷嚏楚科奇海就感冒”的关联。
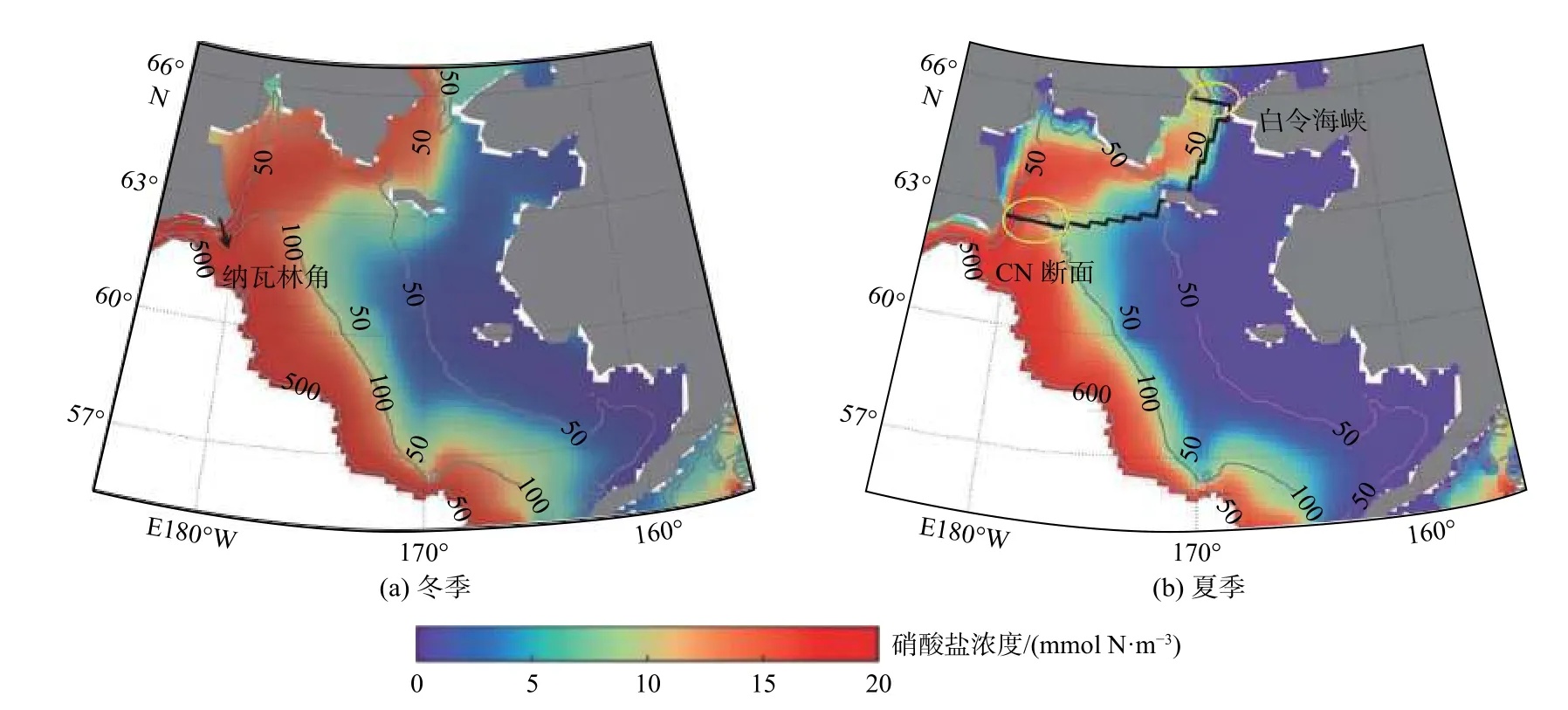
图10 1998—2015 年白令海冬季与夏季500 m 深度平均硝酸盐分布Fig.10 Depth-averaged simulated nitrate concentration in the upper 500 m water column in the Bering Sea averaged for winter and summer seasons over 1998—2015
楚科奇海浮游植物具有较长的生长期,Arrigo 和Dijken[39]指出1998—2009 年间楚科奇海水华时长平均为119 天,而Hill 和Cota[40]认为楚科奇海生长期达150 d。楚科奇海初级生产的年均值在55~720 g C·m-2·a-1之间[4,41-42],伴随着海冰对光限制的改变,楚科奇海初级生产有着显著的季节变化。5 月初,楚科奇海的初级生产开始启动,其值普遍小于300 mg C·m-2·d-1[40],近年的观测发现在5—6 月,楚科奇海存在冰下浮游植物水华[43-44]。夏季(7—8 月),由于表层营养盐被春季水华大量消耗,楚科奇海初级生产向深层发展,最大值出现在次表层25~30 m 之间,与营养盐跃层上界一致[40],且生产力处于相对较高水平,平均值在700~1 400 mg C·m-2·d-1之间[4,45]。秋季(9—10 月),垂向混合作用加强,表层营养盐逐渐得到补充,表层出现强度较弱的秋季水华,总初级生产在255~578 mg C·m-2·d-1之间[46-47]。已有研究强调了太平洋冬季水对东北楚科奇海生态系统的重要性[48-50],然而楚科奇海的初级生产存在较大的空间差异,形成了以楚科奇海南部和汉娜浅滩毗连海区为中心的南北两个生物热点区。5 月南、北热点区初级生产分别可达2 166 mg C·m-2·d-1和407 mg·C·m-2·d-1,7 月更是分别高达3 015 mg C·m-2·d-1和2 016 mg C·m-2·d-1[4],8 月南、北热点区浮游植物平均固碳速率分别为1 630 mg C·m-2·d-1和180 mg C·m-2·d-1[51],9 月南、北热点区初级生产平均值分别为437 mg C·m-2·d-1和126 mg C·m-2·d-1,年均值分别为208 g·C·m-2·a-1和173 g C·m-2·a-1[4]。总体而言,楚科奇海南部生物热点区初级生产高于北部生物热点区[6]。
楚科奇海在整个生长季都维持着相对较高的初级生产,正是这种高生产使楚科奇海发挥着高效的生物泵作用,Zheng 等[14]通过分析NAPA-BGC 模式结果,揭示了楚科奇海2 个生物热点区高初级生产的维持机制。从水团角度的分析结果表明,楚科奇海南北两个生物热点区的初级生产启动均以太平洋冬季水携带的营养盐为基础,而生长季内初级生产的主要贡献者分别为白令海夏季水和太平洋冬季水。南北两个生物热点区生长季真光层内的硝酸盐收支估算结果表明(图11):南区真光层内的硝酸盐主要来源于平流作用输运的白令海夏季水;在北区,受地形诱导的环流结构使得富营养盐的太平洋冬季水可较久地存留于汉娜浅滩附近,待上层水体营养盐被春季初级生产消耗后,垂向混合作用将存留太平洋冬季水内的硝酸盐持续补充至真光层,以此维持着汉娜浅滩附近夏秋季的初级生产,形成该区长达110 d 的水华期。模式结果显示,楚科奇海在1998—2015 年间春季(6—7 月)初级生产(图12 绿线)和碳汇(图7a 绿线)均呈现显著增加的趋势,且前者为因后者为果[28]。考虑到营养盐自白令海峡进入楚科奇海再向下游输运的时间[32],对楚科奇海春季初级生产与白令海峡2—7 月硝酸盐通量年际变化进行相关分析,发现两者的相关性非常显著(r=0.90,p<0.01),高于初级生产与春季楚科奇海营养盐浓度的相关性(r=0.79,p<0.01)(图12),也就是说2—7 月太平洋水营养盐通量决定了春季楚科奇海生物固碳能力,调控了海-气CO2的通量。
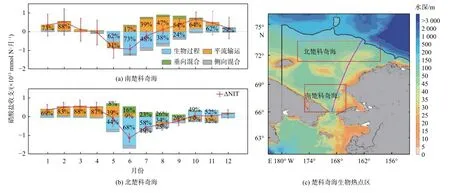
图11 楚科奇海南北两个生物热点区真光层内的硝酸盐收支Fig.11 Nitrate budgets in the euphotic zones of the southern and northern biological hotspots in the Chukchi Sea
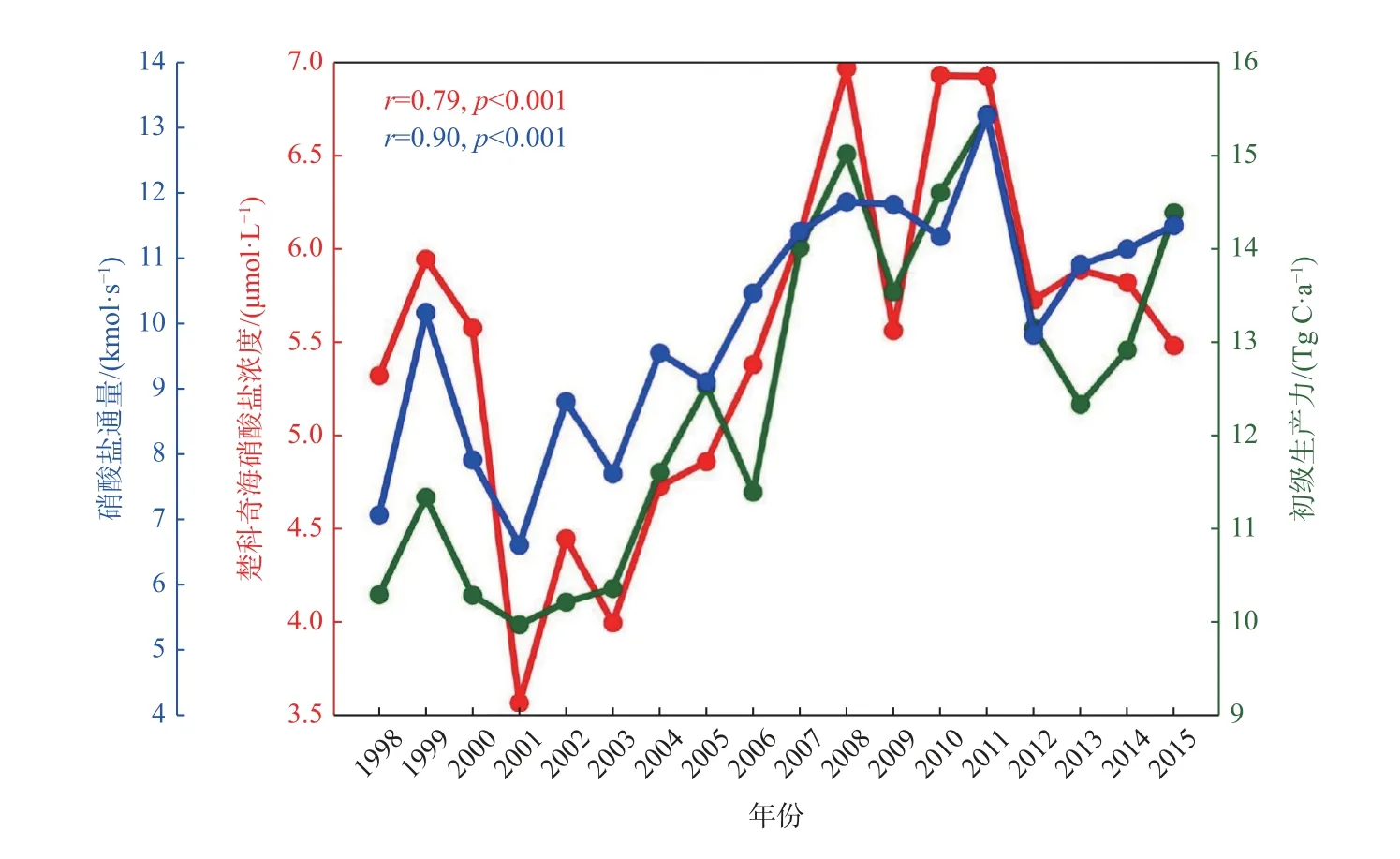
图12 白令海峡2—7 月硝酸盐通量与楚科奇海春季(6—7 月)硝酸盐浓度和初级生产力Fig.12 Nitrate flux through the Bering Strait during February—July and spring(June—July)nitrate concentration and primary production in the Chukchi Sea
5 太平洋扇区有机碳输出
生物泵增汇的关键在于大量颗粒有机碳快速移出真光层,进入沉积埋藏或者进入深海封存。长时间序列观测的难度阻碍了这个方向的发展,沉积物捕获器是近年来开始使用的观测手段,极大推动了人们对太平洋扇区有机碳输出响应环境变化的认识。基于国际合作,2015—2016 年布放在东北楚科奇海的汉娜浅滩南侧CEO 站的沉积物捕获器(37 m)首次获取了太平洋扇区浅海陆架的周年连续观测数据,观测期间颗粒有机碳(Particulate Organic Carbon,POC)垂向通量为145 g C·m-2·a-1,春夏季POC 通量的高值主要由硅藻通量贡献,秋季依然存在较高的有机碳输出[51]。阿拉斯加大学Danielson 团队在白令海峡附近海域布放了2 个锚系(~36 m)和7 个漂流沉积物捕获器,受白令陆架水和安娜德尔水高营养盐水体影响,2017—2018 年2 个锚系站位的POC 通量分别为204 和215 g C·m-2·a-1,这是全球海洋中有记录以来的最高通量之一,且该海域初级生产的输出效率极高,平均为82%[52]。Lalande 等[53]综合2017—2019 年北白令海和楚科奇海3 个生物热点区站位的沉积物捕获器数据,在融冰早的年份发现高叶绿素和硅藻通量的持续时长减少,生物泵效率较低,初级生产的能量主要进入水层生态系统而非底栖生态系。2017—2018 年,韩国团队[54]在楚科奇-东西伯利亚陆坡分别布放了沉积物捕获器,楚科奇海陆坡站位(325 m)的POC 通量为2.1 g C·m-2·a-1且无明显年际差异,东西伯利亚陆坡站位115 m 和335 m 层的POC 通量分别为2.3 和2.0 g C·m-2·a-1,但由于高营养盐陆架水的入侵,2018 年8 月的POC 通量比2017 年8 月高1 个数量级。日本Watanabe 团队在楚科奇海台南部NAP站和CAP 站布放的沉积物捕获器中发现了大量的陆源物质,认为楚科奇海陆坡流和中尺度涡的发展及移动对陆架物质的输运会影响楚科奇海台碳输出[55-57]。我国自然资源部海洋二所陈建芳团队自2008 年起在北冰洋布放沉积物捕获器开展长期监测研究,在参与国际北极漂流冰站计划(Multidisciplinary drifting Observatory for the Study of Arctic Climate,MOSAiC)期间布放了我国首套北极冰基沉积物捕获器,通过分析长时间序列颗粒物通量、浮游生物群落组成、生物标志物等参数取得了丰富的前沿认识[58-61]。在北风海脊附近布放的沉积物捕获器(870 m)数据显示,夏季海冰开阔期颗粒有机碳通量较高(8.78 mg C·m-2·d-1),而秋冬季有冰期颗粒有机碳通量较小(1.46 mg C·m-2·d-1),观测期间POC 总通量为1.9 g C·m-2·a-1;同时他们还发现2009 年颗粒有机碳通量高于2008 年,认为这与海冰融化早、开阔时期长(生长期延长)有关[58]。由此可见,陆架存在极高的POC 沉降通量,这是对陆架高生产力的体现;在陆坡和海盆区,POC 输出通量较低。海冰、营养盐、环流结构等环境因素的变化都会深刻影响太平洋扇区有机碳输出,需要进行持续的长期监测。目前的研究主要聚焦在有机碳移出真光层,进入沉积埋藏或者进入深海封存的相关研究较少[62-66],这限制了人们对太平洋扇区碳汇能力的全面理解。未来需密切结合观测和模式结果,估算太平洋扇区陆架有机碳沉积埋藏通量和向海盆输出通量,深入认识有机碳输出对环境变化的响应。
6 展 望
北冰洋太平洋扇区是海冰快速消退、气候系统转换、环境剧烈变化的区域,在不远的将来,“生物泵”“陆架泵”效率提高会带来陆架、陆坡区自然碳汇能力的提升。海盆区由于生物泵作用较弱,碳汇能力提升并不显著,加之受到酸化的严重限制[67],海冰在高纬海区的进一步消退、甚至30 a 后夏季无冰,都不可能同比提升太平洋扇区对CO2的吸收能力。同样,楚科奇海陆架海底层也在不断酸化[68],我们对于海冰消退、初级生产增加带来的碳汇红利依然不能过分高估。
楚科奇海是巨大的CO2汇,每年可从大气中固定近1 000 万t 碳,以区区55 万km2面积,固碳量可达整个北冰洋太平洋扇区(面积495 万km2)的44% ± 6%。每年沉降到海底的有机碎屑启动了楚科奇海庞大的底栖生物群落的生命活动,并由此支撑了个体巨大的食底栖哺乳动物(如海狮、海豹、海牛等),再通过复杂的生物地球化学循环而在沉积地层中埋藏,这部分碳埋藏通量有待评估,楚科奇海陆架向深海的碳输出通量及“陆架泵”运作机制还需深入理解,观测技术的突破为有机碳输出研究带来曙光。
楚科奇海因太平洋的营养补充而具有极高的初级生产能力,其碳汇增加的结论能否推广到北冰洋其他陆架海区还不确定。东西伯利亚海、拉普捷夫海会受到陆地河流、冰冻圈融化入海携带营养盐(硝酸盐)、DIC(含甲烷)的影响,这些因素叠加在海冰融化背景上使其碳汇变化更加复杂,但因观测数据的缺失,目前还没有定论。喀拉海和巴伦支海则受河流和大西洋水影响,其中巴伦支海因大西洋水缺硅(Si)而呈现微型浮游植物占优的生态特征,固碳能力小于硅藻占优的太平洋扇区[69],目前尚缺乏北冰洋快速环境变化背景下整个生态系统结构响应及其对海洋碳汇能力的影响认识,应开展太平洋扇区和大西洋扇区陆架、陆坡、海盆固碳能力对环境响应的对比研究,改善北冰洋碳汇的预测能力。波弗特海淡水增加与向北欧海泄漏被认为是影响大西洋翻转环流的关键,目前是气候变化的热点话题,这个变化是否与北冰洋碳汇能力变化有关?北冰洋是全球海洋酸化的领头羊,酸化背景下北冰洋碳汇能力变化的定量研究需要关注。当然,北冰洋碳汇能力变化在全球气候系统的反馈一直是学界关注的热点,毕竟寻找自然碳汇、实现“碳中和”以减缓暖化是目标,因此评估北冰洋碳汇能力增加在减缓气候变暖中的作用是当务之急,人为提高碳汇能力的地球工程可行性探究也进入大众视野。
猜你喜欢
沉积学报(2023年1期)2023-04-29 18:57:49
海洋通报(2022年2期)2022-06-30 06:07:04
中国新闻周刊(2021年14期)2021-04-28 23:56:08
阅读时代(2020年1期)2020-09-10 07:22:44
石油钻探技术(2020年3期)2020-08-24 07:28:00
海洋学报(2020年7期)2020-08-06 09:29:24
国外核新闻(2020年7期)2020-03-14 14:54:04
海洋信息技术与应用(2017年2期)2017-06-21 19:51:29
中国科技博览(2016年25期)2016-12-20 18:28:41
应用海洋学学报(2014年1期)2014-11-22 07:17:36
