
基于ISSR分子标记的枫香遗传多样性分析
2022-11-01 04:20黄旭萍陈孝丑吴明晶翁建宇陈发兴
福建农林大学学报(自然科学版) 2022年4期
黄 敏, 黄旭萍, 陈孝丑, 吴明晶, 翁建宇, 陈发兴
(1.福建农林大学园艺学院, 福建 福州 350007;2.福州植物园, 福建 福州 350012;3.福建省将乐国有林场, 福建 三明 353300)
枫香(LiquidambarformosanaHance)为金缕梅科枫香树属落叶乔木,主要分布于秦岭和淮河以南,且为福建省重要的乡土彩叶树种,常用于行道绿化和园林建设等[1-2].枫香树不仅具有观赏价值[3-4]、经济价值[5]和药用价值[6-7],还能消除有害气体,改善气体环境[8].
彩叶树种对于城市景配色和美丽乡村建设具有重要意义,目前我国彩叶树种品种较少.根据国外城市园林发展法[9],我国未来对种苗的需求必然会转向特色种苗,这将促进彩叶树种产业的发展.而枫香不仅色彩鲜艳、叶子美观,而且用途广泛、经济效益高,具有良好的发展前景.ISSR分子标记的主要特点是可以在目标靶标序列未知的条件下使用,拥有条带清晰明亮、多态性丰富、稳定性强、重复性好、简便易于操作、价格比较低廉、产物特异性强等优点[10],已被广泛应用于植物遗传多样性及亲缘关系分析[11-13]、种质资源的保护与鉴定[14]、遗传结构[15]等研究.林骥光等[16]利用ISSR分子标记技术对12份枫香种质资源进行分析,其中4号样本和11号样本的遗传相似度最高.毕泉鑫等[17]对浙江省自然种群枫香进行遗传多样性研究,将5个居群分为两大类群.对枫香种质资源的研究主要集中在繁殖技术[18-19]、造林[20]和材质[21]等,而对彩叶观赏树种的遗传关系研究较少.本试验利用ISSR分子标记技术对来自福建、江西和安徽3个省份的137份彩叶观赏价值高的枫香样品进行亲缘关系及遗传多样性分析,探索不同居群间枫香的亲缘关系,并从分子水平分析不同居群枫香的遗传特点,为彩叶枫香种质资源的收集和优良彩叶观赏价值新品种的选育提供参考.
1 材料与方法
1.1 试验材料
从福建省三明市将乐国有林场收集到的4 178株枫香种质资源中,选出137份秋季呈色良好的枫香种质,样品编号如表1.对选中单株的东南西北4个方位各取一个枝条作标记,从枝条顶端往下数第4片叶片,采集健康枫香嫩叶,放在装有冰袋的采样贮藏箱中保存.用超纯水将叶片表面所带的污染物洗干净后晾干,去除主要叶脉并剪碎后装入离心管,放在-80 ℃冰箱保存备用.
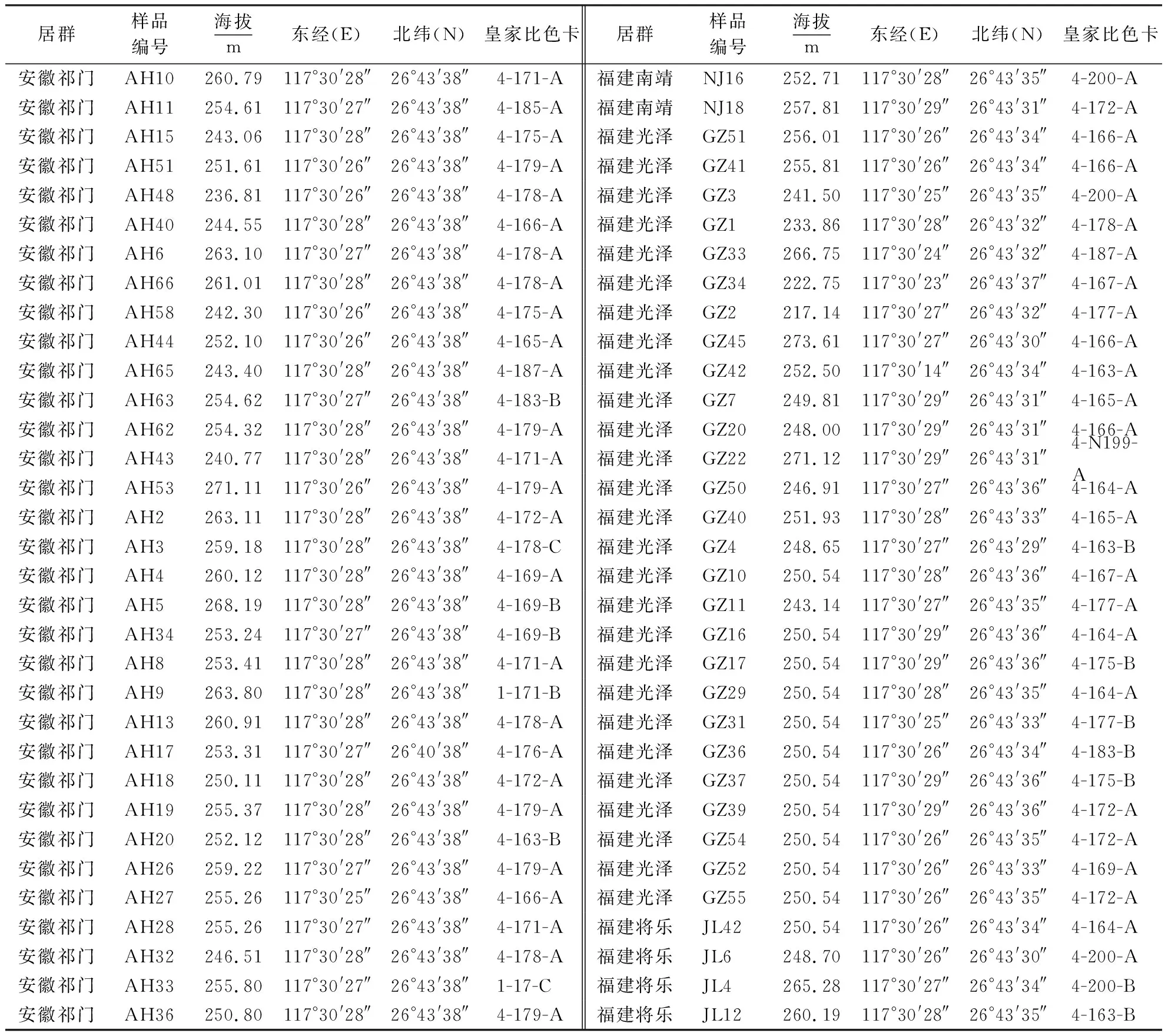
表1 137份枫香样品信息Table 1 Information of 137 L.formosana samples
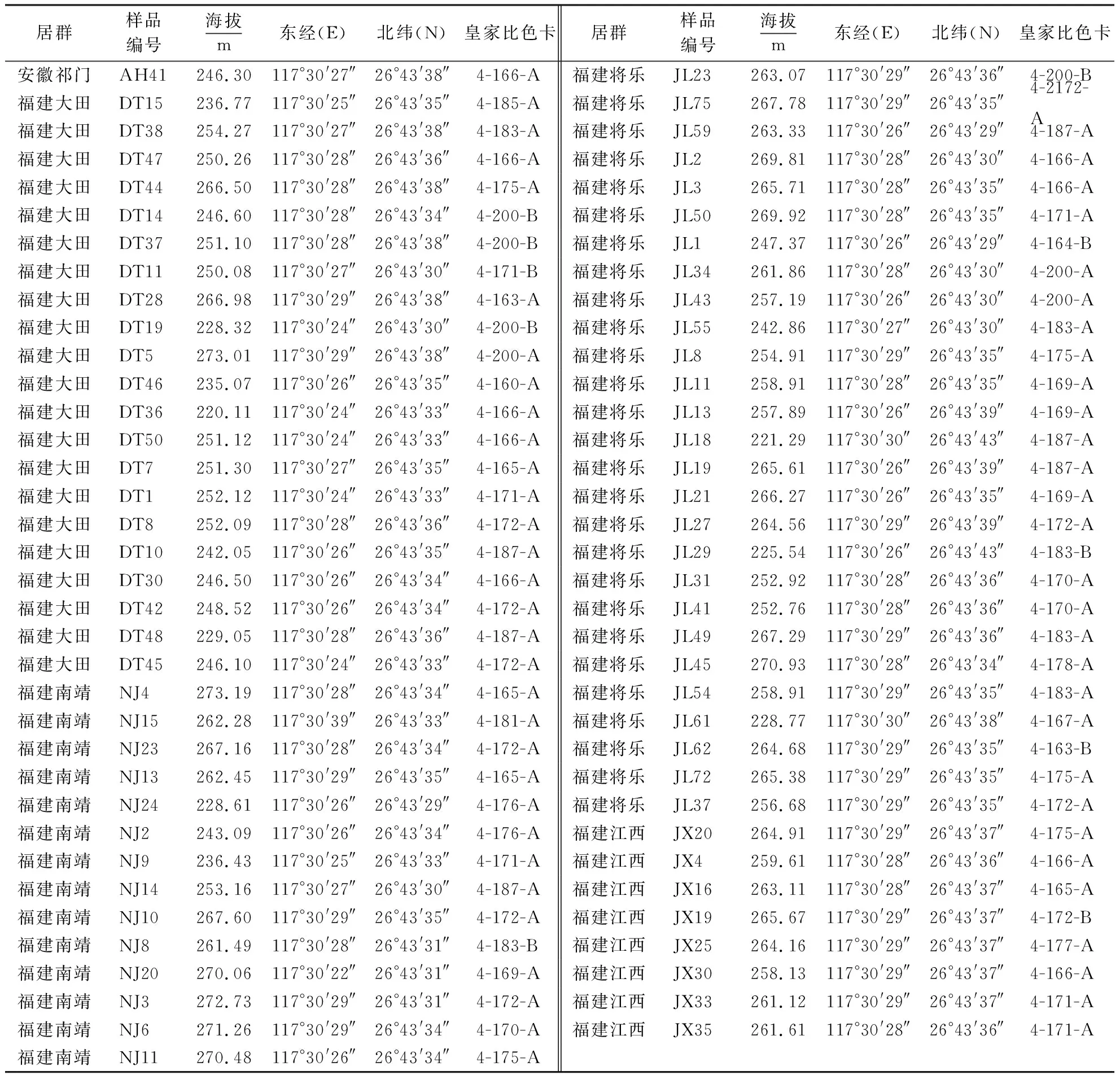
续表1
1.2 主要试剂与仪器
主要试剂包括高效植物基因组DNA提取试剂盒、氯仿、异戊醇、β-巯基乙醇、乙醇、缓冲液A、Goldview核酸染料、Mark DL2000和2×Taq Master Mix、异丙醇.主要仪器设备包括YMY组织研磨仪(浙江托普仪器公司产品)、2-16KL低温冷冻离心机(美国密理博公司产品)、DK-S28恒温水浴锅(上海精宏实验设备公司产品)、VORTEX-KB3旋涡混合器(海门市其林贝尔仪器制造公司产品)、NanoDrop One超微量紫外可见光光度计(上海土森视觉科技有限公司产品)、T100梯度PCR仪(北京楚齐仪表有限公司产品)和Wide Mini-sub cell GT水平电泳槽(济南好来宝医疗器材有限公司产品).
1.3 试验方法
提取枫香基因组DNA采用改良的十六烷基三甲基溴化铵方法[22],所提取的枫香DNA用超微量紫外可见光光度计检测纯度(D260 nm/D280 nm),并用琼脂糖凝胶电泳来检测所提取枫香DNA的质量.将光密度为1.7~2.0的枫香叶片DNA进行ISSR-PCR扩增.用质量分数为1%的琼脂糖凝胶电泳(110 V,30~40 min)检测DNA片段大小和质量.使用1×TAE电泳缓冲液(DNA样品与loading buffer体积比为5∶1)进行点样,计算出浓度后进行稀释,用于后续试验.
ISSR-PCR扩增反应体系[23](20 μL):模板DNA浓度为20 ng,Taq Master Mix用量为10 μL,引物浓度为0.6 μmol·L-1,ddH2O 7.8 μL.PCR反应扩增程序:94 ℃预变性5 min,94 ℃变性30 s.选用本试验室已筛选出的17条引物及退火温度[23](表2),退火30 s,72 ℃延伸90 s,35个循环;最后72 ℃延伸10 min,4 ℃保存.ISSR-PCR的扩增产物利用质量分数为0.8%的琼脂糖凝胶电泳(110 V,30~40 min),成像并保存在Gel DocTM XR凝胶成像系统中.

表2 适合枫香ISSR-PCR扩增的17条ISSR引物的最适退火温度及多态性Table 2 Optimal annealing temperature for ISSR-PCR amplification of L.formosana and polymorphism of 17 ISSR primers
1.4 数据处理
根据扩增条带,记录200~2 000 bp的清晰条带.对同一引物的扩增条带,扩增阳性记为1,扩增阴性记为0,用Excel表格建成0~1数据矩阵.用Popgene 32软件进行亲缘关系分析,用SPSS 19软件对各遗传系数的显著性进行分析.利用NTSYS 2.1软件对数据进行分析,计算样品间的相似系数,并进行非加权组平均法(unweighted pair-group method with arithmetic mean, UPGMA)遗传相似度聚类分析,绘制137份样品和6个居群的遗传聚类图.用iTOL美化遗传聚类图.
2 结果与分析
2.1 DNA的质量检测
本试验所采用的枫香DNA的光密度均为1.7~2.0.电泳检测结果如图1所示,条带明亮、整齐、无污染,说明所提取的样品较好,可用于后续试验.
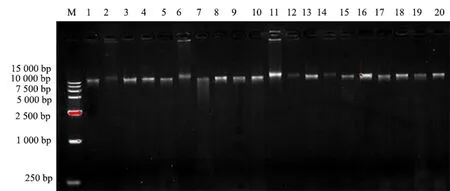
M为15 000 bp的DNA 分子标记量,1~20分别为DT38、DT46、DT36、AH6、AH58、AH63、AH66、AH43、JL59、NJ3、NJ4、NJ8、NJ9、NJ10、NJ24、NJ20、NJ13、JL50、GZ39、GZ51样品编号.图1 部分枫香种质资源DNA提取的电泳效果Fig.1 Electropherogram of DNA extracted from some L.formosana germplasm resources
2.2 同一居群内枫香种质资源的亲缘关系
对6个居群137份枫香材料之间的遗传距离和遗传一致度进行计算,结果表明:137份样品间的遗传一致度为0.350 0~0.922 2,遗传距离为0.081 0~1.049 8.
从表3可知,同一安徽祁门居群内样品的遗传一致度为0.377 8~0.905 6,平均值为0.625 7;遗传距离为0.099 2~0.973 4,平均值为0.489 9.AH3和AH4的遗传一致度最高(0.905 6),相对应的遗传距离最近(0.099 2),说明两者的亲缘关系最近;AH2和AH6的遗传一致度最低(0.377 8),遗传距离最远(0.973 4),说明两者的亲缘关系最远.

表3 6个枫香群体内的遗传距离与遗传一致度Table 3 Genetic distance and genetic identity within 6 L.formosana populations
同一福建大田居群内样品的遗传一致度为0.383 3~0.888 9,平均值为0.603 2;遗传距离为0.117 8~0.958 9,平均值为0.522 8.DT8和DT42的遗传一致度最高(0.888 9),相对应的遗传距离最近(0.117 8),说明两者亲缘关系最近;DT15和DT36的遗传一致度最低(0.383 3),相对应的遗传距离最远(0.958 9),说明两者的亲缘关系最远.
后期时,因经历与北方异族作战,盟中人数骤然减少,风气也更为自由。盟主时常一年轮换一次,也不必特意召开比武大会。而是由现任盟主与选好的下任盟主对打一次,仪式性居多,其结果必定是现任盟主(或真或假的)落败,随后新任盟主上任。
同一福建南靖居群内样品的遗传一致度为0.427 8~0.905 6,平均值为0.6591;遗传距离为0.099 2~0.849 2,平均值为0.426 5.NJ11和NJ16的遗传一致度最高(0.905 6),相对应的遗传距离最近(0.099 2),说明两者亲缘关系最近;NJ13和NJ18的遗传一致度最低(0.427 8),相对应的遗传距离最远(0.849 2),说明两者的亲缘关系最远.
同一福建光泽居群内样品的遗传一致度为0.416 7~0.922 2,平均值为0.628 2;遗传距离为0.081 0~0.875 5,平均值为0.483 2.GZ54和GZ51的遗传一致度最高(0.922 2),相对应的遗传距离最近(0.081 0),说明两者亲缘关系最近;GZ41和GZ4的遗传一致度最低(0.416 7),相对应的遗传距离最远(0.875 5),说明两者亲缘关系最远.
同一福建将乐居群内样品的遗传一致度为0.438 9~0.911 1,平均值为0.642 1;遗传距离为0.093 1~0.823 5,平均值为0.459 8.JL18和JL19的遗传一致度最高(0.911 1),相对应的遗传距离最近(0.093 1),说明两者亲缘关系最近;JL27和JL50的遗传一致度最低(0.438 9),相对应的遗传距离最远(0.823 5),说明两者亲缘关系最远.
同一江西永丰居群内样品的遗传一致度为0.650 0~0.900 0,平均值为0.811 7;遗传距离为0.105 4~0.430 8,平均值为0.211 4.JX4和JX16的遗传一致度最高(0.900 0),相对应的遗传距离最近(0.105 4),说明两者亲缘关系最近;JX20和JX19的遗传一致度最低(0.650 0),相对应的遗传距离最远(0.430 8),说明两者亲缘关系最远.
2.3 不同居群间枫香种质资源的亲缘关系
从表3可知,6个居群中,江西永丰的样品表现为平均遗传一致度最高(0.811 7),相对应的遗传距离最近(0.211 4);福建大田的样品表现为平均遗传距离最远(0.522 8),相对应的平均遗传一致度最低(0.603 2).GZ45和DT36来自不同居群,遗传距离最远,相对应的遗传一致度最低,说明两者的亲缘关系较远.GZ51和GZ54来自相同的居群,遗传距离最近(0.081 0),相对应的遗传一致度最高(0.922 2),说明两者的亲缘关系最近.
从表4、5可知,6个枫香居群的遗传一致度为0.832 8~0.971 1,遗传距离为0.029 4~0.182 9,福建大田和安徽祁门的遗传一致度最高(0.971 1),其对应的遗传距离最近(0.029 4),说明两者亲缘关系最近;福建南靖和江西永丰的遗传一致度最低(0.832 8),其对应的遗传距离最远(0.182 9),说明两者亲缘关系最远.
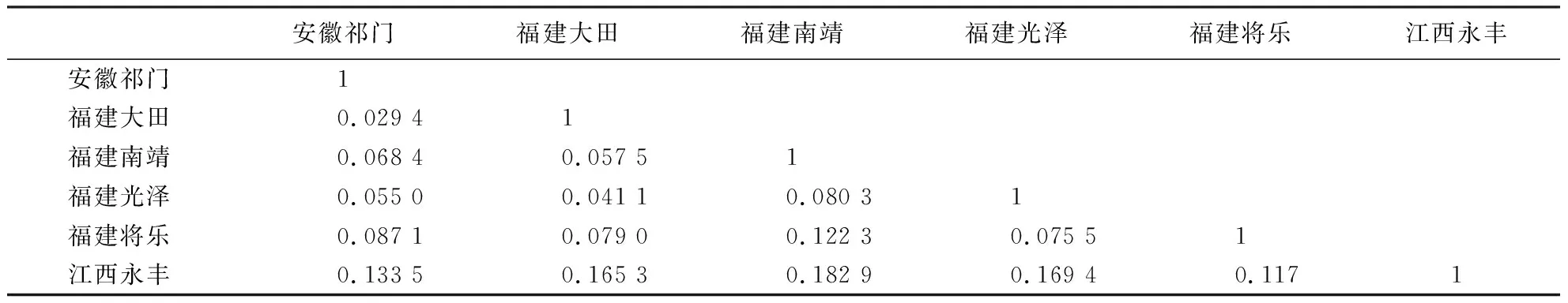
表4 枫香居群间的遗传距离Table 4 Genetic distance among L.formosana populations
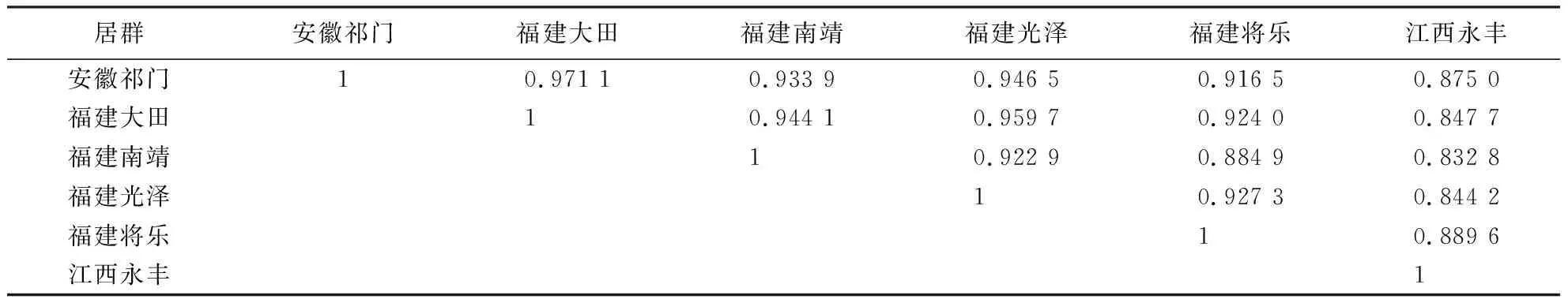
表5 枫香居群间的遗传一致度Table 5 Genetic identity among L.formosana populations
2.4 枫香的遗传多样性
对137份枫香种质资源的总体遗传多样性参数进行分析,结果如表6所示.从表6可知,平均等位基因数为2.000 0,平均有效等位基因数为 1.656 7,平均遗传多样性指数为0.378 8,平均Shannon信息多样性指数为 0.559 0,说明供试的137份枫香样品间的遗传多样性较为丰富.

表6 居群内枫香种质资源的遗传多样性参数分析1)Table 6 Genetic diversity parameters of L.formosana germplasm resource within single population
对6个居群内枫香种质资源进行遗传多样性参数分析,结果如表6所示.从表6可知,6个居群的等位基因数为1.511 1~1.977 8,平均等位基因数为1.638 9;有效等位基因数为1.306 4~1.645 5,平均有效等位基因数为1.506 0;Nei′s基因多样性指数为0.177 7~0.371 9,平均基因多样性指数为0.293 0;Shannon指数为0.265 7~0.508 6,平均Shannon指数为0.435 1;多态位点百分率为51.11%~97.78%,平均多态位点百分率为80.56%;多肽为点数为92~176,平均多态为点数为145.平均等位基因数、有效等位基因数、Nei′s基因多样性指数、Shannon指数、多态位点数和多态位点百分率均表现为福建大田居群最高,江西永丰最低.总体来说,在6个居群中福建大田的遗传多样性高于其他5个居群.
对6个居群间枫香种质资源的遗传多样性参数差异性进行分析,结果表明,等位基因数、有效等位基因数、Nei′s基因多样性指数、Shannon指数、多态位点百分率、多态位点数在江西居群样品与其余5个种源地中均有显著性差异(P<0.05);在福建南靖与福建将乐居群中均无显著性差异(P<0.05).安徽祁门、福建大田、福建光泽种质的等位基因数分别与福建南靖、福建将乐的均有显著性差异(P<0.05);福建大田种质的Nei′s基因多样性指数、Shannon指数与福建南靖、福建将乐的均有显著性差异(P<0.05);福建大田、福建南靖、福建光泽间多态位点百分率、多态位点数均有显著性差异(P<0.05).相对于遗传多样性最高的福建大田居群来说,遗传多样性最低的江西永丰居群的等位基因数少了23.6%,有效等位基因数少了20.61%,基因多样性指数降低了52.22%,Shannon指数降低了51.62%,多态位点数少了47.73%.
M为DL 2 000 DNA 分子标记量,1~24号为部分样品的扩增条带.图2 引物UBC-834与部分样品的扩增结果Fig.2 Amplification results of some L.formosana samples using primer UBC834
2.5 枫香遗传分化
枫香的总遗传分化系数为0.146 3,表明枫香85.7%的遗传分化在居群内,14.63%的遗传分化在居群间.6个枫香居群基因流较大(2.916 5>1),说明不同枫香居群之间的基因交流未受到影响.对6个枫香居群间和居群内的遗传差异分析结果表明,居群内部个体间遗传多样性仅是0.319 0,居群间的遗传多样性为0.373 7.
2.6 UPGMA聚类分析
从图3可看出,当遗传相似系数达到0.92时,将6个居群分为3个大类,第1类包括两个亚群,其中一个亚群包括安徽祁门、福建大田和福建光泽居群,另外一个亚群包括福建南靖居群;第2大类包括福建将乐居群;第3类包括江西永丰居群.
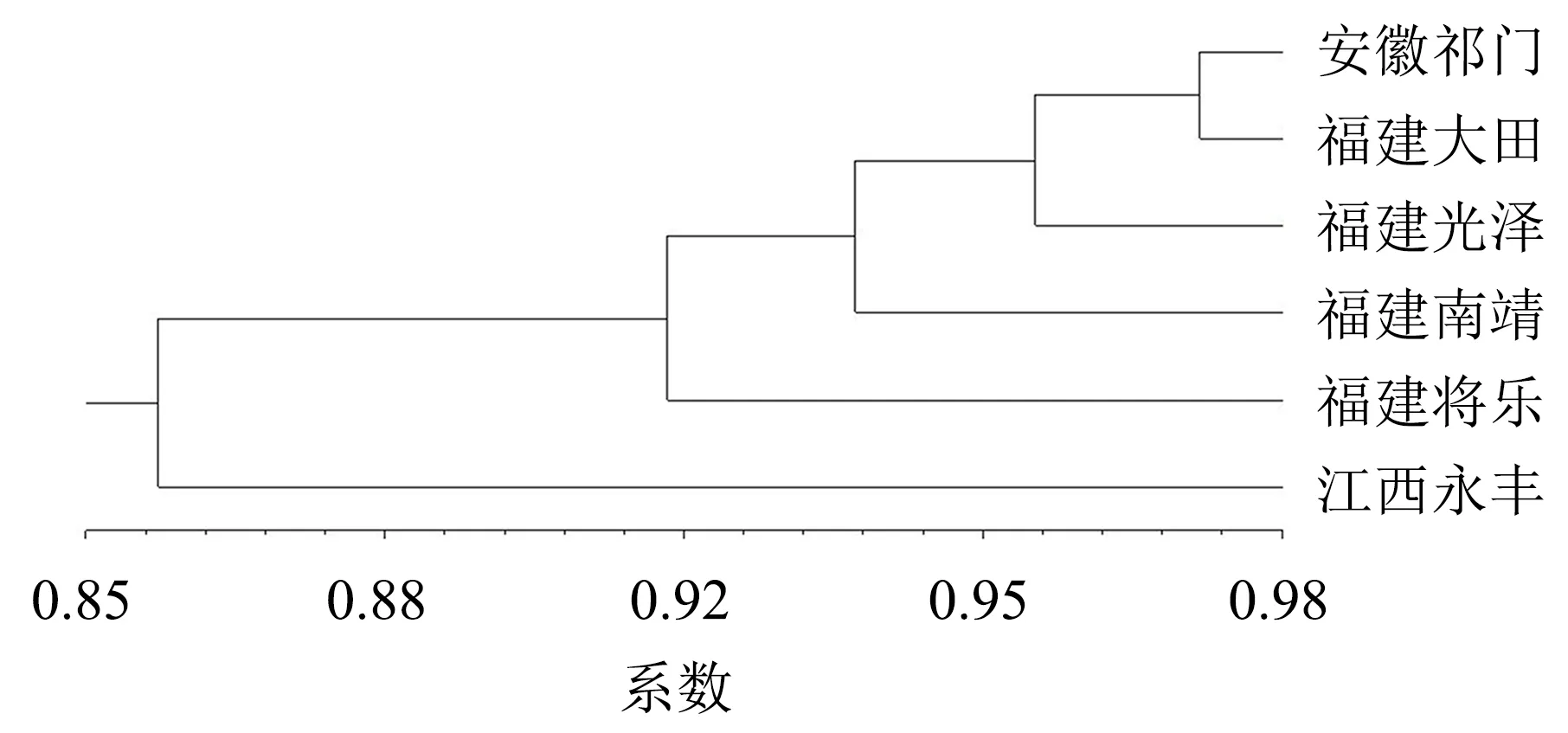
图3 6个群居枫香树的UPGMA聚类图Fig.3 UPGMA cluster map of 6 populations of L.formosana
从图4可看出,当遗传相似系数达到0.62时,将137份枫香材料分为4个大类:第1类包括1份;第2类有两个亚群,一个亚群包括45份,另外一个亚群包括24份;第3类包括两个亚群;第4类包括两个亚群,一个亚群有41份,另外一个亚群有24份.
3 讨论与结论
环境的变化和植物本身内在因素都可能会对遗传多样性造成影响,如外部的气候、温度和内部基因突变、基因流等[23].本试验中平均多态位点百分率为80.56%,表明ISSR是一种枫香亲缘关系鉴定的有效技术手段.137份枫香种质资源遗传多样性高于林骥光[16]和毕泉鑫等[17]对浙江枫香种质资源遗传多样性的研究结果.本试验中,安徽祁门和福建大田居群间的亲缘关系最近,据观察两个居群的枫香叶片叶型相似度较高,说明分子标记可以作为叶片表型分类的辅助手段.枫香居群间的遗传一致度相较于居群内表现为较高且范围较小,遗传距离相较于居群内表现为较近且范围较小,说明枫香居群间有一定的遗传多样性.但是相对于单株来说遗传多样性较低,植物的交配方式会影响植物的群居结构[17],自交的物种遗传多样性主要存在于种群间,异交物种则以种群内为主[25],Sun[26]和李芳芳等[22]对枫香的研究结果表明居群内遗传分化显著高于居群间,而本研究中85.7%的遗传分化在枫香居群内,显著高于居群间的遗传分化,与其研究结果一致.一般认为基因流大于1,可以阻止遗传漂变产生遗传分化[27].本研究中基因流较大(2.916 5>1),说明居群间基因交流丰富,影响了枫香居群间的遗传分化,这与在李芳芳[22]的研究结果一致.

图4 137份枫香树的UPGMA聚类图Fig.4 UPGMA cluster map of 137 L.formosana germplasm resources
在遗传相似系数为0.92时,将6个居群分为3类,第1类为安徽祁门、福建大田、福建光泽和福建南靖居群,第2类为福建将乐居群,第3类为江西永丰居群.第1类中安徽居群和福建的3个居群在地理位置上差异较大,却被聚为一类,说明安徽居群的枫香种质对环境的适应性强,受环境的影响小.第2类福建将乐居群生长状态好,叶片色彩丰富且光泽度.,可能是因为福建将乐居群枫香种质为本地种质,对生长环境适应,也可能是因为枫香自身内部的基因在调控.第3类江西居群单独成类.从实地考察来看,江西居群的枫香种质生长状态差、叶片不易变色且落叶期过早,说明江西居群的枫香种质对环境的适应性弱,受环境的影响大.
猜你喜欢
社会科学战线(2022年7期)2022-08-26
上海工艺美术(2021年3期)2021-09-26
环境与生活(2021年5期)2021-06-24
公关世界(2020年19期)2020-11-13
新课程·下旬(2018年9期)2018-11-14
中国医药导报(2018年14期)2018-08-30
青年时代(2018年29期)2018-01-17
当代贵州(2016年23期)2016-07-21
世界文化(2016年7期)2016-07-04
青少年科技博览(中学版)(2015年10期)2015-01-11
