
糙米和大米非靶向代谢组学分析
2022-10-31 08:56任传英卢淑雯张英蕾管立军黄文功卢卫红
食品科学 2022年20期
任传英,卢淑雯,洪 滨,张英蕾,管立军,3,李 波,3,黄文功,卢卫红
(1.哈尔滨工业大学化工与化学学院,黑龙江 哈尔滨 150001;2.黑龙江省农业科学院食品加工研究所,黑龙江 哈尔滨 150086;3.黑龙江省食品加工重点实验室,黑龙江 哈尔滨 150086;4.黑龙江省农业科学院农产品质量安全研究所,黑龙江 哈尔滨 150086)
水稻是世界上消费量最大的谷物之一,在100多个国家种植,大米是世界一半以上人口的主食,也是中国65%以上人口的口粮。糙米由皮层(6%~7%)、胚(2%~3%)和胚乳(约90%)组成,被称为全谷物。糙米不仅富含基本营养成分,还含有更多的微量元素和生物活性成分,如VB、VE、膳食纤维、酚类化合物、-氨基丁酸、谷维素、低聚糖和植酸。大米是糙米经多道碾磨、抛光、色选,去除全部皮层和胚,仅保留胚乳的产品,因此,糙米在营养上优于大米。
目前,对糙米和大米的研究主要集中在一些已知的营养素和生物活性成分分析,而对于许多未知的营养素和活性成分鲜有报道。代谢组学主要是不同代谢路径产生的底物和产物的小分子代谢物分析(分子质量<1 000 Da),不仅可以全面分析代谢产物含量的变化,还可以阐明不同代谢产物与各处理条件间的变化规律,从而评价不同物质的生物反应及效果。已经在植物代谢组学、微生物代谢组学等领域应用,用于鉴定绿茶的品质和种子萌发过程中代谢物的变化。Pongsuwan等采用高效液相色谱法与质谱法相结合进行不同绿茶的代谢组分析,绿茶中除了检测到表没食子儿茶素外,还检测到未知的表没食子儿茶素没食子酸酯和表儿茶素没食子酸酯,也具有显著差异。本研究利用液相色谱-串联质谱非靶向代谢组学技术研究糙米与大米中的所有代谢产物,采用主成分分析(principal component analysis,PCA)、聚类分析和通路分析等多元统计分析方法,解析糙米和大米间的代谢物种类和数量的变化规律,并探明代谢产物变化参与的关键代谢途径,旨在为糙米的贮藏技术和功能性评价提供新的依据。
1 材料与方法
1.1 材料与试剂
糙米:绥粳18水稻种植于黑龙江省五常市卫国乡,2021年9月末收获脱粒成稻谷,贮存至10月初经砻谷制得糙米,出糙率为81.7%,称取约5 g整粒糙米于10 mL离心管中密封,平行取样3 次,液氮冷冻后贮存于-80 ℃冰箱中,待测。
大米:称取500 g上述糙米采用实验型碾米机碾磨成大米(与糙米制备同一天),整精米率为69.4%。称取5 g整粒大米于10 mL离心管中密封,平行取样3 次,液氮冷冻后贮存于-80 ℃冰箱中,待测。
乙腈、甲酸和甲醇均为色谱纯。
1.2 仪器与设备
Yamamoto Vp-32实验型碾米机 日本佐竹公司;2777C UPLC system液相色谱仪、Xevo G2-XS QTOF质谱仪 美国沃特世公司。
1.3 方法
1.3.1 代谢物提取
将-80 ℃保存的测试样品取出,立即从每管中各选取1 粒米(单粒质量21~23 mg),具体提取过程按照每粒米的实际质量(精确到0.000 1 g)折算成下述25 mg体系等比例添加试剂进行代谢物的提取,具体操作如下:25 mg测试样品倒入1.5 mL EP管中,加入800 µL预冷沉淀剂(甲醇-乙腈-纯水,2∶2∶1,/),放入小钢珠2 颗,转入研磨机中研磨(60 Hz,4 min),小钢珠取出后再将样本冰浴超声(80 Hz,10 min),放入-20 ℃冰箱,静置120 min后离心(4 ℃,25 000×,15 min),取出上清液600 µL,将上清液转入冻干机中冻干,加入10%甲醇溶液600 µL复溶,80 Hz冰浴超声10 min,然后25 000×、4 ℃离心15 min,取出上清液,6 个样本各取出50 µL,然后混合成一个样本(质控样本,quality control,QC)。
1.3.2 液相色谱条件
ACQUITY UPLC HSS T3色谱柱(100 mm×2.1 mm,1.8 μm);柱温50 ℃,流速0.4 mL/min,流动相A为0.1%甲酸溶液,流动相B为0.1%甲酸-甲醇溶液。梯度洗脱程序:0~2 min,100% A,0% B;2~11 min,100%~0% A,0%~100% B;11~13 min,0% A,100% B;13~15 min,0%~100% A,100%~0% B。每个测试样本的上样体积均为5 µL。
1.3.3 串联质谱条件
利用Xevo G2-XS QTOF高分辨串联质谱对从液相色谱柱洗脱下来的小分子,分别进行正、负离子2 种模式采集。正离子采集模式下,锥孔电压和毛细管电压分别为40.0 V和3.0 kV。负离子采集模式下,锥孔电压和毛细管电压分别为40.0 V和2.0 kV。采用全信息串联质谱模式进行质心数据采集,一级扫描是在50~1 200 Da范围内扫描0.2 s,然后使用20~40 eV的能量对所有母离子进行碎裂,再进行二级扫描0.2 s采集所有的碎片信息。在采集数据的过程中,需要对LE信号每间隔3 s进行实时质量校正一次,并且,在测试样本前采集一次QC样本,用于评价样本在整个采集过程中仪器的稳定性状态。
1.3.4 峰提取及鉴定
主要通过软件Progenesis QI(version 2.2)实现,包括峰对齐、峰提取、归一化、谱图解析和化合物鉴定等步骤。峰提取和鉴定的主要参数设置如表1所示。

表1 Progenesis QI主要参数Table 1 Major parameters of Progenesis QI
1.3.5 差异代谢物的鉴定
采用自主研发的代谢组学metaX软件包和Progenesis QI(version 2.2)软件对质谱分离数据进行统计学分析,其中代谢物的鉴定来源于KEGG、HMDB和LipidMaps数据库。差异表达代谢物的筛选采用多变量偏最小二乘判别分析(partial least square-discriminant analysis,PLS-DA)模型前2 个PC的变量投影重要性(variable importance in projection,VIP)值,再结合值和单变量分析差异倍数筛选。筛选条件为:1)VIP≥1;2)<0.053);3)差异倍数≤0.833 3或差异倍数≥1.2,3 个条件取交集,最后得到共有的代谢物即差异代谢物。最后结果以火山图形式呈现VIP值、值和差异倍数三项指标。
1.3.6 代谢物的分类和功能注释
HMDB(www.hmdb.ca)是一个可联网的代谢组学数据库,包含有关人类代谢物、生理浓度、疾病关联、化学反应、代谢途径、参考光谱及其生物学作用的综合信息,具有代谢途径和图谱搜索功能。KEGG是从分子水平信息了解生物系统(如细胞,生物和生态系统)的高层次功能的数据库,尤其是通过基因组测序和其他高通量实验技术而产生的大规模的数据库资源。其中KEGG Pathway数据库是对生命活动中的分子相互作用和化学物质反应的认识,构建了复杂的调控网络,通过强大的通路图形式展示关联的代谢途径及各途径之间的相关性,并进行功能注释,明确特定代谢物参与的主要生化代谢途径和信号转导途径。
1.4 数据处理
首先对质谱分离鉴定的原始数据提取峰列表信息,进行数据预处理和校正,然后按照1.3.5节进行数据统计分析,找出样品间差异表达的代谢物,最后,参考KEGG、HMDB和LipidMaps数据库对代谢物及差异代谢物进行鉴定,并通过KEGG Pathway数据库进行Pathway分析,确定差异代谢物参与的主要代谢途径。
统计分析:包括单变量分析和多变量分析。其中单变量分析包括非参数检验或参数检验、倍数分析。多变量分析包括PCA、聚类分析和PLS-DA。
2 结果与分析
2.1 质控分析
代谢组学研究中,QC分析是评价质谱技术可靠性和高质量的重要方法。在样本检测前首先进行QC样本分析测试,用以平衡、稳定“色谱-质谱”系统,连续扫描得到的图谱为总离子流图(图1)。QC样本为同一个样本,重叠程度越高、峰分离度越高,可以判定本研究的质谱分析仪器状态良好且稳定。
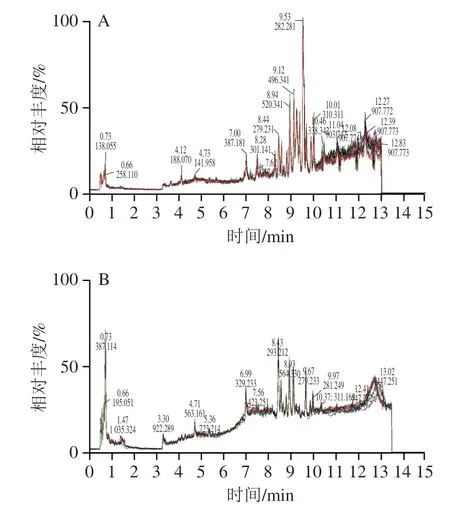
图1 QC样本总离子流重叠图Fig.1 Overlapping total ion current (TIC) chromatograms of quality control (QC) samples
2.2 差异代谢物数量和类别鉴定结果
差异代谢物的定性和相对定量基于数据库KEGG、HMDB、LipidMaps分析得出。从数量上看,糙米与大米相比,正离子模式下,分析出差异代谢物460 个,其中上调300 个、下调160 个。负离子模式下分析出差异代谢物579 个,其中上调383 个、下调196 个(表2)。2 种模式下,上调差异代谢物均多于下调差异代谢物。从类别上看,糙米与大米相比,正离子模式下,差异代谢物中上调的主要是黄酮类、脂肪酸类、羧酸类、有机氧化物、苯类;差异代谢物中下调的主要是黄酮类、脂肪酸类、羧酸类、甘油磷酸和异二戊烯类,黄酮类、脂肪酸类和羧酸类为上调和下调共有类别。负离子模式下,差异代谢物中上调的主要是黄酮类、有机氧化物、脂肪酸类、羧酸类和肉桂酸类,差异代谢物中下调的主要是黄酮类、有机氧化物、脂肪酸类、羧酸类和甘油磷脂(图2)。2 种离子模式下分析的差异代谢物数量极多,其中多数仅有变化趋势,但是差异不显著,见图3。总体上看,糙米与大米相比,差异代谢物中黄酮类最多。黄酮类物质具有抗病毒、抗自由基、抗氧化、降低脂质过氧化反应、抗炎、抗衰老、防癌和预防心血管疾病等作用。Kris-Etherton等研究发现酚类化合物包括亚类黄酮类化合物,广泛存在于在谷物、豆类和坚果中。Yang Zhigang等已经证明,黄酮类物质如麦黄酮、7--芸香苷和7----吡喃葡萄糖苷主要存在于子叶和麸皮中,但不存在于碾磨的谷物中。多酚类物质也是糙米中重要的植物类营养素,糙米中游离酚和结合酚含量均显著高于白米,游离酚平均含量为101.64 mg/100 g,粳型糙米平均含量略高于籼型糙米。糙米在胃部和小肠消化阶段还会显著促进酚类物质释放,消化液中的单体酚酸主要为阿魏酸和原儿茶酸。本研究发现,鉴定到的黄酮类物质中部分由脂肪酸降解得到,如表儿茶素-吡喃葡萄糖苷-苯甲酸酯、芥子油基酯、8-丙炔二吡喃酮、儿茶素-4-醇和甘丙烯醇。这是由于糙米中脂质含量虽然只有1%左右,但易分解,在糙米储藏过程中,脂质在酯酶的作用下分解产生游离脂肪酸,脂肪酸不稳定,可进一步降解产生酚类、酮类物质。

表2 糙米与大米差异代谢物鉴定Table 2 Differential metabolites between brown rice and while rice

图2 主要差异代谢物的类别Fig.2 Classification of major differential metabolites
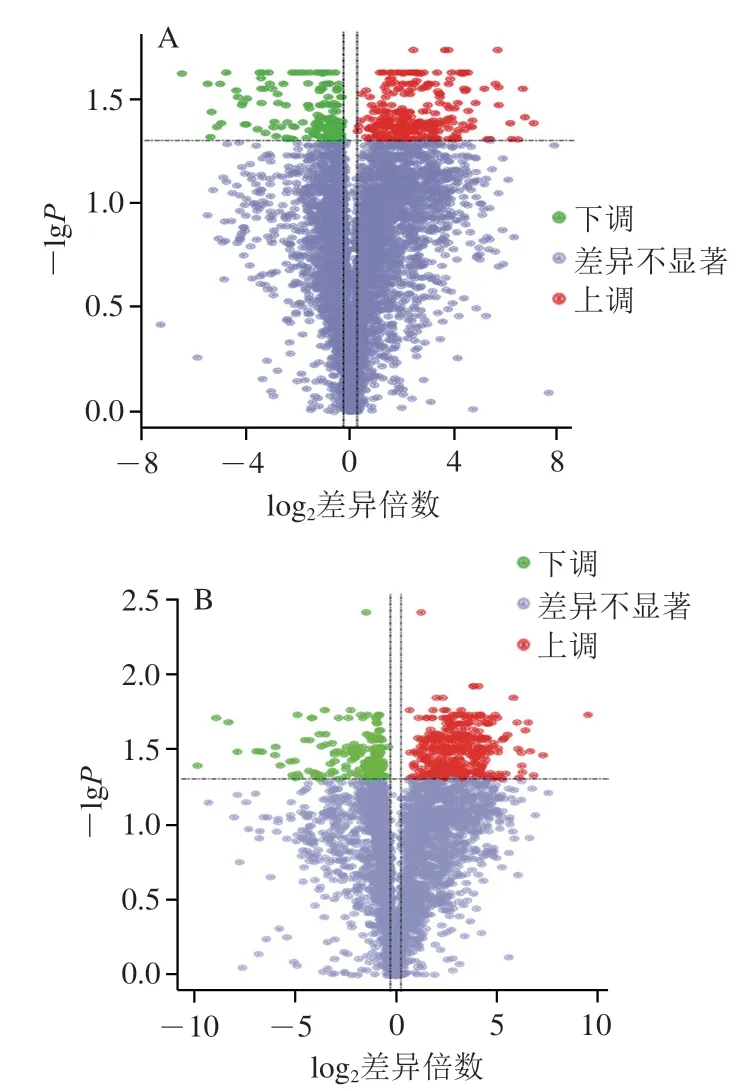
图3 差异代谢物火山图Fig.3 Volcano map of differential metabolites
2.3 差异代谢物的PCA
PCA可反映样本中代谢物丰度情况,样本之间位置越近则越相似,越远则反之。对糙米与大米差异代谢物进行PCA,见图4。2 组样品的置信区间均在95%以内,糙米与大米样品的PC明显不同,分别位于置信区间的左右两侧。从分布模式看,负离子模式下糙米样本之间的距离更近,说明3 个平行样品中的差异代谢物鉴定情况十分接近。正离子模式下,每组3 个平行样品中的差异代谢物均有一定的离散性,说明组内样品间也存在一定的个体差异。这可能是由于测试样品组内的3 个米粒成熟度和粒位不同,导致鉴定到的差异代谢物有所差异。研究表明,成熟度和粒位对大麦品质、营养性和耐储性均有影响。贵州主栽水稻品种不同成熟期大米品质变化,发现在乳熟、蜡熟、成熟、完熟4 个时期中,各质量指标在中后期较高,而在后期有所下降。
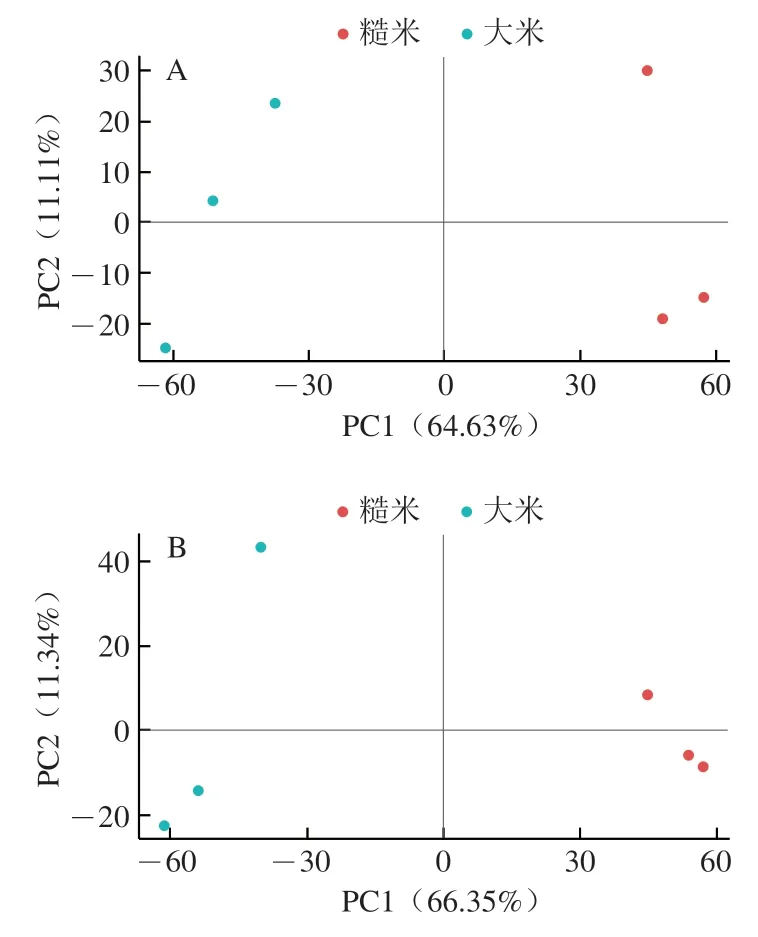
图4 PCA散点图Fig.4 PCA score scatter plots
2.4 差异代谢物的分类和功能注释
糙米与大米全部差异代谢物(正离子模式460 种和负离子模式579 种)聚类分析热图见图5。不同颜色表示不同含量,红色表示含量上调,绿色表示含量下调,从绿色到红色表示该物质表达量由低到高。通过对鉴定的代谢物进行聚类分析后,已鉴定的代谢物主要分为3 类,包括生物活性物质、脂质和植物化学物,其中具有生物活性的化合物居多,正离子模式下生物活性的化合物主要是氨基酸/肽、苯类和有机酸类,如-精氨酸、瓜氨酸、3-甲基组氨酸、-乙酰高丝氨酸、5-羟基--色氨酸、三甲基--组氨酸、5-羟基--色氨酸、谷氨酰--半胱氨酸、4-氟苯甲酸、苯丙酮酸、吲哚、赛洛辛、4-羟基苯甲醛和-乙酰神经氨酸9-磷酸等。相关研究发现,糙米与大米中的蛋白质含量虽然差异不显著,但是氨基酸总量和必需氨基酸总量差异显著,其中以组氨酸、苯丙氨酸和缬氨酸为主。脂质类主要是脂肪酸、脂肪酰、脂肪醇、脂肪酯、脂肪酰胺和脂肪醛。植物化学物主要是黄酮类、生物碱,如鼠李糖苷、染料木黄酮、甘黄酮A、2-脱羧基甜菜碱、天竺葵素、4,8-二羟基-2-呋喃、头孢拉定A和苯并呋喃-1,3-二醇。类黄酮是植物酚类化合物的重要组成,Dong Xuekui等采用广靶代谢组学方法对90多种黄酮类化合物同时鉴定和定量,比较水稻不同发育阶段不同组织中的类黄酮含量,研究表明,黄酮单-C-糖苷、丙二酸化黄酮--己糖苷和一些黄酮--糖苷为主要差异代谢物。Wang Changyuan等采用气相色谱-质谱法进行不同贮藏期糙米代谢组学分析,发现糙米贮藏后代谢产物的数量和类型发生了显著变化,由于采用的分析方法和鉴定数据库不同,仅筛选了34 种差异代谢物,储藏后糙米的碳水化合物、氨基酸和脂肪酸含量降低,而糖醇、胺和醛类含量增加。

图5 差异代谢物聚类分析图Fig.5 Cluster analysis of differential metabolites
如图6所示,正离子模式下,主要代谢是氨基酸代谢、其他次生代谢物的生物合成、辅因子和维生素代谢;负离子模式下,主要代谢是辅因子和维生素代谢、其他次生代谢物的生物合成、碳水化合物代谢、核苷酸代谢和氨基酸代谢(没有分类信息的鉴定结果不参与统计)。对于差异代谢物显著富集的Pathway绘制气泡图(图7),参与相应代谢途径的代谢物鉴定结果见表3、4。正离子模式下,代谢物参与的代谢途径差异显著的主要是苯丙氨酸、酪氨酸、色氨酸的生物合成和组氨酸代谢,其中参与前者的代谢物是苯基丙酮酸和吲哚,参与后者的代谢物是3-甲基组氨酸和三甲基--组氨酸;负离子模式下,代谢物参与的代谢途径差异显著的主要是甜菜碱生物合成、嘌呤代谢、碳代谢、玉米素生物合成和C-支链二元酸代谢,其中参与甜菜碱生物合成的代谢物是2-脱羧基甜菜碱、天竺葵II和七叶树素II,导致糙米具有一定的涩味,参与嘌呤代谢的代谢物是P1,P4-双(5′-黄糖基)四磷酸,参与碳代谢的代谢物是--葡萄糖-6-磷酸、中康酸和5,10-甲基四氢叶酸,参与玉米素生物合成的代谢物是顺式玉米素-7--糖苷和腺嘌呤,参与C-支链二元酸代谢的代谢物是中康酸和反式乌头酸。Zhao Mingchao等采用液相色谱-质谱/质谱法代谢组学分析到黑米中257 种次级差异代谢物,大多数差异代谢物参与的代谢途径主要是酪氨酸代谢、核黄素代谢、类黄酮生物合成、花青素生物合成和苯丙烷类化合物生物合成。
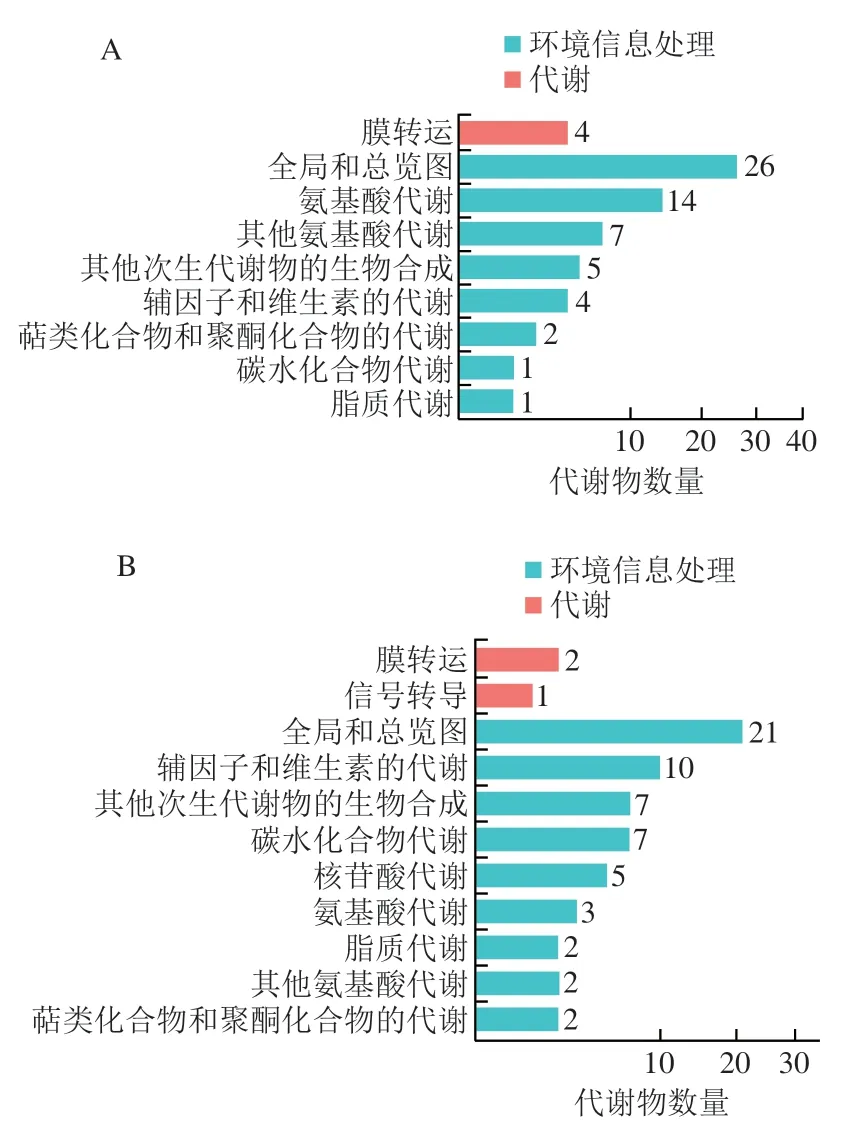
图6 KEGG功能注释条形图Fig.6 Bar graphs of KEGG function annotation

图7 代谢途径富集分析气泡图Fig.7 Bubble diagrams of metabolic pathway enrichment analysis

表3 差异代谢物鉴定结果(正离子)Table 3 Results of identification of differential metabolites in the positive ion mode

表4 差异代谢物鉴定结果(负离子)Table 4 Results of identification of differential metabolites in the negative ion mode
3 讨论
近年来,糙米因其具有较高的营养品质而越来越受到人们的关注,全世界都推荐食用糙米。糙米和精米之间的营养素差异已经被许多研究所确定。Vichapong等发现,黑米、红米、紫米和糙米中酚类化合物、总黄酮和抗氧化活性均高于大米,鉴定的12 种酚类化合物均为游离酚类化合物,其中阿魏酸含量最高。本研究基于液相色谱-串联质谱非靶向代谢组学分析糙米与大米中的差异代谢物,PCA表明,在正离子和负离子模型下,大米和糙米是分离的。PCA正确反映了相应样品中代谢物的积累模式和特异性,该结果表明大米和糙米之间存在代谢组学差异。在这些差异代谢物中,类黄酮最多。负、正离子模式分别为89 个和31 个,除了糖苷类,还鉴定到脂肪酸分解物(如儿茶素-吡喃葡萄糖苷-苯甲酸酯、芥子油基酯、8-丙炔二吡喃酮、儿茶素-4-醇和甘丙烯醇)、与糙米颜色和口感相关的代谢物(胡椒碱、甘草苷、米香苷和姜黄素)。黄酮类化合物是众所周知的药用植物活性成分和生物活性食品成分,可以改善内皮功能并可能降低血压,体外研究表明,多种黄酮类化合物可引起大鼠离体动脉的血管舒张,内皮功能的改善可有助于降低血压。
植物使用脂肪酸合成酰基脂质,用于许多不同的细胞、生理和防御作用,包括合成必需的膜、储存或表面脂质,以及用于信号或防御的各种脂肪酸衍生代谢物的产生,脂肪酸通过与辅酶A或酰基载体蛋白的硫酯键被激活以进行代谢处理。特定的不饱和脂肪对正常寿命、生理功能和神经发育有重要作用,相应的下游代谢物是重要的脂质信号分子。脂肪酸在人体细胞膜结构功能、物质储存和细胞信号传导中发挥重要作用,食用不饱和脂肪酸对人体健康有保护作用,一些必需脂肪酸(亚油酸和-亚麻酸)只能从饮食中摄取。因此,类黄酮和脂肪酸的代谢变化可能有助于糙米的营养价值评价。此外,使用KEGG数据库对差异离子进行注释,正离子模式下代谢途径为2 条(苯丙氨酸、酪氨酸和色氨酸的生物合成、组氨酸代谢),参与代谢物为4 种;负离子模式下代谢途径为5 条(甜菜碱生物合成、C-支链二元酸代谢、嘌呤代谢、玉米素生物合成和碳代谢),参与代谢物为10 种。结果发现,全谷物糙米与碾磨后的大米相比,差异代谢物上调为主,而且通过影响氨基酸代谢、碳代谢、嘌呤代谢、玉米素代谢和甜菜碱生物合成等代谢途径,调节米糠和胚中多种氨基酸、多酚、脂肪酸类物质含量,提升稻米的营养品质。
猜你喜欢
中国药房(2022年10期)2022-05-30
三农资讯半月报(2020年19期)2020-10-27
分析化学(2019年3期)2019-03-30
分析化学(2018年4期)2018-11-02
分析化学(2017年12期)2017-12-25
小学教学参考(语文)(2017年11期)2017-11-30
成长·读写月刊(2017年7期)2017-07-13
中国民族民间医药·上半月(2017年5期)2017-06-12
中国中药杂志(2016年21期)2017-02-16
热带农业科学(2015年4期)2015-06-18
