
不同品种鸭胚胎期脂肪酸合成差异性研究
2022-10-29 07:16李卿青周彦四川省达州市达川区畜牧发展服务中心
中国畜牧业 2022年19期
文│李卿青 周彦(四川省达州市达川区畜牧发展服务中心)
甘伟(四川省达州市农业科学研究院)
文│刘芳 马东* 冯波 高田田 刘旺(陕西省榆林市羊产业发展中心)
胚胎期各物种的脂肪合成部位不同,哺乳动物主要在脂肪组织合成,禽类则在肝中合成。禽类胚胎期脂肪组织的发育取决于血浆甘油三酯(TG)的活性,TG主要来源于肝,通过脂蛋白将合成的脂肪酸输出到肝外组织,为机体提供能量。同时,肝的脂肪合成还可直接促进脂肪组织原位脂肪沉积和长期能量储存。
胚胎期被认为是动物生长发育最重要、最敏感的时期之一,它直接决定了动物未来的生长发育。研究表明,在卵裂早期增加葡萄糖和氧气消耗可以改变卵裂晚期的代谢活性。
在胚胎发育过程中,人们对不同品种鸡的肝脂肪生成情况进行了研究,但不同鸭品种间是否存在差异还未有报道。本研究选择北京鸭、建昌鸭、黑鹜鸭3个鸭品种的20日龄(E20)和28日龄(E28)胚胎为样本,通过检测血浆中总胆固醇(TC)、TG、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)含量和肝中乙酰辅酶A羧化酶(ACC)、脂肪酸延长酶(ELOVL)、脂肪酸合成酶(FAS)表达量来探索不同品种鸭胚胎期脂肪酸合成的差异性。
一、材料与方法
1.试验动物与试验设计。试验选取北京鸭、建昌鸭和黑鹜鸭3个品种,每个品种选取20个鸭受精蛋进行孵化,在孵化的第20、28天,每个品种分别随机选出10个试验蛋采集胚胎肝组织样本,-80℃下冷冻保存。从试验蛋大侧的毛细血管网络中采集血液,血清经4℃3000转/分钟离心10分钟,-20℃保存。
2.酶活指标检测。TC测定试剂盒、TG测定试剂盒、ACC测定试剂盒、ELOVL测定试剂盒、FAS测定试剂盒均来自中国四川制造生物技术公司,HDL-ELISA试剂盒来自中国上海邦毅,LDL-ELISA试剂盒来自中国迈瑞生物。试验操作按照各试剂盒说明书进行。
3.RNA分离及cDNA合成。从-80℃冰箱中取出冻存组织,在液氮中研磨成粉,按照RNA提取试剂盒说明书提取总RNA。将提好的RNA于1.5%琼脂糖凝胶中检测其完整性,并在紫外分光光度计260/280纳米波长处检测纯度和浓度。取部分检测合格的RNA,反转录第一链cDNA(具体步骤参照cDNA第一链合成试剂盒说明书),将产物稀释后保存在-20℃冰箱中备用。
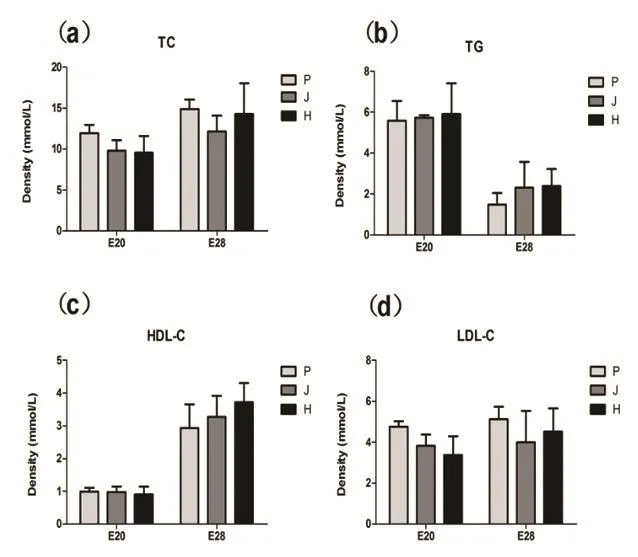
◎图1 血脂因子TC、TG、HDL-C、LDL-C酶活性
4.基因表达测定。实时荧光定量在BIO-RAD CFX system荧光定量PCR仪上进行,反应体系:2×SYBR premix Ex TaqTM 6.25微升,cDNA 1微升,10微米上下游引物各0.25微升,ddH2O 4.75微升。反应程序为:95℃预变性3分钟;95℃变性10秒,64℃ 退火30秒,95℃延伸10秒,40个循环,每个试验重复3次,并以不加模版的cDNA为阴性对照。基因的特异性引物如表1所示,β-actin和GAPDH-rRNA作为内参基因。所有的反应一式三份,数据代表3个独立试验的平均值。目的基因mRNA相对表达量采用2-ΔΔCT方法,根据基因PCR反应的Ct值进行计算,计算结果使用内参基因进行标准化校正。

表1 定量PCR引物
5.统计分析。试验数据使用SPSS 20进行单因素方差分析,采用Duncan’s法进行多重比较,结果以“平均值±标准差”表示,其中P<0.05表示组间差异显著,P<0.01表示组间差异极显著。
二、试验结果
1.3个鸭品种E20和E28期间血脂因子TC、TG、HDL-C、LDL-C酶活性。血脂水平的高低反映了鸭的脂肪酸合成能力,由图1可知,E20和E28两个阶段,不同品种间血脂因子表达量无显著差异。E20时期,3个品种的各血脂因子表达量由多到少依次是TC、TG、LDL-C、HDL-C,E28时期,3个品种的各血脂因子表达量由多到少依次是TC、LDL-C、TG、HDL-C。
2.3个鸭品种E20和E28期间肝组织中ACC、ELOVL、FAS mRNA表达量。由图2可知,E20时期北京鸭3个基因均极显著高于建昌鸭和黑鹜鸭(P<0.01),同时,建昌鸭极显著高于黑鹜鸭(P<0.01)。E28时期,北京鸭的ACC、ELOVL、FAS表达量均显著低于建昌鸭(P<0.05),黑鹜鸭ELOVL表达量显著高于北京鸭(P<0.05),建昌鸭FAS表达明显高于黑鹜鸭(P<0.01)。
3.3个鸭品种E20和E28期间肝组织中ACC、ELOVL、FAS酶活性。由图3可知,E20时期,ACC在3个品种间无显著差异,ELOVL的表达量建昌鸭和黑鹜鸭显著高于北京鸭(P<0.05),FAS仅北京鸭显著高于建昌鸭(P<0.05)。E28时期,ELOVL的表达量在3个品种间无显著差异,建昌鸭的ACC显著高于黑鹜鸭(P<0.05),北京鸭的FAS显著高于建昌鸭(P<0.05)。
三、讨论
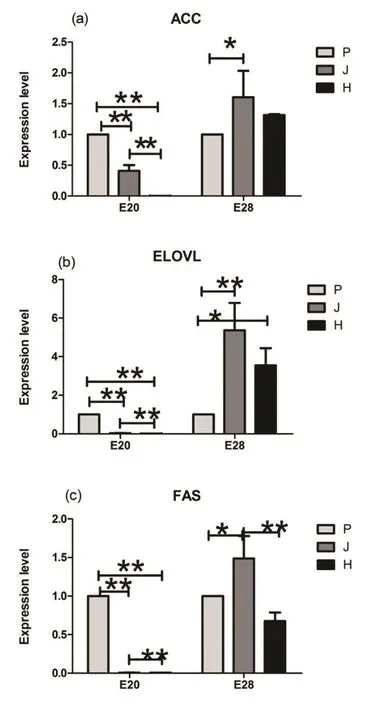
◎图2 肝脂肪合成关联基因mRNA表达水平
肝产生的脂质包括甘油三酯、胆固醇和磷脂,甘油三酯通过血浆极低密度脂蛋白转运到肝外脂肪组织,胆固醇通过与HDL-C或LDL-C结合,在血清中循环。ACC、ELOVL和FAS是肝脂肪合成过程中的关键酶,ACC为限速酶,可催化乙酰辅酶A的羧基化,ELOVL将短链脂肪酸转化为长链脂肪酸以储存能量,FAS催化乙酰辅酶A和丙二酰辅酶A的缩合,通过复杂的七步反应合成长链脂肪酸。因此,我们可以通过上述参数来探讨肝中脂肪合成量。
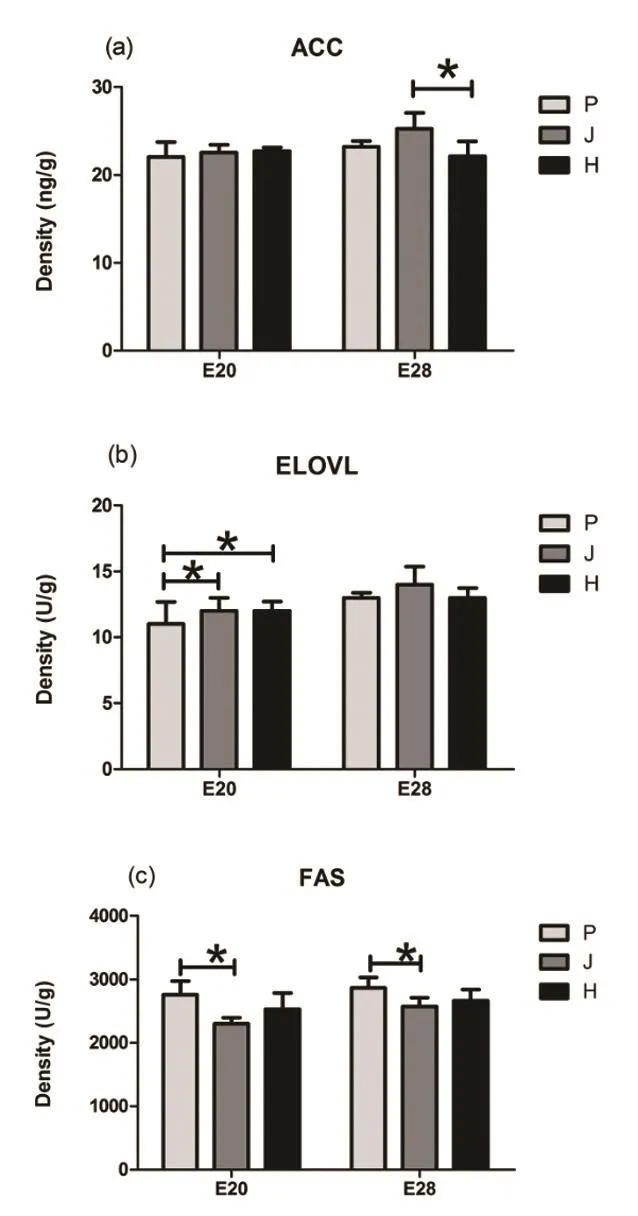
◎图3 肝脂肪合成关联基因酶活性
本研究表明,TC含量高于TG,提示胚胎期胆固醇是主要脂肪酸。3种鸭的TC、TG、HDL-C、LDL-C差异无显著性(P>0.05)。但在其他研究中,品种间TC含量存在差异,如7个自交系鸡蛋黄胆固醇含量差异显著。这些研究大多在卵模型中进行,至今还没有直接研究胚胎期不同品种血浆胆固醇含量。E28的TG含量明显低于E20,可能是由于蛋壳破裂当天TG急剧消耗所致,HDL-C的情况相反,这一趋势可能提示E28阶段HDL转运到肝外的胆固醇多转化为胆汁。E20阶段北京鸭ACC、ELOVL、FAS基因表达明显高于建昌鸭和黑鹜鸭,这些结果与脂肪型鸡ACC基因表达高于胚胎中的瘦肉型鸡结果一致。
以上结果表明,3个鸭品种在胚胎期肝新生脂肪生成(DNL)过程中没有显著差异。以往试验数据表明,北京鸭皮下脂肪和腹部脂肪含量高于建昌鸭和黑鹜鸭。其原因可能是,一方面,DNL的容量受年龄影响。丁芳研究表明,北京鸭肝中,脂质基因表达在第4周或第5周达到高峰,然后随机体生长而下降,这与其他报告的结果一致,即鸟类脂肪细胞的DNL容量主要取决于年龄。另一方面,脂肪酸/甘油三酯的来源有4种:新脂肪发生、细胞质甘油三酯储存、直接吸收肝脂蛋白残基甘油三酯衍生脂肪酸和脂肪组织释放的血浆非酯化脂肪酸,因此,鸭胚胎期脂肪发生的主要方式还有待进一步研究。
四、结论
综上所述,3种鸭胚胎期DNL过程存在显著差异,但血清中最终脂肪酸含量无明显差异。这可能是DNL不是脂肪酸合成的惟一决定因素。本研究首次开展了不同品种鸭胚胎期DNL的研究,从一个角度揭示了不同品种鸭品种间的脂肪形成状况,为今后肝脂肪合成提供了一定的理论参考。
猜你喜欢
传染病信息(2022年3期)2022-07-15
法医学杂志(2022年1期)2022-06-21
健康博览(2021年11期)2021-12-10
作文·小学中高年级(2021年2期)2021-07-01
健康博览(2020年11期)2020-11-23
保健与生活(2020年6期)2020-03-20
家庭用药(2018年9期)2018-11-26
中国卫生产业(2018年12期)2018-05-14
农家参谋(2017年1期)2017-08-13
大众科学(2017年2期)2017-03-02
