
杉木应压木形成中的显微特征及主要代谢成分变化
2022-10-29 01:33杜明秋钟珊丽林二培黄华宏
核农学报 2022年11期
杜明秋 钟珊丽 林二培 黄华宏
(浙江农林大学亚热带森林培育国家重点实验室,浙江 杭州 311300)
木材即次生木质部,是一种被广泛使用的天然材料,与人类的经济社会生活密切相关。树木管胞细胞壁的发育是针叶树木材形成的关键,次生壁作为管胞细胞壁中最厚的一层,对木材的材性影响较大。次生壁是由木质素(18%~35%)和碳水化合物(65%~75%)及少量其他物质交织而成的天然高分子材料,其中碳水化合物包括纤维素(40%~50%)与半纤维素(15%~35%)[1-3]。纤维素是由葡萄糖组成的大分子多糖,其合成与葡萄糖聚合有关[4-5]。半纤维素因物种和细胞类型而异,主要分为三类,即聚木糖类、聚葡萄甘露糖类和聚半乳糖葡萄甘露糖类。针叶材的半纤维素以聚半乳糖葡萄甘露糖类为主[6]。与纤维素、半纤维素(本质上是多糖)不同,木质素是由3种醇单体(对香豆醇、松柏醇、芥子醇)通过碳-碳键和醚键连接形成的一种复杂酚类聚合物。这3种醇单体分别对应三类木质素单体,即对羟苯基木质素(para-hydroxy-phenyl lignin,H-木质素)、紫丁香基木质素(syringyl lignin,S-木质素)和愈创木基木质素(guaiacyl lignin,G-木质素)。其中,愈创木基木质素(G-木质素)是裸子植物中主要的木质素单体,这种芳香族聚合物是重要的次生壁成分[7]。纤维素、半纤维素和木质素等主要成分在次生壁中的类型、结构及含量是决定木材力学和化学特性的主要因素。
应力木通常指在外力作用下形成的弯曲树干,或树枝试图恢复到自然生长状态,而形成具特殊结构和化学组成的木材组织,是研究木材形成的良好模型[8]。在裸子植物中,常出现在倾斜树干或树枝的下部,被称为应压木,与之相对的部位称为对应木[9-10]。在成熟的应压木木质部中,管胞呈圆形或椭圆形,管胞间隙明显,细胞壁出现螺纹裂隙;且应压木次生壁中只有S1和S2层,缺失S3层,同时S2外围多了一层高度木质化区域,称为S2L[6,10-11]。S2L层的木质素含量高,并且异常出现(1,4)-β-半乳聚糖,是裸子植物应压木的典型特征[6]。除了解剖特征的差异,前人通过对比杉木、火炬松与红松应压木和对应木极性代谢物图谱,发现参与纤维素与木质素合成的代谢产物在应压木中产生了显著变化[12]。但在应压木形成过程中,参与管胞壁主要成分合成的代谢物组分及其动态变化规律仍不明确。
杉木(Cunninghamialanceolata)是我国最重要的针叶用材树种之一,广泛分布于秦岭、淮河以南的16个省区,因生长速度快、树干通直、材质均匀等优良特性而具有较高的经济价值,是构成我国乔木林面积的优势树种之一。杉木与栎树、杨树等10个树种的人工林面积占我国乔木林面积的46.30%,是中国南方木材加工业原材料的主要来源[13]。开展杉木木材形成及其调控机制研究是杉木良种培育的重要内容,对提高杉木人工林效益有重要意义。因此,本研究利用应压木模型,以不同处理时间的杉木应压木为材料,对其管胞次生细胞壁结构、木质部主要组分及其代谢成分进行比较分析,以期揭示杉木应压木形成过程中主要代谢物的动态变化规律,为杉木木材形成及其调控机理研究提供理论依据。
1 材料与方法
1.1 植物材料与样品制备
本研究采用的材料为杉木无性系洋061的3年生植株,于2018年5月将植株斜放,使之与竖直方向成45°夹角诱导应压木形成。设置30、60和90 d 3个倾斜处理时间,处理结束后取样,所有样本均取自植株的相同高度与部位,其中斜放树干下侧为应压木(compression wood, CW)、斜放树干上侧为对应木(opposite wood, OW),同时取直立生长植株相同部位(直立木,control/straight wood)作为对照(CK)。将取样部位剪下后,快速剥掉皮层并去掉髓心,将余下的木质部组织快速放入预冷的无菌离心管,液氮速冻后转移至-80℃冰箱保存,用于成分的测定。每种木材样本均取3个生物学重复,共6株无性系参试苗。
1.2 试验方法
1.2.1 显微结构观测 取不同处理时期的茎段,浸没于2.5%戊二醛4℃固定软化过夜,用树脂包埋后利用RM2235切片机(徕卡微系统有限公司,德国)制成2 μm的切片,将切片用1%番红染色,经酒精梯度脱水后,用中性树胶封片后置于DM4000 M显微镜(徕卡微系统有限公司,德国)进行观测和拍照。使用ImageJ测量每张图像上的管胞壁厚,每个时期每个处理测50个样本。参照王杰等[14]、林金安等[15]的报道,利用多糖单克隆抗体LM5检测杉木管胞细胞壁半乳聚糖的分布,使用LSM880激光共聚焦扫描显微镜(卡尔蔡司光学有限公司,德国)进行图像的获取与处理。
1.2.2 木质部主要成分测定 将1.1中的材料置于干净无菌的研钵中,液氮研磨成粉末,并利用Alpha1-4真空冻干机(Marin Christ,德国)于-60℃进行冷冻干燥,然后利用范式洗涤纤维法对木质部中的酸性木质素、纤维素和半纤维素进行测定[16]。
1.2.3 主要代谢成分的提取及衍生化 代谢物提取和衍生化的方法参考石江涛等[17]和张胜龙等[18]的报道并稍加改进,具体如下:准确称取冻干粉50 mg于2 mL离心管中,加入1 400 μL 80%甲醇和25 μL 20 mg·mL-1核糖醇(内标物);置于Vortex-Genie 2振荡仪(Scientific industries, 美国)上振荡5 min充分混匀,于SY超声仪(新上海牌,上海)45℃恒温超声萃取1 h,然后12 000 r·min-1离心20 min,吸取上清液于15 mL离心管中;置于N-EVAP氮吹仪(Organo mation associates Lnc,美国)上吹干后,加入150 μL 4℃预冷的甲氧氨基盐酸盐(20 mg·mL-1)溶液,密封后置于摇床上37℃、160 r·min-1孵育2.5 h;然后加入150 μL N-甲基-N-(三甲基硅烷)三氟乙酰胺[N-methyl-N-(trimethylsilyl)trifluroacetamide, MSTFA]37℃、160 r·min-1孵育30 min;最后加入300 μL吡啶(色谱级)和10 mg无水硫酸钠混匀静止2 min,吸取400 μL衍生液并用有机相微孔滤膜(0.45 μm)过滤于气相瓶中用于气相色谱质谱(gas chromatography mass spectrometry, GC-MS)检测。
1.2.4 色谱条件与质谱条件 利用GC-MS对代谢物进行测定,在ISQ气质联用仪(赛默飞世尔科技有限公司,美国)进行分析。色谱柱:TR-5 MS石英毛细管柱(30 m×0.25 mm×0.25 μm),进样口温度:230℃;升温程序:初始柱温设为100℃保持3 min,以 2℃·min-1升至200℃后再以5℃·min-1升至300℃保持5 min;载气流速:1.5 mL·min-1;进样量:1 μL;分流比:15∶1。采用电子轰击离子源,传输线温度为250℃,离子源温度为200℃,扫描范围为40~600m/z,为确保衍生物能充分被离子化,设置2.5 min的溶剂延迟。
1.2.5 组分鉴定和相对定量方法 利用MS Workstation version 7.0软件对质谱图进行识别读取,将测定所得的化合物与国家标准与技术研究所(National Institute of Standards and Technology,NIST)质谱库进行全扫描匹配,相似度达80%以上并结合分子质量、分子结构以及质核比综合分析后,确认该化合物的化学成分。代谢成分的含量计算采用面积归一法,数值根据加入的内标含量计算。
1.3 数据分析
利用Excel 2019和SPSS 18.0软件进行测定数据的平均值和标准差计算以及显著性检验等分析,使用Origin 8.5和Photoshop CS6软件进行图表制作,参照刘振等[19]和罗钦等[20]的报道使用Simca 13.0软件进行正交偏最小二乘(orthogonal partial least squares discriminant analysis, OPLS-DA)主成分分析。
2 结果与分析
2.1 杉木应压木管胞细胞壁显微结构特征
斜放处理30 d后,首先对杉木应压木形成区域进行观察,结果显示,与对应木和直立木相比,杉木应压木圆盘明显不对称且产生了红棕色加厚区域(图1)。进一步对应压木的显微结构进行观察,发现应压木次生木质部管胞壁明显增厚,管胞多数为圆形或椭圆形,且细胞角隅胞间层(cell corner middle lamella, CCML)出现空腔(图2-A),而对应木和直立木的管胞主要是方形或多边形,其CCML中无空腔(图2-B~C)。对管胞细胞壁的木质素自发荧光进行检测,结果表明在对应木和直立木中,较强的木质素荧光主要出现在CCML,证明该壁层中木质素含量较高;而在应压木中,最强荧光信号则出现在靠近CCML的S2L(图2-D),显示此处木质素含量最高,远离CCML的壁层含量相对较低。使用多糖单克隆抗体LM5表位检测应力木和直立木中的(1,4)-β-半乳聚糖分布,应压木的S2L显示出丰富的半乳聚糖表位(图2-G),对应木、直立木中则是稀疏地分布在S1中(图2-H~I)。上述结果说明,相对于直立木和对应木,应压木细胞壁中化学成分原位分布发生了明显变化。3个时期的应压木典型特征基本一致,因此下文仅放置一个时期的图片。

注:A:斜放处理示意图;B:直立木横切面;C:应力木横切面;CW:应压木;OW:对应木;CK:直立木。
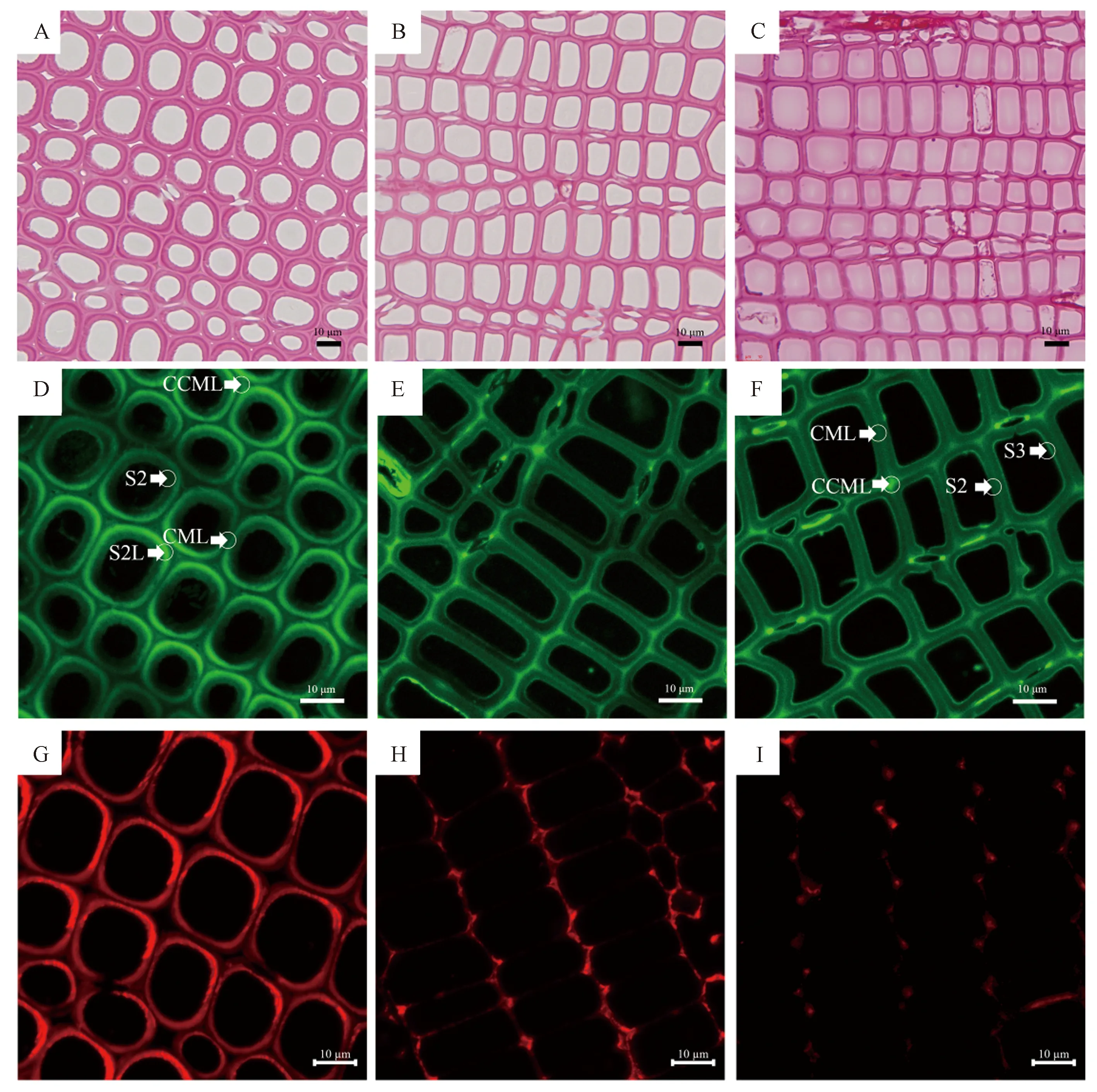
注:A~C:番红染色;D~F:木质素自发荧光;G~I:LM5表位的免疫定位;CCML:细胞角隅胞间层;CML:复合胞间层;S1~S3:次生壁S1~S3层;S2L:次生壁S2层的外层。
对应压木的双壁厚进行测定,结果如图3-A所示。应压木木质部的双壁厚极显著大于直立木,但60~90 d增厚不明显。
2.2 杉木应压木中木质纤维素分析
化学成分分析结果表明,3个处理时间的应压木木质素相对含量均极显著高于对应木和直立木,但不同时期的应压木间木质素含量无明显差异(图3-B);60和90 d应压木纤维素相对含量低于直立木,且低于同时期的对应木(图3-D),并随着斜放时间的延长呈现下降趋势;与木质素相同,同一时期应压木半纤维素相对含量显著高于直立木,并且随斜放时间的增加呈现上升趋势,但不同时期应压木的半纤维素含量无显著变化,除90 d时对应木中半纤维素相对含量显著高于直立木,其他时期对应木与直立木无明显变化(图3-C)。
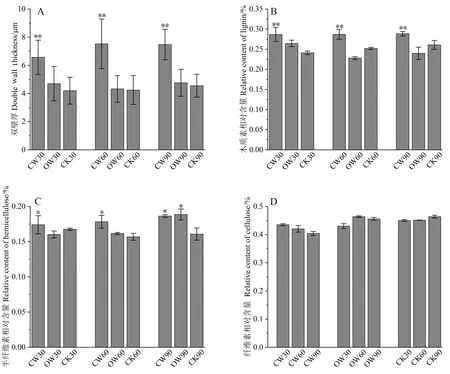
注:CW30~CW90分别表示处理30、60和90 d后的应压木;OW30~OW90分别表示处理30、60和90 d后的对应木;CK30~CK90分别表示处理30、60和90 d后的直立木;*:相对于CK显著;**:相对于CK极显著。
2.3 杉木应压木中差异代谢物的鉴定
利用GC-MS对杉木应压木、对应木和直立木的木质部代谢成分进行检测,结果均检测到39个色谱峰(峰面积大于0.1%)(图4)。对这39个色谱峰进行解谱分析,发现20个为已知的代谢成分(同分异构体算作一种),其相对应的峰面积占到总峰面积的90%以上;此外,有7个峰未能得到鉴定。对检测到的20种已知化合物进行分类,其中有机酸7种、单糖3种、二糖2种、糖苷类1种、醇类3种、酯类1种、氨基酸2种、酚类1种(表1)。谱图鉴定发现2个松醇和果糖的指示峰,这些化合物含有多个取代三甲基的位点,在未完全衍生化情况下导致异构体的产生,在色谱图上显示为2个峰,质谱库鉴定为同种化合物,因此定量分析时将其看作同一个峰。代谢谱显示多个蔗糖的指示峰,由于其细小致密,分析时合并为一个峰处理。进一步分析这些代谢成分在不同样本中的相对含量及变化规律可知,糖类物质含量较高,其中蔗糖相对含量个体最高可达到28.81%,其次是有机酸类和醇类,其他几种化合物含量相对较少。
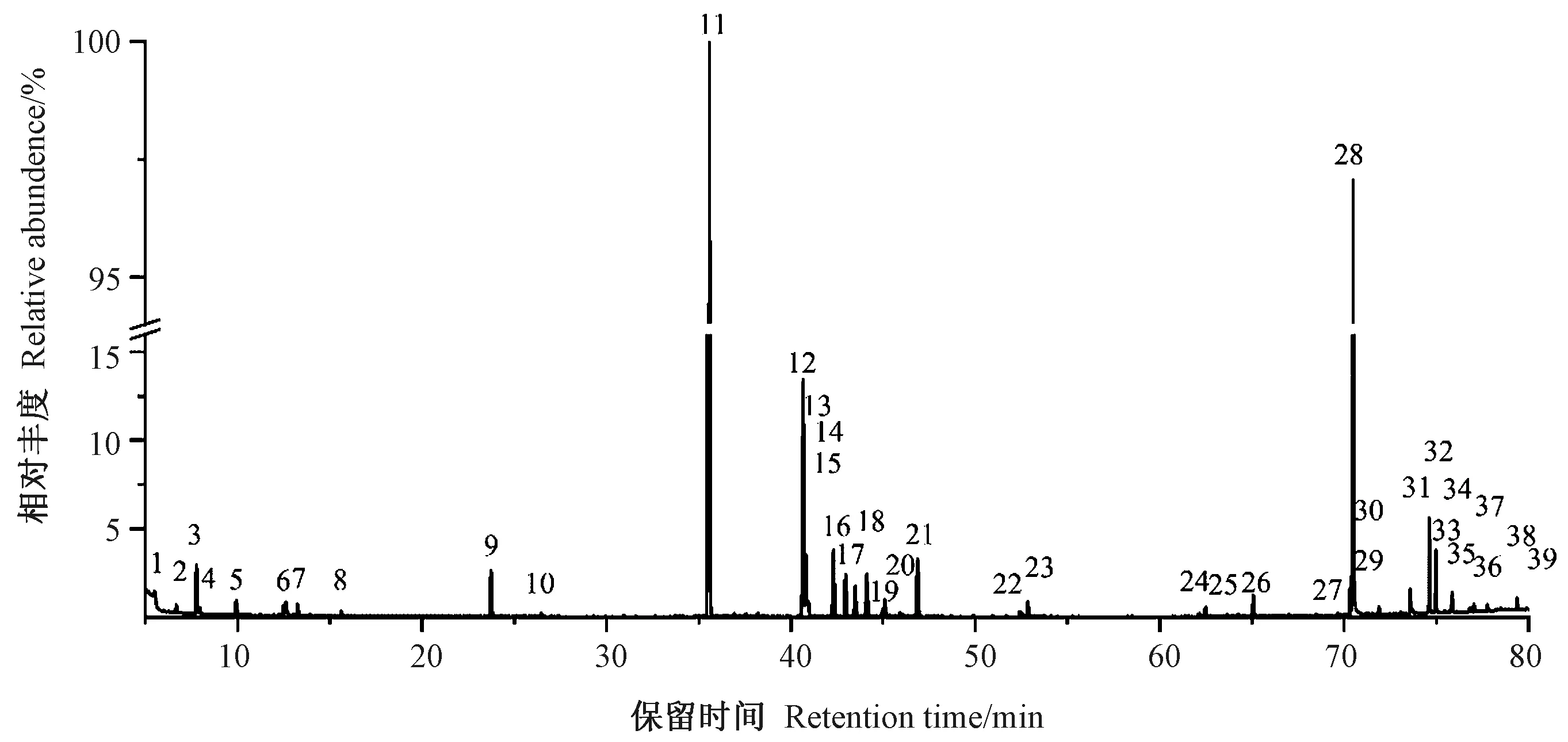
注:1:吡啶;2:吡啶;3:乙醇酸;4:缬氨酸;5:甘油;6:磷酸酯;7:硅烷醇;8:甘氨酸;9:苹果酸;10:未知化合物;11:核糖醇(内标);12:松醇;13:莽草酸;14:柠檬酸;15:奎宁酸;16:D-果糖;17:D-(-)-果糖;18:葡萄糖;19:半乳糖;20:核糖酸;21:未知化合物;22:松醇;23:棕榈酸;24:肌醇;25:未知化合物;26:异海松酸;27:未知化合物;28、29:蔗糖;30:未知化合物;31:儿茶素;32:儿茶素;33:乳糖1;34:未知化合物;35:乳糖2;36:β-甲基-D-半乳糖苷1;37:未知化合物;38:蔗糖;39:β-甲基-D-半乳糖苷2。
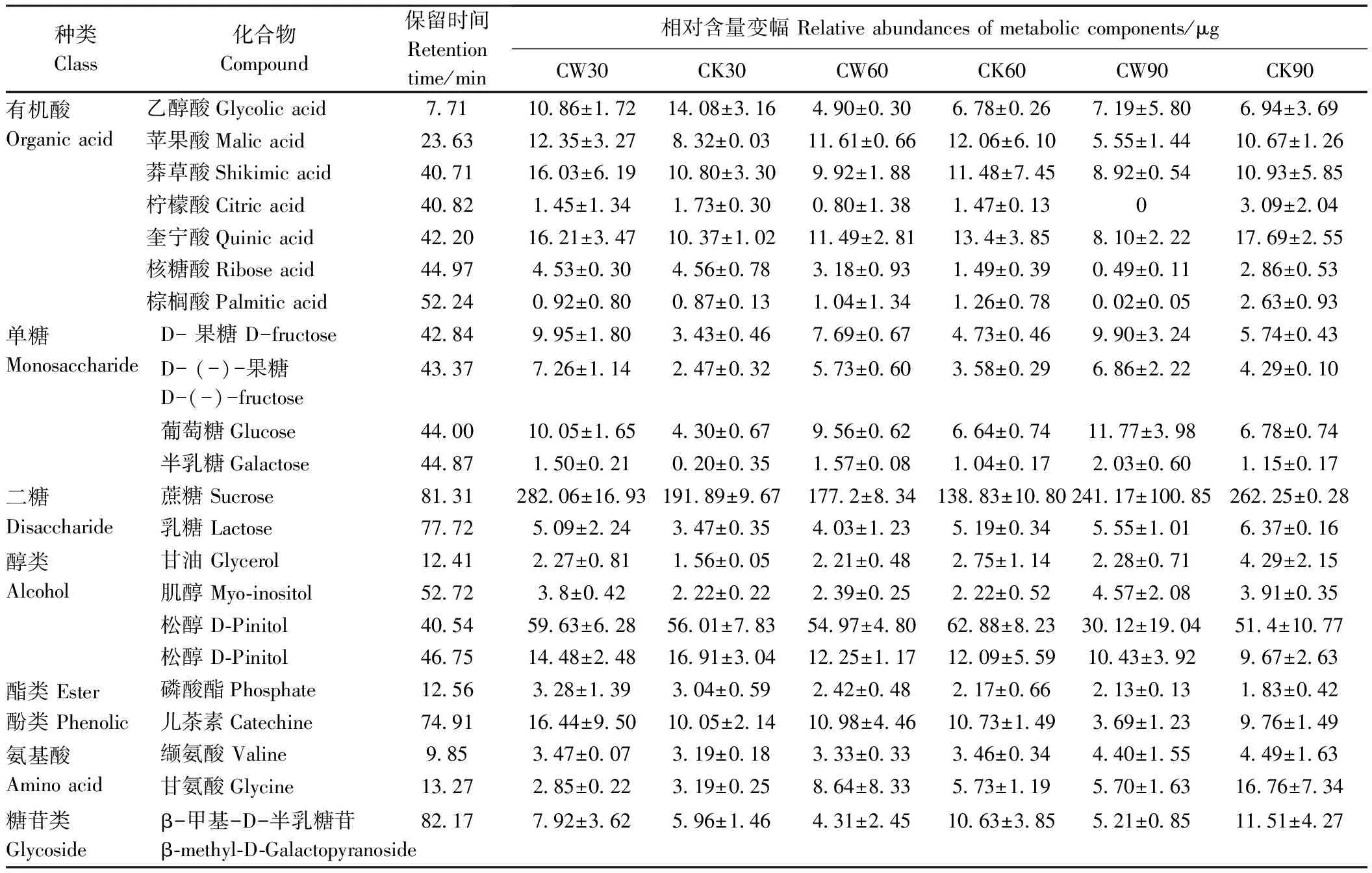
表1 3个时期主要代谢成分的相对含量变幅
根据图5结果对比应压木和直立木在30、60和90 d的代谢化合物相对含量,发现在杉木斜放处理30 d后,16种化合物的比值大于1,表明应压木中该化合物含量大于直立木中的含量。其中有10种化合物的相对含量比率,包括有机酸种类(苹果酸、奎宁酸、莽草酸和棕榈酸)、糖类(蔗糖和乳糖)、酚类(儿茶素)、两种氨基酸和β-甲基-D-半乳糖苷,斜放30 d后高于斜放60 d,直至90 d应压木含量低于直立木。属于糖类的果糖(将两种果糖合并为一种计算)、葡萄糖和半乳糖以及肌醇的相对含量比率在30 d时高达1.7以上,这几种化合物在60和90 d的应压木中虽然呈现下降趋势,但仍然处于高于同时期直立木的水平。应压木中乙醇酸在30和60 d低于直立木,90 d后含量略高于直立木。柠檬酸在3个时期均呈现应压木低于直立木的水平。甘氨酸与核糖酸在3个时期呈现倒“V”型的分布规律,30 d应压木中化合物含量与直立木相比无显著变化,60 d比值大于1.5,90 d低于0.4。松醇和磷酸酯在3个时期无明显变化。
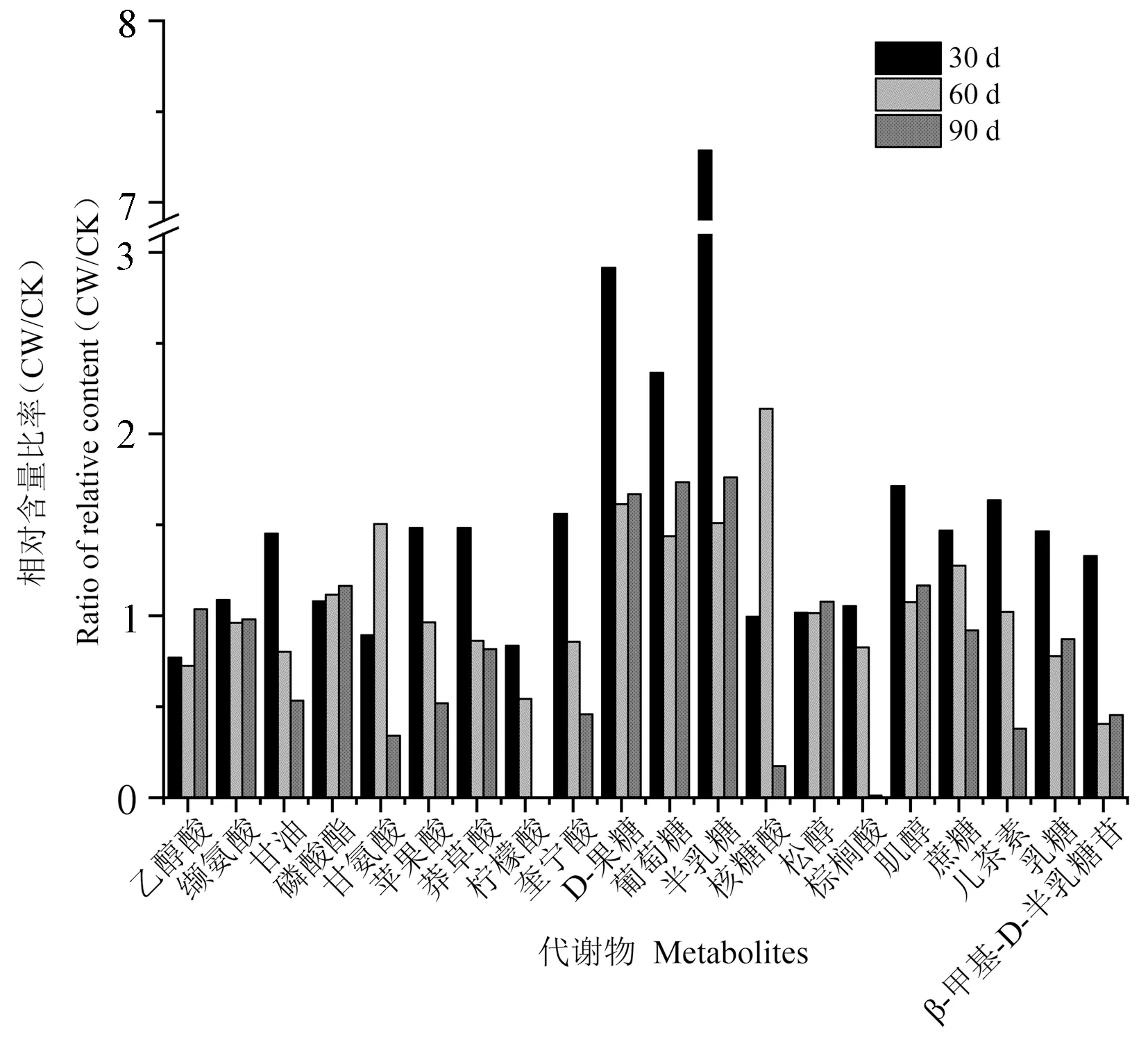
图5 杉木应压木(CW)和直立木(CK)在30、60和90 d的相对含量比率
3 讨论
3.1 杉木应压木的解剖结构
应压木是裸子植物偏离垂直方向生长时,为了抵抗重力吸引,在茎下部产生的一种具有特殊结构和成分的木材组织,被认为是研究木材形成的良好模型[21]。本研究利用该模型对杉木应压木的木质部解剖特性和主要代谢成分变异规律进行了系统研究,结果表明杉木仅经30 d的斜放诱导处理后,应压木木质部管胞壁显著增厚,是直立木及对应木管胞双壁厚的1.5倍,且出现木质化程度较高的S2L层,CCML处出现空腔,无S3层。该结果与马尾松[22]、火炬松[23]、云南松[10]和日本柏树[24]等众多裸子植物的应压木结构变化一致,皆是次生壁增厚,微纤丝夹角发生了改变,管胞出现螺纹裂痕,S3缺失。根据木质素自发荧光且荧光强度与木质素浓度呈线性正比的现象,利用高分辨荧光图像检测杉木不同壁层的木质素原位分布,本研究结果表明,相比对应木和直立木,应压木S2L与S2层的木质素含量均有升高。已有用细胞壁成分有特异性的抗体对辐射松应压木进行免疫杂交的研究,结果发现S2L层沉积了(1,4)-β-半乳聚糖,S2层出现阿拉伯半乳聚糖蛋白沉积,以及管胞螺纹裂隙周边出现(1,3)-β-葡聚糖的沉积[6,25]。杉木的半纤维素表位与辐射松相似,S2L处沉积了(1,4)-β-半乳聚糖,对应木和直立木中仅在S1层周围出现。纵向对比3个时期的应压木变化,发现应压木形成过程中的解剖变化不显著,不同时期之间变化趋势类似。
3.2 杉木应压木形成中的化学成分含量变化
测定结果显示,杉木应压木双壁厚随斜放时间的增加逐渐增厚,而纤维素相对含量呈现下降趋势,因此推测应压木管胞次生壁增厚的原因极有可能与木质素以及半纤维素如半乳聚糖、阿拉伯半乳聚糖蛋白、葡聚糖等在S2外层异常沉积有关。对应木的双壁厚虽然与直立木无明显差异,但其木质素相对含量随斜放时间的增加呈现下降趋势,而纤维素相对含量则呈现上升趋势(图3),由于对应木和应压木来源于同一棵树的对立两边,对应木的管胞壁显微特征虽然与直立木无明显差异,但其管胞次生壁化学成分在斜放诱导期间可能发生了与应压木相反的变化。
3.3 杉木应压木形成中主要代谢成分分析
在木材形成过程中,纤维素、木质素与半纤维素的合成涉及复杂的代谢途径和代谢组分。通过对比应压木和直立木的代谢谱,发现在木质部当中主要是糖类物质富集,表明木材形成是以糖代谢为主的木质纤维素生物合成。木质素的生物合成普遍认为可分为以下三个步骤:首先是光合作用后的同化产物到芳香氨基苯丙氨酸、酪氨酸和色氨酸等的合成过程,称为莽草酸途径;然后是从苯丙氨酸到羟基肉桂酸及其辅酶A酯类的合成过程,称为苯丙酮酸途径;最后是羟基肉桂基辅酶A酯类到木质素单体及其聚合物的过程,称为木质素合成的特异途径[26]。研究发现在莽草酸合成途径中脱氢奎宁酸和莽草酸脱氢酶两种酶相互融合,共同促进了莽草酸途径的第3和第4个阶段,增加了代谢物流通的效率[27]。苹果酸参与植物体内多个代谢途径,其中苹果酸酶以苹果酸为前体催化反应,为肉桂醇脱氢酶提供烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate, NAPDH)生成木质素单体[28]。因此,莽草酸、奎宁酸、苹果酸是与木质素合成相关的代谢成分[29-30]。经过30 d的斜放诱导处理,杉木应压木中的与木质素合成相关的物质如苹果酸、奎宁酸、莽草酸相对含量高于同时期的直立木,证明应压木木质化程度高于直立木(表1)。30 d应压木木质素相关成分水平接近60 d的直立木,表明在应压木形成的初期,代谢通量主要流向苯丙烷代谢途径,此结果与30 d后应压木中木质素含量迅速累积的现象一致。
碳水化合物包括半纤维素和纤维素,尿苷二磷酸葡萄糖(uridine 5′-diphospho glucose, UDPG)是纤维素与部分半纤维素生物合成的直接起始底物,其中半乳聚糖、半乳糖醛酸、木聚糖、阿拉伯半乳聚糖和葡甘聚糖的合成,均以UDPG为前体通过不同途径转化成尿苷二磷酸木糖、阿拉伯糖、乳糖等为单体而构建[31-32]。已有研究表明半乳聚糖、甘露聚糖和木聚糖等多糖的分布与最终木质化程度有关[33]。本研究代谢谱解析结果显示了杉木应压木形成过程中半乳糖、葡萄糖等半乳聚糖和葡甘聚糖合成底物的增加,分析发现相比直立木以及30和60 d的应压木,90 d后的应压木的代谢产物主要富集了糖类物质。该现象与应压木双壁厚逐渐增厚、半纤维素含量逐渐增加相关,上述糖类物质可能是决定木质素沉积在管胞细胞壁各层中的位置和数量的控制因素。本试验结果发现应压木中果糖、蔗糖等糖类化合物的相对含量均随斜放时间的增加而降低(图5),其中蔗糖在应压木90 d后已下降至低于直立木的水平,而单糖物质果糖、葡萄糖和半乳糖仍处于高于直立木的水平,表明在应压木90 d后,蔗糖酶与蔗糖合酶的表达极有可能仍处于上调水平,其将蔗糖转化为葡萄糖与果糖,增加了UDPG与半纤维素合成的效率。这可能是由于木质素沉积的模式在生长季节保持不变,夏季木质化细胞数量达到峰值后逐渐降低造成的[33]。柠檬酸既能在基础生命代谢中起中间产物的作用,又能改进应压木的抗压强度[34]。本试验结果发现柠檬酸在3个时期均呈现低于直立木的水平(表1),而与此相关的原因还未有相关结论,有待进一步研究。
4 结论
杉木应压木形成中呈现出管胞变圆,S2L增生以及管胞次生壁增厚,木质素与半纤维素含量显著增加等现象。GC-MS检测到共有20种化合物,其中有机酸7种、单糖3种、二糖2种、糖苷类1种、醇类3种、酯类1种、酚类1种,应压木中差异性代谢物相对含量显著高于直立木。在应压木形成初期,即30 d斜放处理诱导后,有机酸类和糖类物质大量在木质部累积,为木质素合成和细胞壁增厚提供了代谢中转产物。当斜放诱导时间增长至90 d后,有机酸类物质含量下降至低于直立木的水平,为保证半纤维素的合成,单糖类物质的含量仍处于高于直立木的水平,表明木质素合成是应压木形成初期的主要反应,半纤维素的合成速率则随着诱导时间的延长而逐渐提高。
猜你喜欢
化工进展(2022年9期)2022-10-13
化工进展(2022年7期)2022-08-01
防护林科技(2022年2期)2022-04-25
疯狂英语·读写版(2020年11期)2020-12-21
农民致富之友(2020年8期)2020-05-11
发明与创新·中学生(2019年10期)2019-11-17
中国造纸(2019年6期)2019-09-10
农家科技下旬刊(2019年12期)2019-01-13
现代装饰·理论(2016年9期)2016-12-26
企业文化·中旬刊(2014年12期)2014-12-30
