
粒毛盘菌多糖在不同消化阶段的生物活性
2022-10-21 00:51:58张鑫淼何雅玲
生物学杂志 2022年5期
张鑫淼, 何雅玲, 叶 明
(合肥工业大学食品与生物工程学院, 合肥230009)
真菌多糖是一类天然高分子碳水化合物,主要存在于发酵液、菌丝体及子实体中,因其所具有的抗肿瘤、免疫调节等一系列显著的药理学活性,在食品、药品等领域有较好的应用潜能[1-2]。多糖的生物活性很大程度上取决于其理化性质及结构特性,但胃肠消化的复杂环境如pH、温度、酶等都可能对活性结构产生较大影响。因此,确定多糖在胃肠道消化过程中的变化对准确评估其生物活性和生物可及性具有重要意义[3]。
近年来,关于多糖消化特性的研究引起国内外研究人员广泛的关注[4]。消化是人体代谢吸收的重要生理过程,它能将生物大分子转化为能被机体吸收的小分子物质,进而通过代谢发挥功能。然而,体内试验由于技术难度大,成本昂贵和伦理限制等难以进行。静态体外消化模型是通过模拟真实生理消化条件预测食物或药物在体内的变化及生物可及性的有效方法[5],因其高效、经济、安全等显著优势逐渐应用于多糖消化特性的评估。
粒毛盘菌(Lachnum)属于晶杯菌科(Hyaloscyphaceae),是腐生类的高等真菌,其物种丰富,深层发酵可以产生多种代谢产物[6]。近年来,研究主要从不同种粒毛盘菌的发酵液中提取得到多糖,对其结构、生物活性等进行表征及探究。研究表明,粒毛盘菌多糖具有抗氧化[7]、抗肿瘤[8]、肝保护[9]等多种重要的生物活性,有较好的研究开发前景。然而,粒毛盘菌多糖在模拟唾液及胃肠道消化条件下的消化特性、生物活性变化尚未有相关报道。
研究通过体外消化模型在接近真实生理消化条件下探究粒毛盘菌多糖的消化特性,如分子质量、还原糖、微观结构等变化,并探究其在消化过程中抗氧化活性的变化及其对脂解的影响,为粒毛盘菌多糖体内消化吸收的探究提供理论基础,有助于其在功能食品添加剂方面的开发应用。
1 材料和方法
1.1 材料
粒毛盘菌(Lachnum)YM40菌株子实体分离自黄山(安徽,中国),保藏在合肥工业大学微生物资源与应用实验室。DEAE Cellulose-52购自合肥博美生物;Sephadex G-100购自北京索莱宝科技有限公司;单糖标品、葡聚糖标品购自美国Sigma公司;3,5-二硝基水杨酸(DNS)、2,2-联苯基-1-苦基肼基(DPPH)、2,2′-联氮-双-3-乙基苯并噻唑啉-6-磺酸(ABTS+)、胰酶(来自猪胰腺)购自阿拉丁科技有限公司;胃蛋白酶购自德国BioFroxx公司;研究中使用的所有其他试剂均为分析纯级试剂。
1.2 仪器与设备
LC-10A高效液相色谱仪(配有RI-502示差检测器和BRT105-104-102串联凝胶柱),日本岛津公司生产;ICS5000+离子色谱仪和6700傅里叶红外光谱仪,美国赛默飞世尔科技公司生产;723N(PC)可见分光光度计,上海悦丰仪器仪表有限公司生产。
1.3 方法
1.3.1 粒毛盘菌多糖的提取纯化
粒毛盘菌多糖的提取纯化参考文献[6]。粒毛盘菌发酵液经抽滤、减压浓缩后醇沉。沉淀复溶后用Sevage法除蛋白、H2O2脱色、透析、冷冻干燥即得粗多糖LEP。将LEP溶液以0.1 mol/L NaCl作为洗脱液用DEAE Cellulose-52柱(2.6 cm×40 cm)洗脱得LEP-2。用Sephadex G-100柱(1.5 cm×60 cm)对LEP-2进一步纯化得到主要组分,冷冻干燥得纯多糖LEP-2a。
1.3.2 LEP-2a的体外模拟消化
(1)唾液对LEP-2a的消化作用。唾液由4名20~30岁的健康志愿者提供[10]。收集的唾液在6 400 r/min下离心10 min得上清液。将LEP-2a溶液(4 mg/mL)与人唾液,唾液与蒸馏水分别按照1∶1混合,37 ℃水浴中孵育消化。分别在5、30、60 min取样,采用沸水浴灭酶5 min。调整消化液的pH值为7,然后9 700 r/min离心10 min取上清液。取唾液消化60 min的样品透析后冷冻干燥,命名为LEP-2aG。
(2)模拟胃液对LEP-2a的消化作用。模拟胃液的配置参考文献[11]。配制2 L胃液电解质溶液,包含2.2 g KCl,6.2 g NaCl,0.3 g CaCl2·2H2O和1.2 g NaHCO3。取1 500 g胃电解质溶液加入400 mg胃蛋白酶及3 mL CH3COONa溶液(1 mol/L,pH 5),0.1 mol/L HCl将其pH值调至3得到配制的胃液。将模拟胃液与 LEP-2a溶液(4 mg/mL),蒸馏水与模拟胃液分别以1∶1的比例混合,37 ℃水浴振荡。消化期间保持pH值为2.5。分别在5、30、60、120、180、240 min时取样。取胃液消化240 min的样品命名为LEP-2aS。样品处理方法与(1)相同。
(3)模拟肠液对LEP-2a消化作用。模拟肠液的配置参考文献[11]。1 L肠电解质溶液包含0.65 g KCl,5.4 g NaCl和0.33 g CaCl2·2H2O。取500 g肠电解质溶液加入500 g胰酶溶液(质量分数为7%)、1 000 g胆酸盐溶液(质量分数为4%)和0.13 g胰蛋白酶,充分搅拌混匀之后,用NaOH (0.1 mol/L) 调pH值为7.0。将LEP-2a、蒸馏水模拟胃液消化得到的混合物用NaOH调整pH值为7后,分别与模拟肠液以10∶3的比例混合,37 ℃水浴中孵育。消化期间,保持反应溶液pH值为7.0。分别在消化的5、30、60、120、180、240 min时取样。肠液消化240 min的样品命名为LEP-2aI。样品处理方法与(1)相同。
1.3.3 LEP-2a及其消化过程中理化性质分析
(1)主要化学成分分析。总糖的含量采用苯酚-硫酸法[12]测定。Bradford法[13]测定蛋白质含量。糖醛酸含量用间羟基联苯法[14]测得。单糖组成参考文献[15]对样品处理并通过赛默飞ICS-5000+ HPIC高压离子色谱色谱系统测定。
(2)分子量测定。在岛津LC-10A高效液相色谱上采用高效凝胶渗透色谱(HPGPC)法[16]测定多糖分子量。柱温保持在40 ℃,以0.05 mol/L NaCl为流动相,流速为0.6 mL/min,样品进样量为20 μL。根据葡聚糖标品的校正曲线计算样品分子量。
(3)模拟消化过程中还原糖含量测定。分别取1.3.2节唾液、胃液、肠液不同阶段的消化样品的上清液,用3,5-二硝基水杨酸(DNS)法[17]测定还原糖含量。取0.5 mL DNS溶液加入0.5 mL样品溶液中摇匀,沸水浴5 min,冷却后加入4 mL蒸馏水,测定540 nm处的吸光度。以蒸馏水替代多糖在不同阶段的消化液作参比溶液。还原糖标准曲线以葡萄糖为标品按照上述方法测得。根据不同阶段消化液中LEP-2a含量计算相对还原糖含量(%)。
(4)形态特性。用原子力显微镜(AFM)观察多糖组分的微观构象。取5 μL完全溶解的样品溶液(20 μg/mL)滴在新切割的云母表面并在室温下干燥。采用轻敲模式获得AFM 图像[18]。
1.3.4 模拟消化过程中LEP-2a抗氧化活性
根据前人报道的方法测定多糖体外模拟消化过程中对DPPH[19]和ABTS+[20]自由基的清除活性及其还原能力[21]变化。分别以不同消化阶段的消化液作为参比溶液以消除消化液自身抗氧化活性的影响。
1.3.5 模拟消化条件下LEP-2a对脂质分解的影响
用参考文献[22]的方法,将20%的玉米油、吐温80溶液(质量分数为2.5%)与不同含量的多糖溶液混合,高速均质制备多糖-脂质乳液(0.5%、1.0%)。同时,以去离子水代替多糖溶液作空白对照。体外模拟消化模型测定多糖对消化脂解的影响。胃液和肠液的制备及对多糖-脂质乳液的体外模拟消化过程均采用1.3.2所述方法实施。其中,模拟胃和肠消化均为100 r/min连续搅拌2 h。反应过程中监测并维持唾液消化时恒定pH 6.8、胃消化时pH 2.5和肠消化时pH 7.0所需的NaOH溶液的量。根据NaOH的消耗量计算反应过程中游离脂肪酸(FFA)的释放量:
其中:VNaOH表示NaOH消耗的体积;CNaOH表示NaOH的浓度,即0.1 mol/L;MWlipid表示玉米油的平均分子质量,即872 g/mol;Wlipid表示反应液中玉米油的质量。
1.3.6 统计学分析
所有实验重复3次,数据表示方式:平均值±标准差。采用SPSS软件进行单因素方差分析(ANOVA),评价差异显著性。P<0.05表示存在显著性差异。
2 结果与分析
2.1 LEP-2a及其消化组分的化学组成
LEP-2a及其消化组分的化学组成如表1所示。LEP-2a是分子质量约为50.3 ku的杂聚糖,总糖含量超过99%,糖醛酸含量约为1.48%±0.16%,未检测到蛋白质。模拟消化后糖醛酸含量升高,Yuan等[3]对Holothurialeucospilota多糖消化研究中也观察到了这一现象。单糖测定结果表明,LEP-2a及其消化组分主要由甘露糖、半乳糖、葡萄糖及半乳糖醛酸组成,但胃肠消化后葡萄糖及甘露糖相对含量降低,可能与其独特化学结构有关。此外,LEP-2a的单糖组成和分子量与以前的研究均不相同[6,9,23-24],表明它是一种新的多糖。
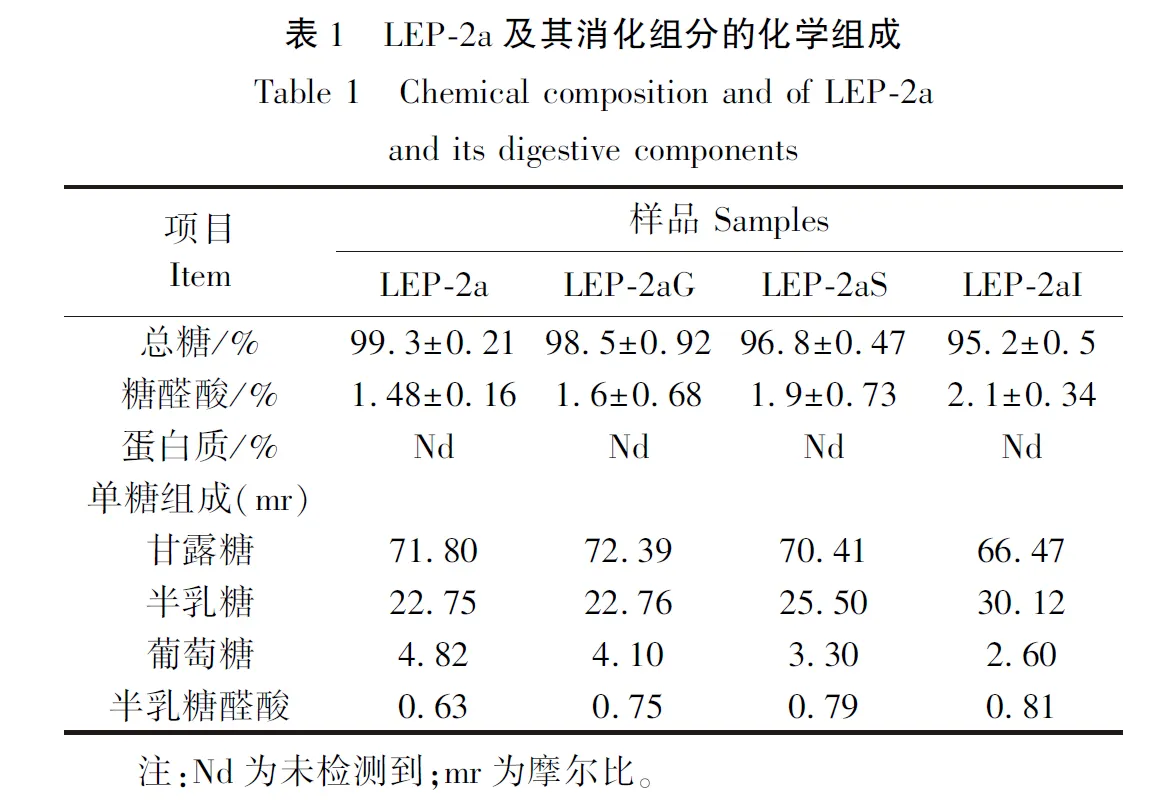
2.2 LEP-2a及消化组分的分子质量分析
LEP-2a及其消化组分的分子质量(Mw)如图1所示。以不同分子质量的葡聚糖作标品,得到分子质量校正曲线方程:y=-0.220 8x+13.29(R2=0.9905),其中y为lgMw,x为保留时间。唾液消化后LEP-2a的保留时间及平均分子质量未发生明显变化,表明LEP-2a不能被唾液消化降解。胃肠消化后,LEP-2a的保留时间由38.897 min延长到39.02 min和39.235 min,其分子质量也从50.30 ku下降到47.25 ku和42.36 ku,表明胃肠消化使LEP-2a发生了降解[25]。
图1 LEP-2a及其消化组分的分子质量分布Figure 1 Molecular weight distribution of LEP-2a and its digestive components
2.3 消化过程中还原糖变化
多糖通常作为聚合物存在于水溶液中,糖苷键断裂可使其还原端数量增加,分子质量降低[26]。因此,测定消化过程中还原糖含量变化判定LEP-2a的降解原因。以葡萄糖为标品测得还原糖标准曲线为y=1.607 4x+0.002 6,R2=0.998 2;其中,x为葡萄糖质量浓度(mg/mL),y为OD540。如图2所示,唾液消化过程中LEP-2a的还原糖含量无显著性差异,分子质量也未发生明显变化,表明LEP-2a不能被唾液消化。还原糖含量在模拟胃液消化过程中缓慢增加,在肠液消化阶段迅速且显著增加(P<0.05)。在模拟肠液消化180 min时,还原糖含量增至10.55%±0.33%。这些结果与胃肠消化后分子质量显著降低相一致,表明LEP-2a主要在胃肠中被降解。
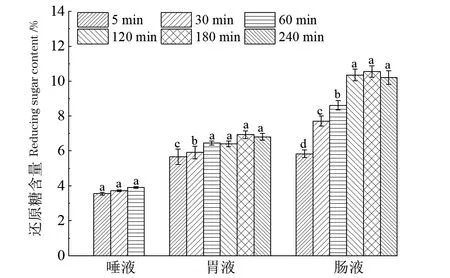
误差线为标准差;不同字母表示显著性差异(* P<0.05)。图2 体外唾液、模拟胃和肠液消化对LEP-2a还原糖含量的影响Figure 2 Effects of in vitro digestion (saliva, simulated gastric and intestinal) on reducing sugar content of LEP-2a
2.4 微观形貌变化
原子力显微镜(AFM)是表征多糖微观形貌的工具。LEP-2a及其消化产物的AFM平面及三维图像如图3所示。AFM检测到LEP-2a的微观高度为2.5 nm,高于单个多糖链(0.1~1.0 nm)[27],表明具有分支结构。唾液消化物LEP-2aG的高度为2.4 nm,与LEP-2a相比未发生明显变化,推测其受唾液消化影响较小。胃消化产物LEP-2aS和肠消化产物LEP-2aI的高度显著降低,高度分别为1.8 nm和0.8 nm。结合还原糖及分子质量变化,推测可能是胃肠消化使LEP-2a的糖苷键断裂,从立体网状结构变为链状结构,分支程度降低。微观形貌更直观地反映了LEP-2a主要在胃肠消化阶段被消化分解。
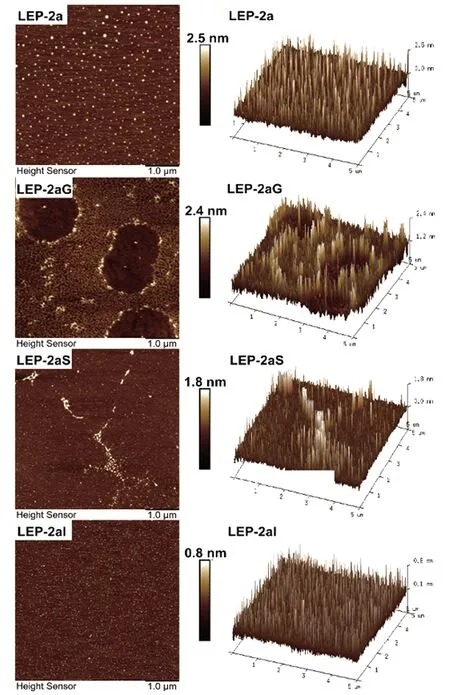
图3 LEP-2a及其消化组分的原子力显微镜图像Figure 3 Atomic force microscopy images of LEP-2a and its digestive component
2.5 消化过程中多糖抗氧化活性的变化
胃肠消化过程中多种因素如pH、温度、酶等可能使多糖结构发生改变,进而影响其抗氧化活性。如图4所示,LEP-2a对ABTS+和DPPH自由基的清除能力及其还原能力在唾液消化过程中无显著变化,这与其还原糖及分子质量在唾液消化阶段保持稳定相一致。LEP-2a对ABTS+的清除能力[图4(a)]随着模拟胃液和肠液消化的时间延长缓慢增强,在肠液消化阶段120 min时达22.5%±0.47%,随后趋于稳定(P<0.05)。LEP-2a对DPPH[图4(b)]具有良好的清除能力且胃液和肠液的消化对清除能力的影响较显著(P<0.05),在胃液反应240 min时清除能力达52.6%±1.36%。还原能力通常与防止过氧化物的某些前体反应和过氧化物的形成有关[28],是评估多糖潜在抗氧化活性的重要指标。LEP-2a具有一定的还原力且随着胃液消化时间的增加而增强[图4(c)],在肠液消化60 min时趋于稳定。因此,LEP-2a在体内依然保持较好的抗氧化活性,受唾液消化影响较小,但模拟胃肠消化阶段随消化时间的延长,抗氧化活性增强。
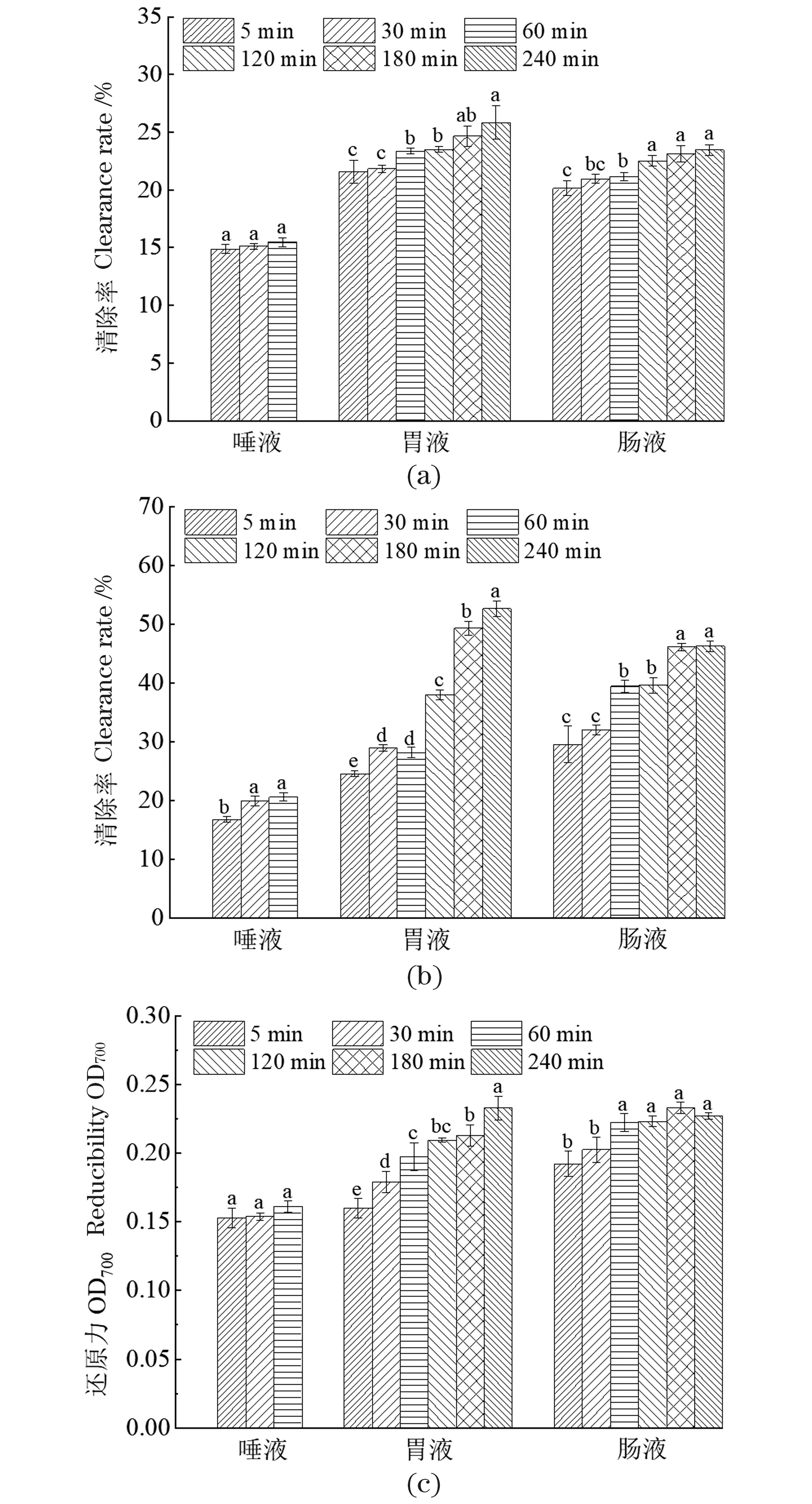
(a)ABTS自由基清除活力;(b)DPPH自由基清除活力;(c)还原力。误差线为标准差;不同字母表示显著性差异(P<0.05)。图4 体外唾液、模拟胃和肠液消化对LEP-2a抗氧化活性的影响Figure 4 Effects of in vitro digestion (saliva, simulated gastric and intestinal) on antioxidant activities of LEP-2a
2.6 消化脂解
不同含量的LEP-2a溶液(0、0.5%、1.0%)对胃肠消化阶段脂质分解的影响如图5所示。不添加LEP-2a时,脂质乳液在胃液消化环境下[图5(a)]被迅速分解释放游离脂肪酸(FFA)。在胃消化120 min时,空白组有6.30%±0.12%的游离脂肪酸释放并仍呈上升趋势。相比之下,LEP-2a组的脂质分解在胃消化40 min时即趋于稳定,且消化120 min时加入1% 多糖的乳液仅有0.34%±0.03%的脂质被分解。此外,模拟肠液消化120 min时,空白对照组有25.73%±0.26%的游离脂肪酸释放,但加入1%多糖的实验组仅有17.01%±0.29%的脂质分解。尽管实验中在消化过程中FFA释放相对较低,但结果与以前的工作一致[29],释放的FFA量低可能与脂质乳液粒径及玉米油的添加量有关。因此,LEP-2a的加入可以有效抑制脂质在胃肠中的消化分解。
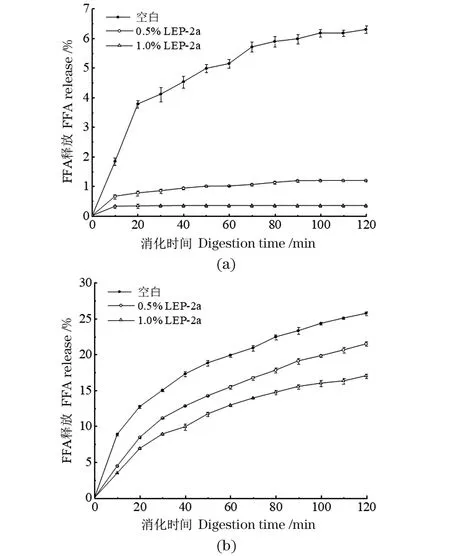
(a)模拟胃消化过程中FFA释放;(b)模拟肠消化过程中FAA释放。图5 模拟消化条件下多糖对脂质分解的影响Figure 5 Effect of polysaccharides on lipolysis under simulated digestion
3 讨论与结论
研究采用体外模拟消化模型(唾液、胃液及肠液)探究粒毛盘菌YM40多糖(LEP-2a)的消化特性及不同消化阶段的生物活性。研究表明,LEP-2a在体外唾液消化阶段理化性质及微观结构变化不显著,张冠亚等[30]在对铁皮石斛多糖的研究中也观察到了这一现象。事实上,大多数天然非淀粉多糖都不能被人唾液消化[26]。但在胃液和肠液消化过程中分子质量降低、还原糖含量增加(P<0.05)、糖醛酸含量升高、微观高度明显降低,表明胃肠消化使LEP-2a发生降解。这一变化可能是模拟胃液的酸性pH及肠液中的酶、温度等因素使糖苷键断裂[31]。不同真菌多糖的消化特性存在差异,Ding等[32]将灵芝多糖在体外模拟消化和体内发酵后发现该多糖在消化道中不能完全降解,主要在大肠中被分解和利用。Wang等[25]发现桦褐孔菌(Inonotusobliquus)多糖经胃肠模拟消化后分子质量由105.02 ku降低至36.74 ku,抗氧化及降血脂活性增强。对多糖消化特性的研究有利于探究其益生元特性,为进一步体内活性的研究提供基础。
多糖的抗氧化活性通常与其分子质量、糖醛酸含量、链构象等密切相关[3]。LEP-2a在模拟胃液和肠液消化过程中抗氧化活性随时间延长而增强,但受唾液消化影响较小。Miao等[34]研究发现,在一定程度上降低分子质量可以增强多糖的抗氧化活性。此外,多糖中存在的醛酸基团可以激活异构碳的氢原子进而增强多糖的供氢能力[20]。因此,模拟胃肠消化过程中LEP-2a抗氧化能力升高,可能归因于消化条件下糖苷键断裂使还原端数量增加,分子质量降低[34],糖醛酸含量增加,提高了供氢能力。
此外,LEP-2a可以较好地抑制脂质在模拟胃肠消化环境中迅速分解,可能由于多糖的加入增加了消化液的黏度,促进脂滴絮凝,从而限制脂肪酶与脂滴的接触[22]。但多糖对脂质消化影响的确切机制有待进一步研究。LEP-2a对脂质消化分解的抑制能力有利于降低餐后血脂水平的迅速升高,增强饱腹感,可以应用于富含油脂的食品中以促进健康。
LEP-2a主要在胃肠消化阶段因糖苷键断裂发生降解,经胃肠消化后抗氧化性增强且该多糖能有效抑制脂肪的消化分解。研究探索了粒毛盘菌多糖的消化特性,为其体内活性研究及进一步开发应用提供基础。后续将对该多糖进行结构鉴定并分析构效关系,进一步揭示粒毛盘菌多糖在人胃肠消化过程中的消化及吸收特点。
猜你喜欢
西南大学学报(自然科学版)(2022年12期)2022-12-20 02:32:14
中国食用菌(2020年8期)2020-11-08 13:41:50
科学Fans(2020年3期)2020-05-13 14:30:32
中成药(2018年10期)2018-10-26 03:41:28
生物技术通报(2018年4期)2018-03-31 17:16:11
现代检验医学杂志(2016年1期)2016-11-12 13:20:00
兽医导刊(2016年12期)2016-05-17 03:51:32
动物营养学报(2015年10期)2015-12-01 02:26:20
现代检验医学杂志(2015年5期)2015-02-06 01:42:35
现代医药卫生(2014年18期)2014-03-11 19:33:20
