
黑沙蒿醇提物对肉仔鸡生长性能、免疫指标及相关基因表达量的影响
2022-10-21 02:46:44郭燕菲石璐璐乌力雅素郭文亮杜海东徐元庆史彬林
中国兽医学报 2022年8期
郭燕菲,石璐璐,乌力雅素,郭文亮,杜海东,金 晓,徐元庆,史彬林
(内蒙古农业大学 动物科学学院,内蒙古 呼和浩特 010018)
抗生素是过去几年提高畜禽生产效益的常见添加剂,但因其累积效应会造成畜禽自身免疫力下降和病毒耐药性增强,近年来抗生素已被明令禁止使用,因此寻找绿色、安全,能够改善畜禽机体健康状态的饲料添加剂已成为研究的重点。中草药植物因其分布广泛、资源丰富且含有多种有效活性成分成为替代抗生素的最佳选择。已有研究表明,中草药饲料添加剂在促进畜禽生长发育、提高其生产性能方面有着积极作用[1]。目前,国内外常用的植物活性物质提取方法有水提法以及醇提法,以此可得到较高含量的黄酮、多糖以及酚类化合物,已有研究表明,这些活性物质可以提高动物增重水平,促进动物生长,增强炎性因子的释放,提高免疫器官指数,是改善畜禽健康的天然高效免疫调节剂[2-4]。
黑沙蒿(Artemisia)俗称沙蒿,为菊科蒿属植物,其分布极为广泛,是我国北方地区主要的灌木之一。黑沙蒿味辛苦,性微温,全草均可入药,其茎叶可用于抗炎和消肿,根可用于止血,而且富含蛋白质、脂肪、维生素和微量元素等营养成分[5]。近些年来已有研究报道,黑沙蒿作为饲料添加剂对动物可起到抗病、保健、促进生长、提高免疫功能的作用[6],但基本集中于其水提物上,而有关黑沙蒿醇提物(Artemisiaordosicaethanol extract,AOEE)的研究鲜有报道。本研究以AOEE作为饲料添加剂添加到肉仔鸡饲粮中,探讨不同添加剂量的AOEE对肉仔鸡生长性能和免疫功能的影响,从而得出AOEE在肉仔鸡饲粮中的最适添加量,旨在为AOEE在动物生产中的推广应用提供参考。
1 材料与方法
1.1 实验动物与材料实验动物选取体质量(37.35±0.23)g相近的240只1日龄AA肉鸡,雌雄各半,于2020年8月26日至10月7日饲养于内蒙古农业大学动物科学学院动物营养代谢室。
试验材料:黑沙蒿采自内蒙古鄂尔多斯,自然阴干后粉碎,经石油醚萃取脱色脱脂,采用超声波辅助乙醇浸提处理,以60%乙醇为提取溶剂,料液比为 1∶30,在50℃、200 W功率下超声1 h,将提取液浓缩冻干即为AOEE,经NaNO2-Al(NO3)3-NaOH分光光度法[7]测定,AOEE中黄酮含量高达55.61%,经苯酚硫酸法[8]测定,AOEE中多糖含量为14.59%。
1.2 试验设计与饲粮本试验采用单因子随机区组设计,将240只肉仔鸡按照体质量相近原则随机分成5 个日粮处理组,每组6 个重复,每个重复8只鸡(雌雄各半),试验期为42 d。对照组饲喂基础饲粮,试验组分别在基础饲粮中添加250,500,750,1 000 mg/kg的AOEE。参照NY/T(33-2004)肉鸡营养需要量设计玉米-豆粕型基础饲粮。基础饲粮组成与营养水平值见表1。
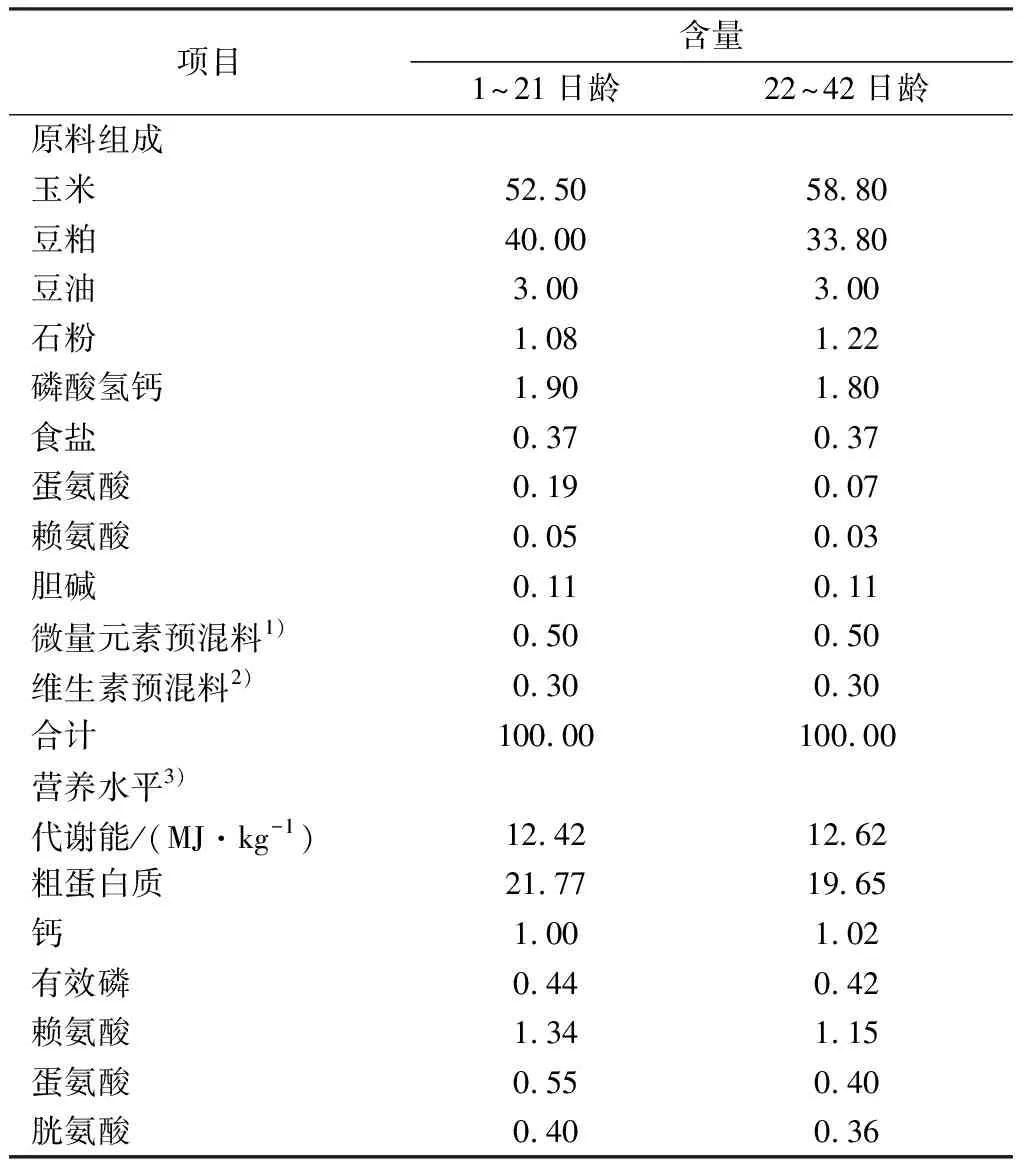
表1 基础饲粮组成及营养水平(风干基础) %
1.3 饲养管理试验期间肉鸡均采用单层笼养方式,每笼为1个重复(8只鸡),鸡只自由采食和饮水。试验1~3 d每天保持23 h光照,4~21 d每天保持10 h光照,22~42 d每天保持23 h光照。试验前对鸡舍环境、鸡笼和器具等进行彻底消毒。采用自然通风方式保持舍内空气清洁,鸡笼统一采用红外线加热装置的自动控温系统,试验前3 d保持舍内温度为(33±1)℃,之后每周降低3℃,直到降到21℃后保持恒定,试验期间每天人工清粪1次并保证料槽饲料和饮水充足。尽量保证各组的试验条件一致,并且每日观察并记录鸡的精神状态,及时剔除病鸡。
1.4 样品采集与制备分别于试验21与42 d屠宰分离脾脏、胸腺、法氏囊并称质量,采集肝脏、脾脏各2份,一份置于锡箔纸中,-20℃保存用于免疫指标的测定,另一份置于冻存管中,-80℃保存用于基因表达的测定。
1.5 测定指标及方法
1.5.1生长性能 分别于试验1,21,42 d早晨空腹称质量,以重复为单位计算试验鸡平均体质量(ABW)、体增重(BWG)并准确记录采食量和剩料量,计算试验鸡在试验前期、试验后期以及试验全期的采食量(FI)和饲料转化率(FCR),FCR=BWG/FI。
1.5.2免疫指标 于试验21与42 d屠宰后分离鸡的完整脾脏、胸腺和法氏囊,称质量并记录,计算免疫器官指数。免疫器官指数(g/kg)=免疫器官鲜质量(g)/活体质量(kg)。
称取0.5 g肝脏、脾脏组织样分别放入10 mL离心管内,按l∶9比例加入4.5 mL预冷生理盐水进行匀浆,充分匀浆后用4℃离心机3 000 r/min离心15 min,离心后取上清液。采用考马斯亮蓝法测定匀浆液上清的蛋白浓度,用ELISA试剂盒测定匀浆液上清中白细胞介素1β(IL-1β)、白细胞介素2(IL-2)、白细胞介素4(IL-4)、白细胞介素6(IL-6)、免疫球蛋白A(IgA)、免疫球蛋白G(IgG)和免疫球蛋白M(IgM)含量。
1.5.3免疫相关基因表达量 使用Trizol试剂盒提取肝脏、脾脏组织总RNA;用Hifair®Ⅱ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus)试剂盒将总RNA反转录为cDNA;使用Hieff®qPCR SYBR Green Master Mix试剂盒实时荧光定量PCR,步骤如下:95℃ 30 s;95℃ 5 s,60℃ 33 s,35个循环反应。95℃ 15 s;60℃ 60 s,95℃ 1 s,1次循环。所有样品做2个重复,并进行熔解曲线分析。以β-肌动蛋白(β-actin)为内参基因,采用2-ΔΔCt法[9]计算肝脏、脾脏免疫相关基因Toll样受体4(TLR4)、髓样分化因子88(MyD88)、核因子-κB p65(NF-κB p65)、NF-κB p50、IL-1β和IL-6的表达量。免疫相关基因引物序列如表2所示。

表2 免疫相关基因引物序
1.6 数据处理与统计分析试验数据采用SAS 9.2分析软件进行单因素方差分析(ANOVA),不同处理组的均值采用Duncan氏多重比较,采用回归分析探讨AOEE添加的剂量效应,P<0.01表示为差异或剂量效应极显著,P<0.05表示为差异或剂量效应显著,0.05≤P≤0.1为差异或剂量效应趋于显著。
2 结果
2.1 AOEE对肉仔鸡生长性能的影响如表3所示,在试验21 d,各试验组肉仔鸡的平均体质量无显著性差异(P>0.05),但有一次线性升高趋势;在试验42 d,随着AOEE添加量的增加,肉仔鸡的平均体质量、体增重呈极显著的一次线性或二次曲线升高效应(P<0.01),且与对照组相比,饲粮中添加750和1 000 mg/kg AOEE对肉仔鸡增重效果显著(P<0.05);各试验组肉仔鸡的采食量在前期和后期均无显著性差异(P>0.05);随着AOEE添加量的增加,试验前期的饲料转化率呈显著的二次曲线升高效应(P<0.05),试验后期的饲料转化率呈显著的一次线性升高效应(P<0.05),试验全期的饲料转化率呈显著的一次线性升高效应(P<0.05),且有二次曲线升高趋势。
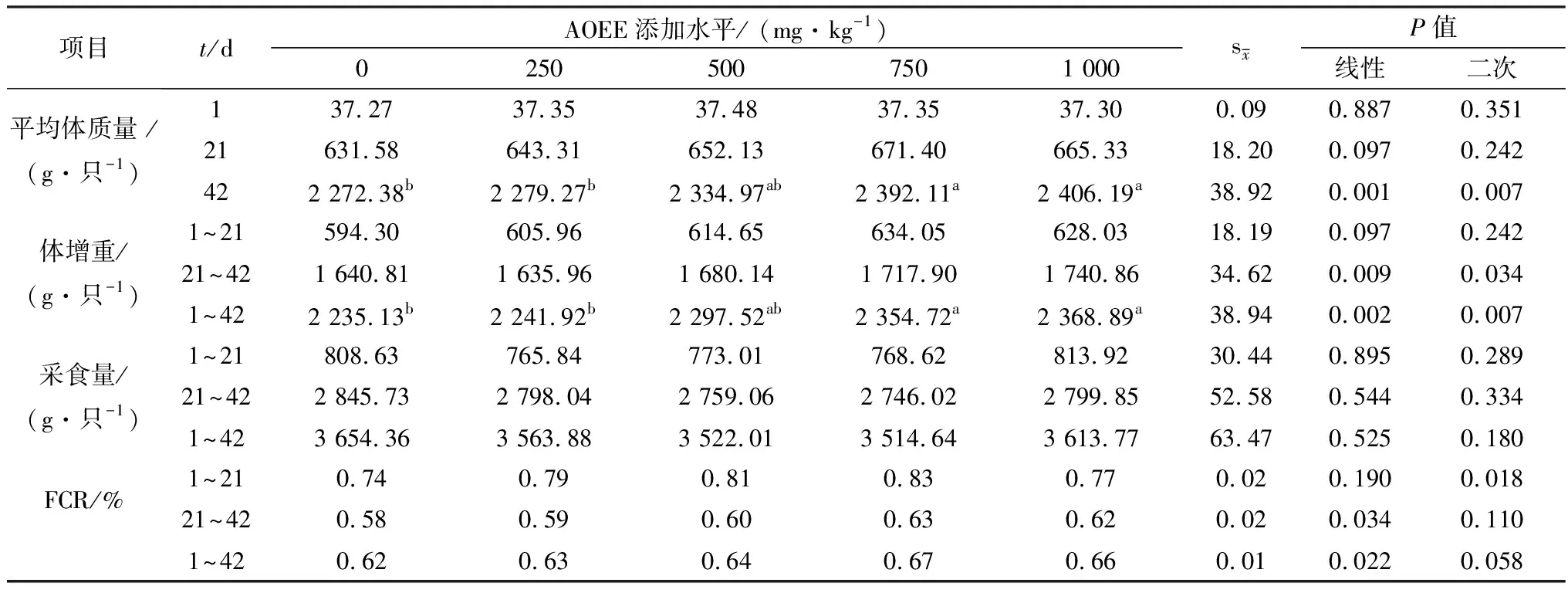
表3 AOEE对肉仔鸡生长性能的影响
2.2 AOEE对免疫器官指数的影响由表4可知,在试验期21 d,随着AOEE添加量的增加,各试验组肉仔鸡的胸腺指数有一次线性上升趋势,法氏囊指数有一次线性或二次曲线上升趋势;在试验期42 d,随着AOEE添加量的增加,肉仔鸡脾脏指数呈显著的一次线性升高效应(P<0.05),有二次曲线上升趋势,胸腺指数呈显著的一次线性或二次曲线升高效应(P<0.05)。
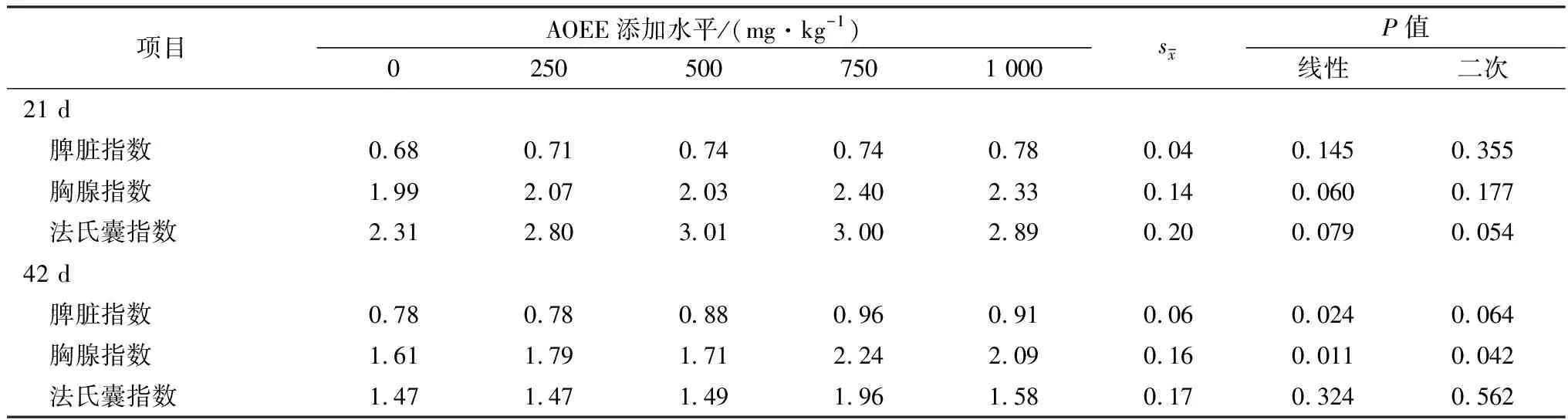
表4 AOEE对肉仔鸡免疫器官指数的影响 g/kg
2.3 AOEE对肉仔鸡肝脏、脾脏免疫指标及相关基因表达的影响从图1中可以看出,在试验期21 d,随着AOEE添加量的增加,肉仔鸡肝脏中IgA含量呈显著的一次线性或二次曲线升高效应(P<0.05),IgM含量呈极显著的二次曲线升高效应(P<0.01),IL-1β含量呈显著的二次曲线降低效应(P<0.05),IL-6呈极显著的一次线性或二次曲线降低效应(P<0.01);方差分析结果表明,与对照组相比,饲粮中添加750 mg/kg AOEE显著提高了肝脏中IgM的含量(P<0.05),并显著降低了IL-1β和IL-6的含量(P<0.05);在试验期42 d,随着AOEE添加量的增加,肉仔鸡肝脏中IgM含量呈极显著的二次曲线升高效应(P<0.01),IL-2含量呈极显著的一次线性或二次曲线升高效应(P<0.01),IL-4含量有一次线性升高效应(P<0.05),IL-1β含量有一次线性降低趋势;与对照组相较,饲粮中添加500 mg/kg AOEE显著提高了IgM的含量(P<0.05),饲粮中添加500,750,1 000 mg/kg AOEE显著提高了IL-2的含量(P<0.05)。
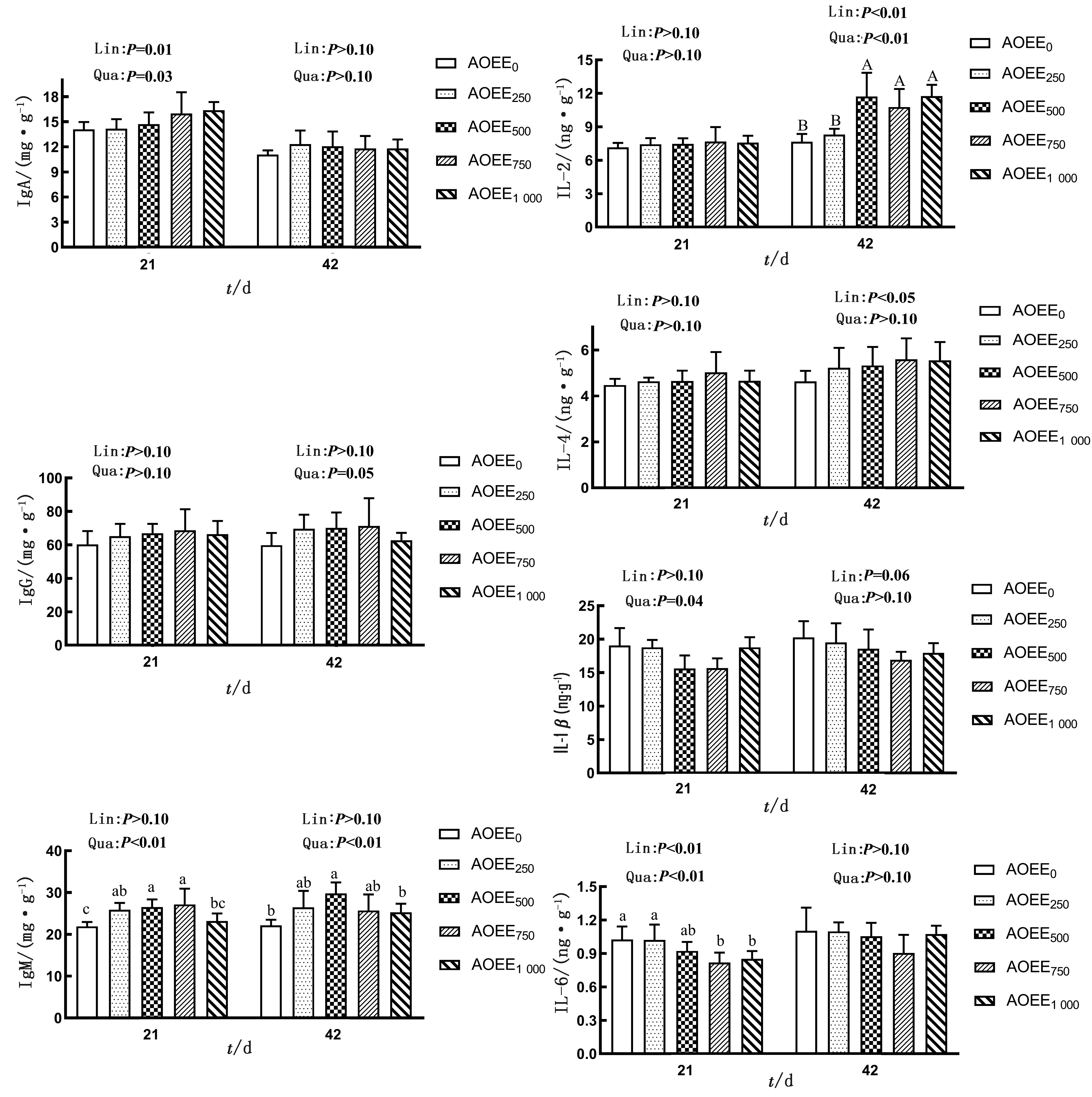
注:Lin.为一次线性;Qua.为二次曲线。柱状图上方无肩标字母或含相同字母表示单因素方差分析差异不显著(P>0.05);不同小写字母表示单因素方差分析差异显著(P<0.05);不同大写字母表示单因素方差分析差异极显著(P<0.01)。下同
如图2所示,在试验期21 d,随着AOEE添加量的增加,肉仔鸡肝脏中TLR4表达量有一次线性降低趋势,MyD88表达量呈显著的二次曲线降低效应(P<0.05),NF-κB p65表达量呈显著的一次线性降低效应(P<0.05),有二次曲线降低趋势,IL-6表达量呈极显著的一次线性或二次曲线降低效应(P<0.05),且方差分析结果表明,与对照组相比,饲粮中添加不同剂量的AOEE,均可下调肉仔鸡肝脏中IL-6表达量,且效果极显著(P<0.01);在试验期42 d,随着AOEE添加量的增加,TLR4表达量呈显著的一次线性或二次曲线降低效应(P<0.05),MyD88和NF-κB p50表达量呈极显著的一次线性或二次曲线降低效应(P<0.01),NF-κB p65表达量有二次曲线降低趋势,IL-6表达量呈极显著的一次线性(P<0.01)或显著的二次曲线(P<0.05)降低效应,方差分析结果表明,与对照组相比,饲粮中添加750和1 000 mg/kg AOEE极显著下调了MyD88表达水平(P<0.01),显著下调了NF-κB p50和IL-6表达量(P<0.05)。
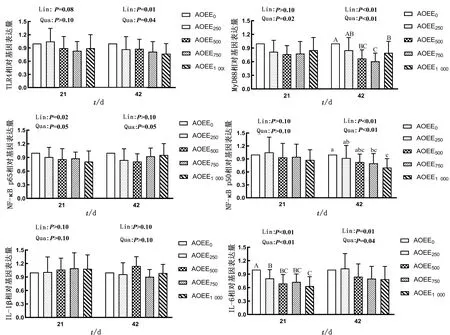
图2 AOEE对肉仔鸡肝脏免疫相关基因表达的影响
从图3中可以看出,在试验期21 d,随着AOEE添加量的增加,肉仔鸡脾脏中 IgA含量有一次线性或二次曲线升高趋势,IgG和IgM含量呈极显著的一次线性或二次曲线升高效应(P<0.01),IL-6含量有一次线性降低趋势,且方差分析结果表明,饲粮中添加500~1 000 mg/kg AOEE均可提高肉仔鸡脾脏中IgG和IgM的含量,且效果极显著(P<0.01);在试验期42 d,随着AOEE添加量的增加,肉仔鸡脾脏中IgA和IL-2含量呈极显著的一次线性(P<0.01)或显著的二次曲线(P<0.05)升高效应,IL-4含量呈显著的一次线性或二次曲线升高效应(P<0.05),IL-6含量呈显著的二次曲线降低效应(P<0.05),有一次线性降低趋势。

图3 AOEE对肉仔鸡脾脏免疫指标的影响
由图4可知,在试验期21 d,随着AOEE添加量的增加,肉仔鸡脾脏中TLR4 和IL-1β表达量有二次曲线降低趋势(P<0.05);在试验期42 d,随着AOEE添加量的增加,肉仔鸡脾脏中TLR4表达量有一次线性降低趋势,NF-κB p65表达量呈显著的一次线性降低效应(P<0.05),有二次曲线降低趋势。

图4 AOEE对肉仔鸡脾脏免疫相关基因表达的影响
3 讨论
3.1 AOEE对肉仔鸡生长性能的影响已有研究表明,蒿属植物富含黄酮类、有机酸以及微量化学元素,作为饲料添加剂能够显著提高动物新陈代谢,进而发挥促生长作用[10]。饲粮中添加黑沙蒿水提物可提高肉仔鸡日增重,且随着添加剂量的增加呈现出显著的一次线性或极显著的二次曲线升高效应[11]。此外,张晶等[12]报道,艾蒿醇提物可以显著提高肉仔鸡的末体质量,改善动物的生长性能,进一步研究发现,饲粮中添加艾蒿醇提物可显著缓解免疫应激下肉鸡的平均日增重的下降[13]。本试验结果表明,在饲粮中添加750 mg/kg AOEE可显著提高肉仔鸡的末体质量和增重,随着AOEE添加量的增加,肉仔鸡饲养后期以及全期的体增重和饲粮转化率呈显著的一次线性或二次曲线升高效应,说明适宜剂量的AOEE具有促进肉仔鸡生长的作用。经分析AOEE中的大部分活性物质为黄酮类化合物,而黄酮是一类具有抗菌活性的植物化学物质,它含有大量的羟基,酚羟基的氢键能够与碱性氨基酸侧链的氮原子结合,进入大肠杆菌内,导致细菌死亡,还可以对抗消化道中的致病菌和其他有害细菌,改善肠道菌群平衡,促进肠道消化吸收,从而提高肉鸡的饲料转化率[14],因此推断AOEE的促生长效果可能是黑沙蒿黄酮发挥作用。
3.2 AOEE对肉仔鸡免疫器官指数的影响禽类的免疫器官主要是脾脏、胸腺和法氏囊。脾脏是禽类最大的外周免疫器官,富含吞噬病菌的免疫细胞,并产生免疫球蛋白,是机体细胞免疫和体液免疫的中心[15];胸腺是细胞免疫的中枢器官,也是T淋巴细胞分化成熟的场所,与细胞免疫和体液免疫密切相关,在维持机体免疫稳定方面起重要作用[16];法氏囊又称腔上囊,是禽类特有的中枢免疫器官,是B淋巴细胞分化、发育、成熟以及免疫球蛋白基因分化和基础基因库形成及扩展的主要器官,对维持机体体液免疫功能十分重要[17]。因此,脾脏、胸腺和法氏囊的大小可反映禽类的免疫状况,免疫器官相对质量增加,预示着机体免疫机能增强[18]。本试验结果表明,试验期内肉仔鸡的脾脏指数和胸腺指数随AOEE添加量的增加呈现出显著的一次线性或二次曲线升高效应,对法氏囊发育也有一定的促进作用,这与先前的研究柑橘乙醇提取物以剂量依赖方式提高肉仔鸡免疫器官指数结果相符[19]。
3.3 AOEE对肉仔鸡肝脏、脾脏免疫指标的影响在畜禽饲养的过程中,由于各种外界因素会引起机体产生应激,损伤机体免疫功能。免疫球蛋白作为体液免疫的重要组成部分,不仅能够促进单核细胞和巨噬细胞的吞噬作用,还可以与抗原结合产生多种生物学效应,在机体的特异性免疫应答中发挥十分重要的作用[20]。肝脏作为机体最大的网状内皮细胞吞噬系统,在防御毒性物质侵袭、防止病原体感染中发挥重要作用[21]。本试验结果表明,与对照组相比,饲粮中添加500~1 000 mg/kg AOEE显著提高肉仔鸡肝脏中IgM的含量,极显著提高脾脏中IgG及IgM的含量,且呈剂量依赖性地随饲粮中AOEE添加量的增加而升高,这说明适宜剂量的AOEE可以促进肉仔鸡体液免疫。细胞因子是细胞间信息传递的信使,是反映机体免疫应答与炎症调控的重要指标[22]。门连超等[23]在研究藏药兔耳草醇提物对小鼠免疫功能的影响时发现,藏药兔耳草醇提物可显著提高小鼠机体内IL-2和IL-4的水平,在本研究中,AOEE提高了肝脏和脾脏中抗炎因子IL-2和IL-4的分泌,降低了促炎因子IL-1β和IL-6的水平,因此可以确定,适宜剂量的AOEE可以通过调节肉仔鸡免疫球蛋白和细胞因子的水平来调节机体免疫功能。这可能与黄酮类化合物能够增强巨噬细胞吞噬能力,通过调节先天性和适应性免疫系统中免疫细胞因子和免疫球蛋白的产生来减轻炎症,还能够通过与细胞因子受体的结合起到抗炎作用,从而增强机体免疫能力有关[24]。
3.4 AOEE对肉仔鸡肝脏、脾脏免疫相关基因表达量的影响TLR4是细胞膜上的一种模式识别受体,可对不同病原相关分子模式进行识别、结合,并传递信号给下游的炎症信号转导因子NF-κB[25]。NF-κB是细胞核内调节免疫基因表达的关键转录因子,主要由p65和p50结合成二聚体保持沉默状态[26]。当TLR4受到细胞外信号刺激时,通过信号转导衔接蛋白MyD88激活NF-κB,进而促进末端炎症因子肿瘤坏死因子α(TNF-α)、IL-1和IL-6等的转录释放,诱发免疫反应[27]。有研究报道,滨蒿总黄酮可作用于信号通路中病毒感染相关靶点,使TLR3、NF-κB mRNA表达水平明显下调,同时也显著降低IL-6、TNF-α和iNOS mRNA表达水平,由此认为滨蒿总黄酮可通过抑制TLR3/NF-κB信号通路的作用和机体炎症反应来发挥免疫调节和抗炎的作用[28]。李昆[29]研究发现,注射LPS后显著上调了肉鸡各组织中TLR4、MyD88和NF-κB的表达量,说明免疫应激导致了TLR4的过度表达,MyD88、NF-κB的表达量也显著上升,然而在饲粮中添加黑沙蒿提取物可以不同程度地降低上述基因的表达水平。在本试验中,与对照组相比,饲粮中添加AOEE可显著降低MyD88、NF-κB/p50的mRNA的表达,并显著下调了炎症因子IL-6的mRNA表达水平,且随着AOEE添加量的增加,肉仔鸡肝脏中TLR4受到抑制,其下游信号分子MyD88、NF-κB/p65、NF-κB/p50的转录水平均呈现显著的一次线性或二次曲线降低效应,从而调控了IL-6的表达量,在试验后期尤为显著。这可能由于黄酮类化合物在胞外可与细胞表面的TLR4相互作用,抑制下游通路NF-κB的激活[30],也有可能是黄酮类化合物进入细胞直接抑制IKK(NF-κB激酶)的活性,下调NF-κB的激活,从而降低肉仔鸡免疫器官的炎症反应[31]。因此可以推断,AOEE可能通过黄酮类化合物抑制TLR4/NF-κB信号通路来调节机体免疫。
猜你喜欢
福建中学数学(2023年5期)2024-01-25 17:41:36
养殖与饲料(2021年5期)2021-11-30 20:05:15
现代畜牧科技(2021年10期)2021-11-19 08:42:20
草食家畜(2021年3期)2021-06-08 03:18:16
中国猪业(2019年7期)2019-11-16 07:15:28
兽医导刊(2019年20期)2019-08-19 10:39:18
云南教育·中学教师(2019年12期)2019-08-13 07:28:20
今日畜牧兽医(2018年9期)2018-02-13 09:05:24
数学学习与研究(2016年19期)2016-11-22 11:39:30
明清小说研究(2015年3期)2015-11-30 01:26:03
