
东亚孑遗植物柳杉属的遗传分化及其种群进化历史*
2022-10-16 07:47李鑫玉王敏求袁美灵SaneyoshiUeno武星彤蔡梦颖YoshihikoTsumura文亚峰
林业科学 2022年6期
李鑫玉 王敏求 袁美灵 Saneyoshi Ueno 武星彤 蔡梦颖, Yoshihiko Tsumura 文亚峰
(1.中南林业科技大学风景园林学院 长沙 410004; 2.日本森林综合研究所森林分子遗传学与生物技术部 筑波 305-8687;3.筑波大学生命与环境科学学院 筑波 305-8572)
植物现代地理分布和遗传变异格局是多种生物与非生物因素协同作用的结果 (Harrisonetal., 2001; Wangetal., 2013),其中,地理和环境气候变化是最关键的驱动因素,会对物种形成、遗传分化、种群进化产生显著影响 (Hewitt, 1996; 2004; Begonetal., 2006)。地理因素主要包括山脉隆起、陆地变迁、海水退却等地质事件,而全球气候变化则以第四纪冰期-间冰期的循环最为剧烈,其对植物物种形成和现代地理分布所造成的影响极其深刻 (Bennett, 1997)。
现代植物类群中,第三纪孑遗植物以其起源古老、形态特殊、进化缓慢吸引了众多研究者的关注(Tiffney, 1985, Tiffneyetal., 2001; Milneetal., 2002),被称为植物“活化石”。晚第三纪和第四纪期间,受全球气候变化的影响,这些曾在第三纪广泛分布的植物逐渐退缩至东亚、北美和欧洲西南部三大冰期避难所 (Milneetal., 2002)。这些区域中,东亚植物区系是全球植物多样性最为丰富和复杂的地区,拥有全球最古老和最完整的植物多样性演化系列。同时,现代东亚植物区系反映了重大的古地质和环境气候变化 (邱英雄等,2017),是研究北半球植物起源和进化的关键地区。杉科 (Taxodiaceae) (现归属于柏科)是第三纪孑遗植物中独特的裸子植物类群,多为单型属或寡型属。著名的活化石植物水杉在该区域的发现,也使其成为中国-日本植物区系的谱系代表 (Chenetal., 2018)。鉴于杉科植物悠久的演化历史,独特的生活习性,及其在东亚地区的广泛分布,研究它的进化历史和地史分布对于揭示整个松杉类植物的系统演化、地理分布及全球古植物地理区系和古气候变化均有重要意义。
柳杉属(Cryptomeria)树种为东亚特有,是现存杉科植物中最原始的类群,祖先的大多数原始性状在该属得到保留 (于永福,1995)。柳杉属间断分布于我国和日本,长期的人工栽培与广泛利用,使其起源、天然分布地和分类都存在很大争议。有的认为日本和我国东部的天目山、武夷山是该属的起源地和天然种群分布中心 (王江等,2007; 袁美灵等,2019),但也有认为柳杉原产日本,柳杉在我国属外来物种 (Chenetal., 2008)。柳杉属的分类存在1个种和2个种的争议,较多学者认为柳杉属是单种属,包括柳杉(Cryptomeriajaponicavar.sinensis)和日本柳杉(Cryptomeriajaponicavar.japonica)两个变种 (Hayashi, 1960; Fuetal., 1999),也有认为长期的地理隔离和种群分化,使柳杉和日本柳杉产生了可遗传的表型变异,其应属于2个独立的种(Cryptomeriafortunei和Cryptomeriajaponica) (王江等,2007)。
日本柳杉作为日本的“国树”,不仅占据了44 %的人工林资源,而且其天然林分布广泛 (Tsumuraetal.,2011),从北部的青森县至南部的屋久岛都有较大面积分布。迄今为止,柳杉属的遗传学和谱系地理学研究多以日本柳杉为主。早期的表型分析和现代分子遗传学研究表明,日本柳杉天然种群间存在显著的遗传分化 (Kojo, 1987; Tsumuraetal., 2007;2012; 2014; Kimuraetal., 2013),可分为Omote-sugi和Ura-sugi2个地理变种,分别分布于日本岛的太平洋沿岸和日本海沿岸。日本柳杉的种群进化历史也广受关注,花粉化石资料显示,170万年前的气候变化使日本柳杉产生了适应性特征,种群分化可能发生在30万年前。Kimura等(2014)基于SSR数据和贝叶斯计算方法,认为日本柳杉不同地理谱系的分化时间为8万年前。利用核基因组序列数据,Moriguchi等(2019)得到的日本柳杉分化时间为30万年前。在化石资料不足的情况下,估算物种的分化时间,既与不同的标记方法有关,也与遗传分化模型和基因突变率的选择有直接关系。虽然以上2个研究的分化时间略有差异,但共同显示日本柳杉谱系分化发生在末次盛冰期(LGM)之前的较近时间,类似于“活化石”植物银杏(Ginkgobiloba),中国现存银杏种群的遗传足迹仅可追溯至39万年前 (Hohmannetal., 2018)。最近的研究也表明,第四纪冰期气候变化,特别是末次盛冰期对东亚地区柳杉的遗传多样性和遗传分化产生了深刻影响 (Tsumuraetal., 2020),但该研究只用了1个柳杉天然种群,对于分析柳杉和日本柳杉的种群进化历史尚有不足。因此,东亚全域范围内的柳杉种群研究,特别是柳杉和日本柳杉的遗传分化和种群历史动态有待进一步深入。
基于此,本研究以东亚全域分布柳杉属树种的天然种群为研究材料,利用核基因组微卫星标记技术,解析该属的遗传变异格局,以期掌握其遗传多样性水平和遗传结构,阐明其种群进化历史。在此基础上,分析地理和气候变化在柳杉遗传分化中的作用,进一步探讨柳杉属的起源和分类等科学问题。
1 材料与方法
1.1 研究材料
根据我国柳杉资源分布现状,选取浙江、福建和安徽等省区的柳杉天然林(或古树林)种群为研究材料。2017—2018年采集到10个种群共189个单株(胸径大于100 cm)样本(表1、图1),取嫩叶适量,利用改良CTAB法提取基因组DNA (张景丽,2014),符合质量要求的DNA稀释至20~30 ng·μL-1用于试验分析。日本柳杉样本来源于日本青森县、秋田县和新泻县等8个天然种群的109个单株,其DNA由日本森林综合研究所提供(表1、图1)。
1.2 基因分型
从日本柳杉核基因微卫星(nSSR)中筛选14个多态性丰富的位点用于基因分型 (Tanietal., 2003; Moriguchietal., 2003; Moriguchietal., 2014) (表2),这些位点经检测分析均不含无效等位基因。正向引物5′端加荧光接头FAM,HEX或TAMRA。PCR扩增体系为15 μL,含2 × Taq PCR MasterMix (Tiangen) 7.5 μL,正向引物(F)和反向引物(R)各0.75 μL(10 μmol·L-1),DNA模板3 μL,ddH2O 3 μL。PCR反应程序为: 94 ℃预变性4 min; 94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸45 s,共35个循环; 72 ℃延伸10 min。1.6 %琼脂糖凝胶电泳检测PCR扩增产物,利用ABI3730xL分析仪进行基因分型(Liz 500为内标),GeneMarker version 2.20软件 (Hulceetal., 2011)统计基因分型结果。
1.3 数据分析
1.3.1 遗传多样性与遗传分化 利用GenAlEx6.5 (Peakalletal., 2012)统计等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)和基因流(Nm)等遗传参数。HP-Rare 1.1 (Kalinowski,2005)计算各种群等位基因丰富度(Ar)和私有等位基因丰富度(PAr)。FSTAT (Goudet,1995)用于估算近交系数(FIS)、总的遗传多样性(Ht)、种群内遗传多样性(Hs)和遗传分化系数(GST和FST)。基因流根据公式Nm= (1/FST-1)/4计算获得。
1.3.2 遗传结构 根据地理来源将东亚地区的柳杉属植物样本分为柳杉和日本柳杉2个组群,利用Arlequin 3.0 (Excoffieretal.,2005) 按组群间、种群间和种群内进行遗传变异分子方差分析(AMOVA)。STRUCTURE 2.3.3 (Pritchardetal.,2000) 用于遗传结构分析,在混合模型 (Admixture model)下,设置Burn in period和MCMC (Markov Chain Monte Carlo) 的迭代参数分别为50 000和100 000,K值设定为1~18,每个K值运算10次,其余使用默认设置(Q值为0.9)。根据STRUCTURE HARVESTER (Earletal.,2012)计算ΔK数值的分布,确定最优K值。利用CLUMPP软件 (Jakobssonetal., 2007) 计算对应K值的最优Q-matrix结果,并利用Distruct软件 (Rosenberg, 2004) 输出可视化图形。基于种群间FST矩阵,利用MEGA 7软件(Kumaretal.,2016)构建系统发育树(邻接法neighbor-joining, NJ)。基于R语言平台,执行‘a.score’主成分判别分析(DAPC) (Jombart, 2008)。用GenAlEx6.5软件(Peakalletal., 2012)进行Mantel检验,检测地理距离和遗传距离的相关性,设置999次随机置换。
1.3.3 种群历史动态 利用DIY ABC软件 (Cornuetetal.,2014)基于近似贝叶斯算法,模拟分析柳杉属种群的历史动态。95%置信水平下,采用Logistic回归,使用最接近模拟数据的1%观测数据,估算每个进化事件的后验概率(posterior probability, PP),后验概率最大的进化事件为最优进化事件。根据STRUCTURE、主成分判别分析(DAPC)和NJ聚类分析结果,按不同的基因库分别构建东亚地区柳杉的种群进化事件。

表1 柳杉属种群地理位置及样本数量Tab.1 Locality information and sample size of Cryptomeria populations
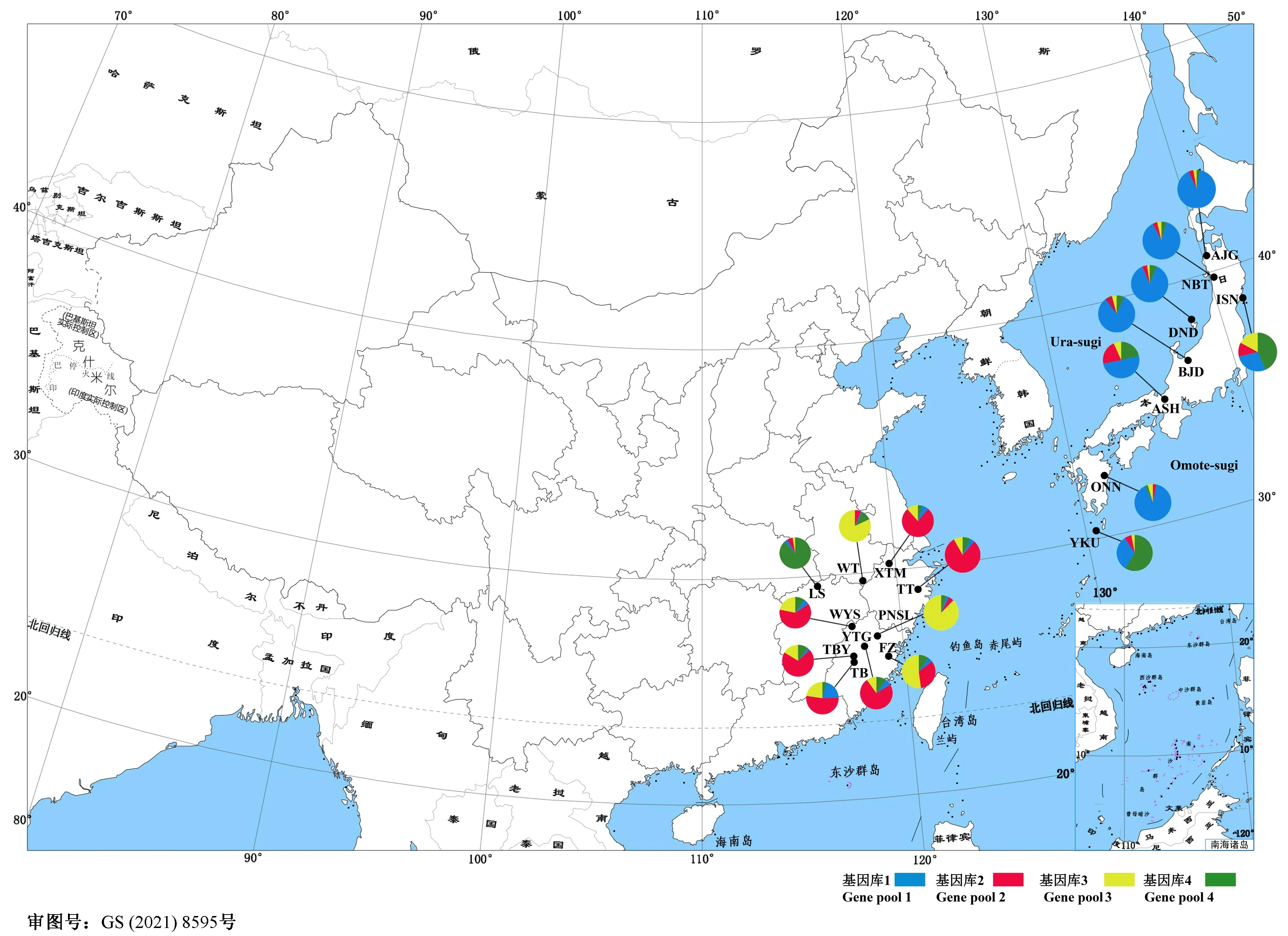
图1 柳杉属种群地理分布及基于CLUMPP结果的基因库分布图(K = 4)Fig. 1 Geographic distribution of Cryptomeria populations and gene pool distribution map of each population based on CLUMPP results (K = 4)
(1)K=2时,分为柳杉(CHN)和日本柳杉(JAN)2个基因库,构建3种进化事件:
事件1: 柳杉(CHN)和日本柳杉(JAN)由同一祖先种群分化而来(t1);
事件2: 日本柳杉(JAN)在t1时间由柳杉(CHN)分化而来;
事件3: 柳杉(CHN)在t1时间由日本柳杉(JAN)分化而来。
(2)K=4时,分为中国东南部种群(CHS)、庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi) 4个基因库,构建4种进化事件:
事件1: 日本海沿岸种群(Ura-sugi)于t1时分化出太平洋沿岸种群(Omote-sugi),随后t2、t3时间依次分化出庐山种群(LS)和中国东南部种群(CHS);
事件2: 中国东南部种群(CHS)和日本海沿岸种群(Ura-sugi)由同一祖先种群分化而来(t1),随后,二者于t2时间发生混合事件,产生太平洋沿岸种群(Omote-sugi),t3时间,太平洋沿岸种群(Omote-sugi)与中国东南部种群(CHS)混合,分化出庐山种群(LS);
事件3: 中国东南部种群(CHS)、庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi)由同一祖先种群分化而来(t1);
事件4: 庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi)种群于t1时间从同一祖先种群分化而来,随后t2时间中国东南部种群(CHS)由庐山种群(LS)分化而来。
2 结果与分析
2.1 遗传多样性与遗传分化
14个nSSR位点共检测到219个等位基因,不同位点的等位基因数(Na)从2到33不等,平均为15.643。柳杉属总的遗传多样性(Ht)为0.570,种群内遗传多样性(Hs)为0.508 (表2)。不同种群的平均等位基因数(Na)在2.714~9.286之间,平均期望杂合度(He)和观测杂合度(Ho)分别为0.488和0.472。以最小种群样本数6为基数,庐山种群(LS)具有最高的等位基因丰富度和私有等位基因丰富度 (Ar = 5.271,(PAr = 0.976),天宝岩高海拔种群(TB)的等位基因丰富度最低(Ar = 2.453)。柳杉属种群遗传分化系数(FST)为0.136,基因流(Nm)为1.588 (表4),近交系数(FIS)从-0.100到0.294,平均值为0.060。在组群水平,柳杉的遗传多样性(Na= 4.571,Ho= 0.442)低于日本柳杉(Na= 4.634,Ho= 0.510) (表3),但柳杉的遗传分化程度(FST= 0.121)明显高于日本柳杉(FST= 0.028)。
2.2 遗传结构
根据AMOVA分析结果,柳杉属的遗传变异主要来自种群内的个体间,种群内遗传变异占总变异的86.37% (P< 0.001) (表4)。STRUCTURE分析表明,K= 2时出现明显拐点并取得最大值,说明柳杉属种群最有可能划分为2个基因库(gene pool) (图2a、b),分别对应柳杉(除LS种群)和日本柳杉。K= 3时,柳杉种群中出现新的基因库(黄色),日本柳杉种群未出现新的基因库。K= 4时,日本柳杉分为Ura-sugi和Omote-sugi两个明显的地理谱系,而中国庐山种群(LS)与Omote-sugi属同一基因库(绿色) (图2c)。NJ系统发育树与主成分判别分析(DAPC)结果显示柳杉属可分为中国东南部(CHS)、中国庐山(LS)、日本海沿岸(Ura-sugi)和太平洋沿岸(Omote-sugi)四个谱系(图3、4)。
Mantel检验结果显示,柳杉属种群的遗传距离和地理距离之间不具有显著相关性(R2=0.008 5,P=0.171) (图5a)。在组群的水平上,柳杉(排除庐山种群)中没有发现二者之间的显著相关性(R2=0.016 4,P=0.226) (图5b),但日本柳杉中则具有显著相关性(R2=0.223 5,P=0.018) (图5c)。

表2 14个nSSR位点的遗传多样性参数Tab.2 Genetic diversity parameters of 14 nSSR loci

表3 柳杉属种群的遗传多样性参数Tab.3 Genetic diversity parameters of the Cryptomeria populations
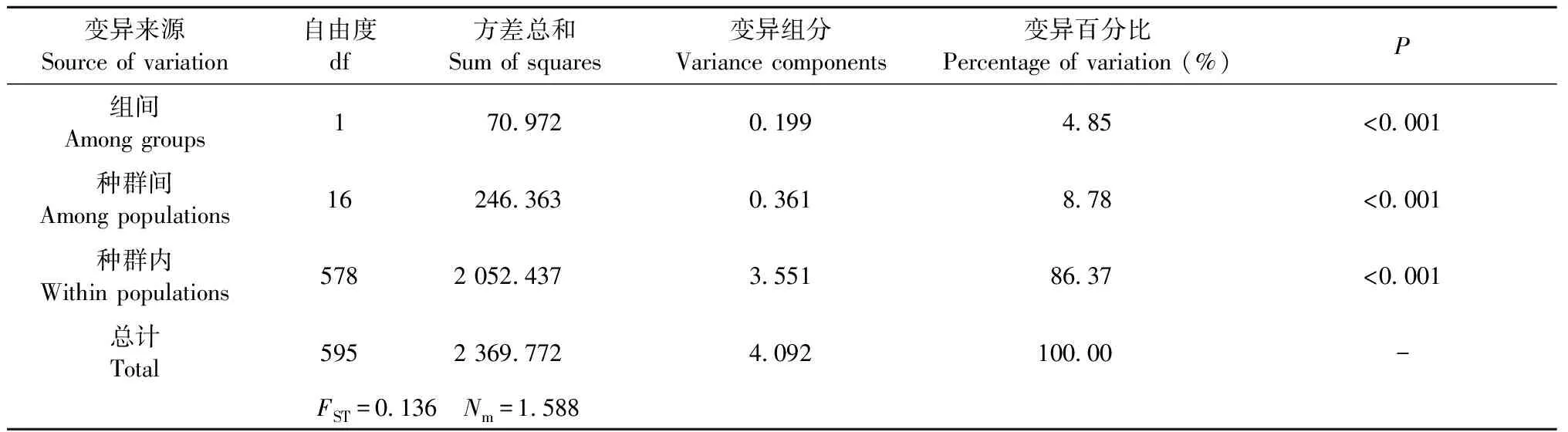
表4 柳杉属种群间分子方差分析(AMOVA)Tab.4 Analysis of molecular variance (AMOVA) of Cryptomeria populations
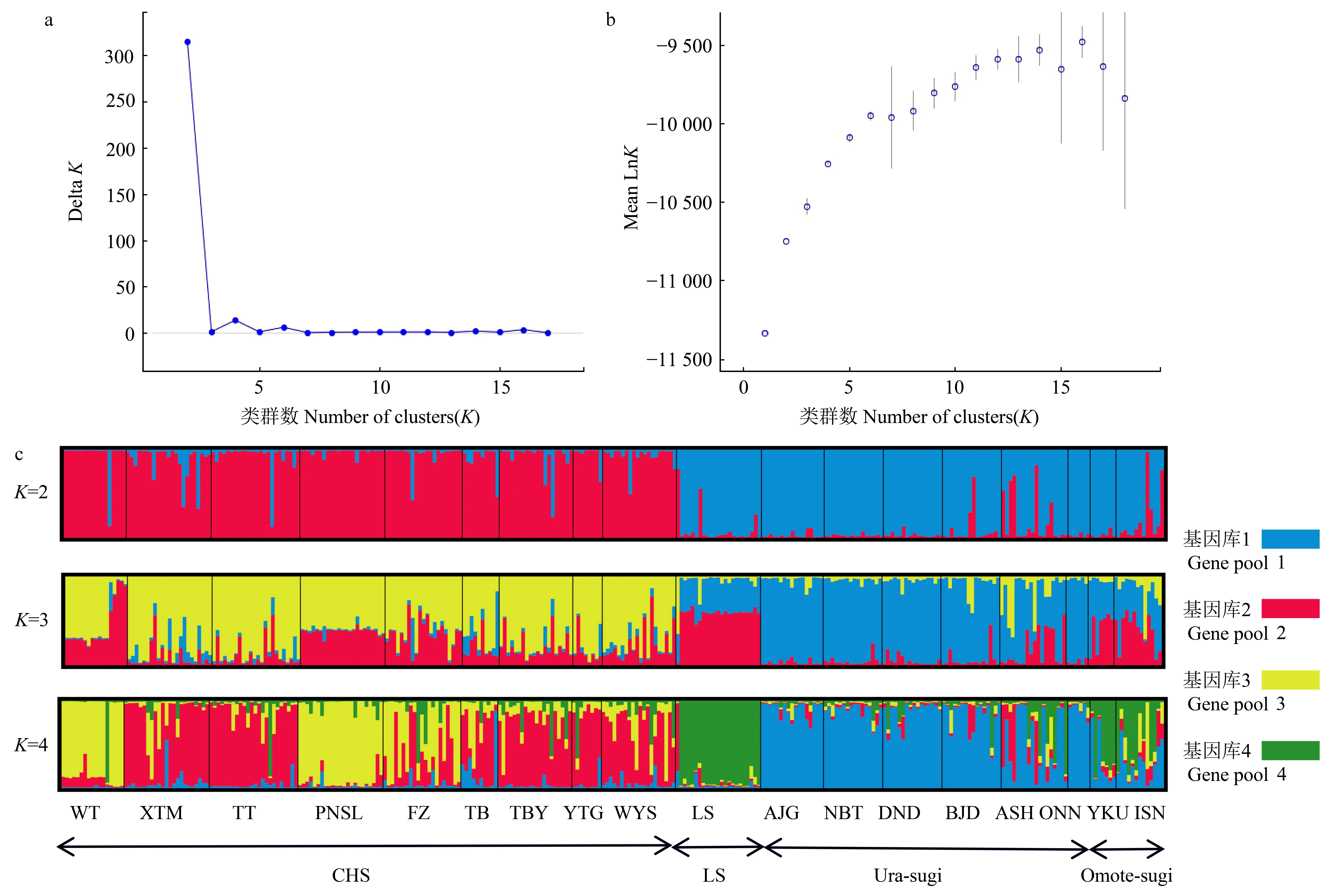
图2 柳杉属种群STRUCTURE分析结果Fig. 2 Results of STRUCTURE analysis of the Cryptomeria populationsa: K值与Delta K关系图; b: K值与Mean LnK关系图; c: 当K=2, 3, 4时, STRUCTURE分析结果的直方图。 CHS: 中国东南部种群; LS: 中国庐山种群; Ura-sugi: 日本海沿岸种群; Omote-sugi: 太平洋沿岸种群。a: Relationship between Delta K and K value; b: Relationship between Mean LnK and K value; c: Histogram of the STRUCTURE analysis for the model with K=2, K=3 and K=4;; CHS: the populations of Southeastern China; LS: the populations of Lushan China; Ura-sugi: the populations of the Japan Sea coast; Omote-sugi: the populations of the Pacific coast.
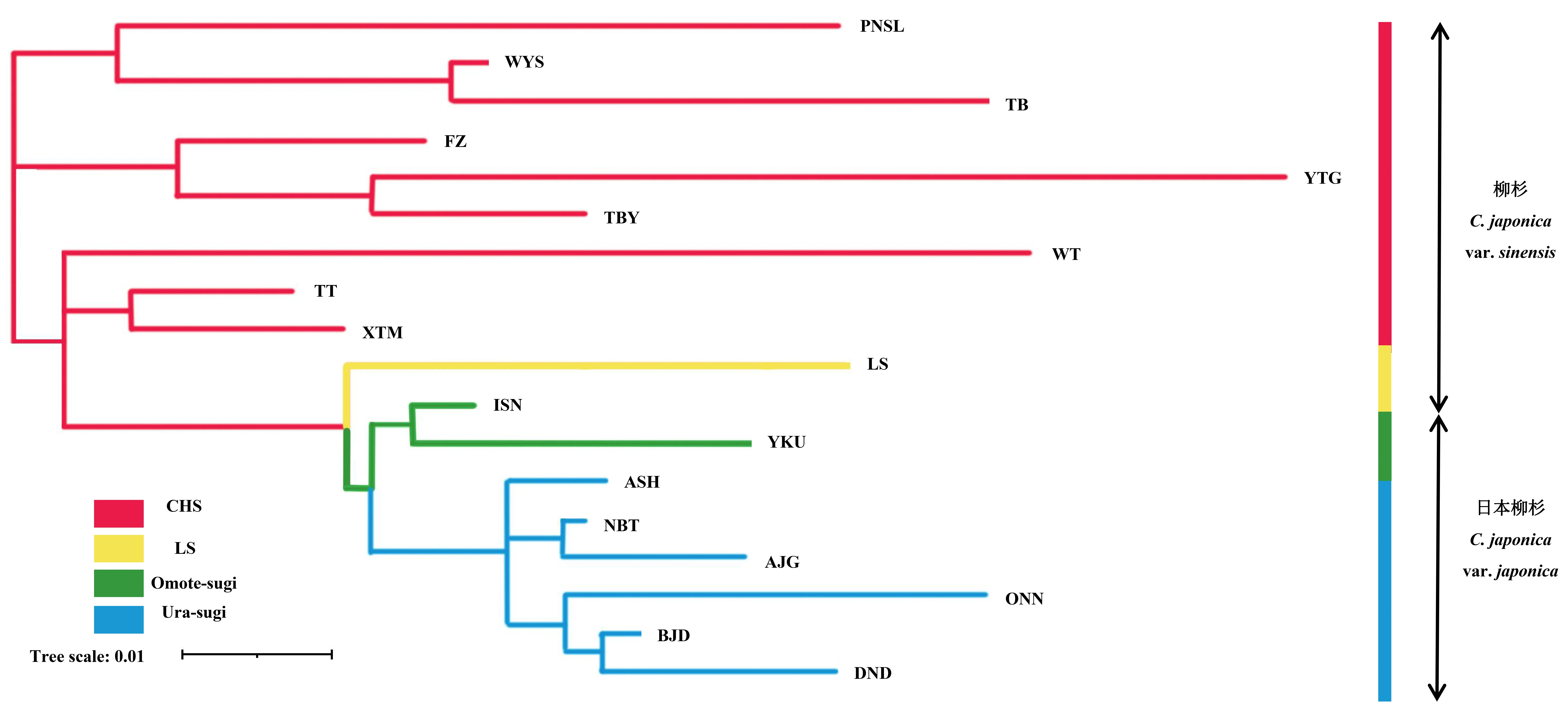
图3 基于成对FST矩阵的柳杉属种群系统发育树(NJ法)Fig. 3 Phylogenetic tree of the Cryptomeria populations based on pairwise FST values (NJ method)CHS: 中国东南部种群; LS: 中国庐山种群; Ura-sugi: 日本海沿岸种群; Omote-sugi: 太平洋沿岸种群。CHS: the populations of Southeastern China; LS: the populations of Lushan China; Ura-sugi: the populations of the Japan Sea coast; Omote-sugi: the populations of the Pacific coast.
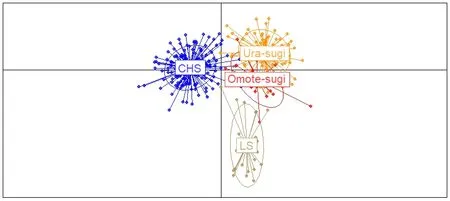
图4 柳杉属种群的主成分判别分析图(DAPC)Fig. 4 Principal component discriminant analysis (DAPC) of the Cryptomeria populationsCHS: 中国东南部种群; LS: 中国庐山种群; Ura-sugi: 日本海沿岸种群; Omote-sugi: 太平洋沿岸种群。CHS: the populations of Southeastern China; LS: the populations of Lushan China; Ura-sugi: the populations of the Japan Sea coast; Omote-sugi: the populations of the Pacific coast.
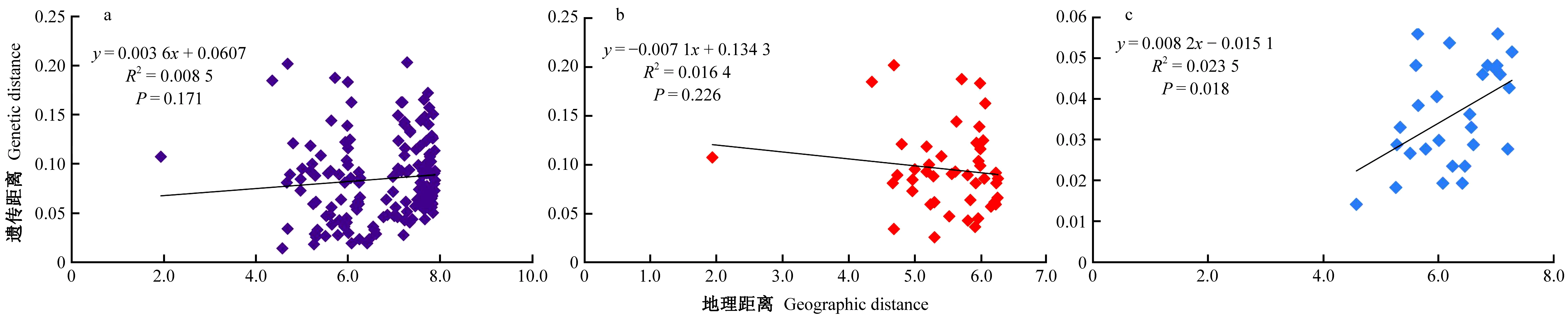
图5 柳杉属种群地理距离与遗传距离相关性检验Fig. 5 Geographical distance against genetic distance for the Cryptomeria populationsa: 柳杉属种群; b: 柳杉种群(除庐山LS); c: 日本柳杉种群。a: Cryptomeria populations; b: C. japonica var. sinensis populations (except LS); c: C. japonica var. japonica populations.
2.3 种群历史动态
为阐明柳杉属树种的种群历史动态,根据STRUCTURE与DAPC分析结果,按不同的基因库分2步对其种群历史进行模拟分析。当K=2时,3个进化事件的后验概率(PP)分别为0.994 9,0.005 0和0.000 1,事件1为最优事件,即柳杉和日本柳杉在t1(1 010代)时从同一祖先种群分化而来,其有效种群分别为N1= 3 880;N2= 9 240;NA= 92 (图6)。当K=4时,最优事件为3 (PP=0.991 9),即中国东南部种群(CHS)、中国庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi)由同一祖先种群分化而来,分化时间t1为1 140代,其有效种群分别为N1= 3 820;N2= 7 840;N3= 4 850;N4= 7 350;NA= 44.6 (图7、表5)。

图6 基于近似贝叶斯算法的柳杉属进化事件模型(K = 2)Fig. 6 The scenarios of Cryptomeria populations based on Approximate Bayesian Computation (K = 2)CHN: 柳杉种群; JAN: 日本柳杉种群; PP: 事件的后验概率; t1: 事件的分化代数; N1, N2, NA: 有效种群的大小。CHN: C. japonica var. sinensis; JAN: C. japonica var. japonica; PP: the posterior probability of scenario; t1: the generation of scenario; N1, N2, NA: the size of effective population.

图7 基于近似贝叶斯算法的柳杉进化事件模型(K = 4)Fig. 7 The scenarios of Cryptomeria populations based on Approximate Bayesian Computation (K = 4)CHS: 中国东南部种群; LS: 中国庐山种群; Ura-sugi: 日本海沿岸种群; Omote-sugi: 太平洋沿岸种群; t1, t2, t3: 事件的分化代数; N1, N2, N3, N4, NA: 有效种群的大小。CHS: the populations of Southeastern China; LS: the populations of Lushan China; Ura-sugi: the populations of the Japan Sea coast; Omote-sugi: the populations of the Pacific coast; t1, t2, t3: the generation of scenario; N1, N2, N3, N4, NA: the size of effective population.
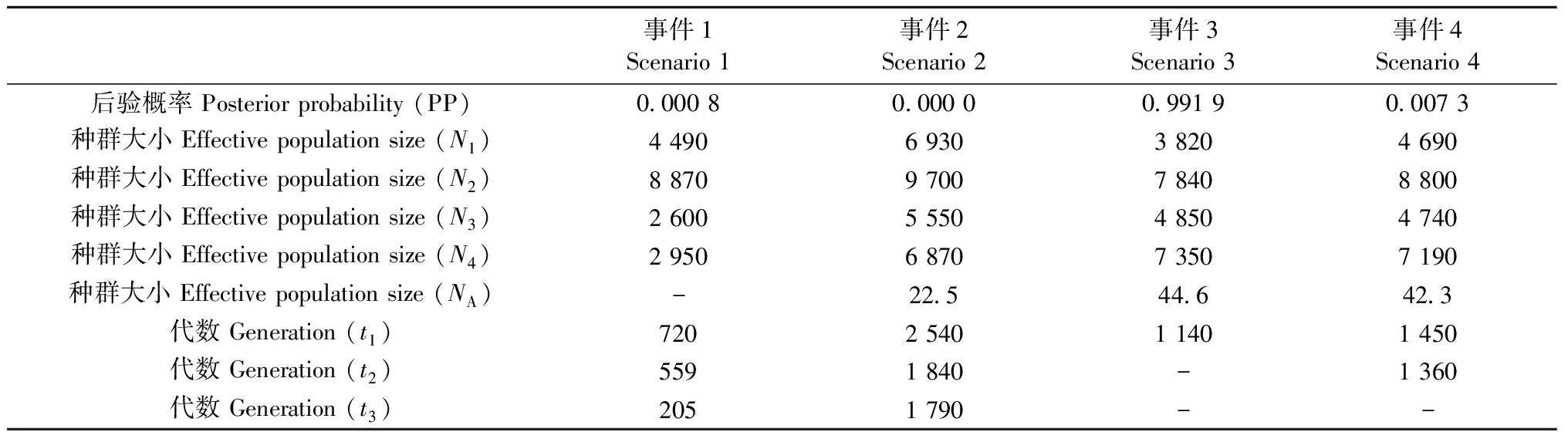
表5 基于DIY ABC的各进化事件的后验分布的中值估计(K = 4)Tab.5 Median estimation of posterior distributions for each scenario based on DIY ABC (K = 4)
3 讨论
3.1 种群遗传多样性与遗传分化
本研究表明,柳杉属树种的遗传多样性水平中等(Ho= 0.472,He= 0.488),其值低于杉木(Cunninghamialanceolata) (He= 0.687) (李霞等,2020) 和长苞冷杉(Abiesgeorgei) (He= 0.751) (程小毛等,2016),但高于水松(Glyptostrobuspensilis) (Ho= 0.317,He= 0.272) (Wuetal.,2019) 、台湾杉(Taiwaniacryptomerioides) (Ho= 0.231,He= 0.370) (李江伟,2014) 等杉科孑遗植物。种群间存在较高的遗传分化(FST= 0.136),可分为柳杉和日本柳杉2个地理种群。柳杉具有遗传多样性低、种群间遗传分化大、基因流小的特点(Na= 4.571,Ho= 0.442,FST= 0.121)。
植物遗传多样性会受繁育系统、遗传漂变、基因流和人为干扰等因素的影响 (文亚峰等,2010),近交繁殖、遗传漂变、基因流受限、种群规模变小都会导致遗传多样性的降低。和日本柳杉相比较,柳杉表现出明显的近交繁殖特性(FIS= 0.123),种群间基因流十分有限(Nm= 1.816),远低于日本柳杉的基因流指数(Nm= 8.679),更低于针叶树种基因流平均值(Nm> 3) (Ledig,1998)。考虑到86.37%的变异发生在种群内,我们认为有限的基因流以及由此所导致的近交繁殖是造成柳杉遗传多样性低的主要原因。根据Cai等(2020)的研究结果,柳杉的种群数量自中更新世以来呈持续下降的趋势,而全新世以来人类活动的增强使柳杉生境片断化日趋严重,这些因素进一步加剧了其遗传多样性的衰减。相比较而言,日本柳杉天然林分布广泛,且得到了较好地保护,人为干扰少,保持了较高的遗传多样性。日本岛中部的山地阻隔,太平洋和日本海两侧环境气候的差异,从而导致日本柳杉种群产生了显著的遗传分化,分为Omote-sugi和Ura-sugi两个地理变种 (Kojo, 1987; Tsumuraetal.,2007, 2012, 2014),本文基于东亚地区全域的柳杉属种群研究也支持这一结论。
3.2 种群遗传结构
从STRUCTURE分析结果可看出,除中国庐山种群(LS)外,与其地理来源一致,所有柳杉属树种种群可分为柳杉和日本柳杉2个地理种群(K= 2),DAPC分析结果则分为更明显的4个谱系,分别是中国东南部种群(CHS)、中国庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi)。基于FST的NJ聚类结果,日本柳杉种群间有较近的亲缘关系,而柳杉种群间的遗传距离相对较远,显示出松散的聚类关系,分布地之间互有交叉。地理隔离(IBD)分析结果很好地证实了这一点,柳杉种群的地理距离与遗传距离间相关性不显著,而分布范围较小的日本柳杉种群则存在地理隔离现象。
一般来说,如果没有强烈的人为干扰和选择作用,植物会表现出明显的地理隔离。柳杉种群IBD消失的原因可能与人为干扰有关,古代频繁的贸易活动和长期人工栽培为遗传物质的转移提供了可能,从而使不同地理起源的柳杉种群产生了混杂。令人惊奇的是中国庐山种群(LS)并未归入柳杉聚类群,而是与日本柳杉有更近的亲缘关系。Cai等 (2020)认为中国庐山种群是日本柳杉引种而来,也有研究表明日本柳杉在20世纪初首先在庐山引种成功 (沈作奎等,2004)。但从我们分析结果来看,中国庐山种群不仅具有最高的遗传多样性,而且该种群的私有等位基因丰富度也显著高于日本柳杉,且DAPC分析进一步显示,中国庐山种群虽归入日本柳杉类群,但仍与日本柳杉的两个谱系Omote-sugi和Ura-sugi有较远的遗传距离。因此,关于中国庐山种群的起源和来源值得进一步深入研究。
3.3 种群进化历史
DIY ABC基于近似贝叶斯算法估算种群分化时间和有效种群数量。不同基因库(K= 2和K= 4)模拟分析结果共同显示: 柳杉属是由同一祖先种群(NA)分化而来,两组进化事件所对应的分化时间也较为一致,分别为1 010代(K= 2)和1 140代(K= 4)。柳杉通常需要20-30年开花结实,自然条件下,由于光照,林间郁闭等因素影响,其开花结实时间会有所推迟。参考日本柳杉研究结果 (Kimuraetal.,2014; Tsumuraetal.,2020),设定柳杉的自然更新时间为100~300年,最优进化事件所对应的种群分化时间在11万~34万年前,即柳杉和日本柳杉的最早分化时间发生在34万年前,结果与日本柳杉不同种群的分化时间大致相同 (Moriguchietal.,2019)。柳杉作为第三纪孑遗植物,其种的形成和分化可追溯至108.1百万年前,但其谱系分化时间仅只有34万年,类似于“活化石”银杏 (Ginkgobiloba) (Hohmannetal.,2018),其现存种群的谱系分化时间也仅39万年,表现为古老的孑遗种,年轻的谱系分化。根据化石资料,柳杉在侏罗纪晚期或更早起源于东亚东北地区 (于永福,1995),随后扩散至欧洲和北美各地,第三纪晚期和第四纪冰期的全球气候变化导致柳杉在欧洲和北美相继灭绝,仅在东亚有所保留,成为著名的第三纪孑遗植物。第四纪冰期的气候变化对其种群分化和种群数量产生了显著影响 (Tsumuraetal.,2020)。我们认为,11万~34万年前受第四纪三次亚冰期(庐山亚冰期)的影响,东亚地区柳杉种群数量迅速减少,退缩至中国东南部和日本西南部的避难所,分化为柳杉和日本柳杉种群,后随着间冰期的到来,海平面的上升使东海作为地理屏障隔离并加剧了东亚地区柳杉谱系间的分化。在此期间,第四纪亚冰期多次暴露的东海陆桥 (200万~130万年前,20万~1.5 万年前) 起到了“扩散廊道”的作用,为柳杉和日本柳杉提供了二次接触的机会。中国-日本植物区系中的很多植物,如蛛网萼(Platycraterarguta) (Qietal.,2014) 、领春木属(Euptelea) (Caoetal.,2016)、黄山梅属(Kirengeshoma) (Qiuetal.,2009; Yuanetal.,2014)也存在这种现象。综合现有证据,中国庐山种群可能是冰期避难所交汇地的残存种群,也可能是日本柳杉引种的结果 (Caietal.,2020),还需要后续更深入的研究。
3.4 柳杉属的分类及其科学保护
种群进化历史分析表明,虽然柳杉属树种起源古老,但柳杉和日本柳杉的谱系分化时间并不是很长,地理隔离使2个种群之间产生了一定程度的遗传变异和分化,但对于物种形成而言,34万年的遗传分化尚不足以形成新种。根据“整合物种概念”(刘建全,2016),我们认为柳杉和日本柳杉属于分化路上的姊妹种。与日本柳杉相比较,柳杉资源丰富,但总体遗传多样性不高,天然林破坏严重,使其遗传资源亟待有效保护。浙江省天目山种群(XTM)拥有最著名、保存最好的柳杉种群,在中国东南部柳杉种群中遗传多样性最高(Ho= 0.508),福建天宝岩(TBY)和福建武夷山种群(WYS)存在较多私有等位基因,种群变异特殊,应作为核心种群予以重点保护。安徽黄山(WT)、福建天宝岩高海拔地区(TB)以及福建南平(YTG)种群数量少,遗传多样性低,需优先予以保护。针对其他散生柳杉资源或人工林,应采取就地和迁地保护相结合的措施,通过建立保护小区的方式,加强管理,减少人类活动干扰。同时,加强种质资源的保护和收集,特别是大树、古树和风水林种群,以达到保护遗传多样性的目的。
4 结论
柳杉属树种可分为柳杉和日本柳杉2个地理种群,以及中国东南部种群(CHS)、中国庐山种群(LS)、日本海沿岸种群(Ura-sugi)和太平洋沿岸种群(Omote-sugi) 4个遗传谱系。柳杉属的总遗传多样性中等,种群间存在较高的遗传分化。柳杉和日本柳杉是由同一祖先种群在11万~34万年前同时分化而成,分化后两个地理种群拥有各自独立的进化路线。第四纪气候变化、地理隔离和全新世以来的人为干扰是东亚地区柳杉遗传变异格局形成的主要原因。与日本柳杉相比较,柳杉资源丰富,但总体遗传多样性不高,天然林种群破坏严重,亟待科学有效保护。
猜你喜欢
今日农业(2022年15期)2022-09-20
清华金融评论(2022年4期)2022-04-13
昆明医科大学学报(2022年1期)2022-02-28
现代企业(2021年2期)2021-07-20
阅读(低年级)(2018年8期)2018-05-14
中学生物学(2018年8期)2018-03-01
少年文艺·开心阅读作文(2014年2期)2014-02-27
古代文明(2014年1期)2014-02-23
文学教育·中旬版(2012年4期)2013-02-01
中学生物学(2008年6期)2008-08-29
