
84K杨PagMSBP1/2a基因对木质素合成的影响*
2022-10-16 07:47:56胥雅静王佳伟赵岩秋黄李超卢孟柱
林业科学 2022年6期
胥雅静 王佳伟 赵岩秋 江 成 黄李超 安 轶 曾 为 张 进 卢孟柱
(浙江农林大学林业与生物技术学院 省部共建亚热带森林培育国家重点实验室 杭州 311300)
木材即次生木质部,是维管形成层细胞增殖、分化的产物,包含木质部细胞次生细胞壁加厚的过程(Chaffeyetal., 2002)。次生壁是木材的主要成分,主要由木质素、纤维素、半纤维素等构成。其中木质素是植物中含量仅次于纤维素的第二大高聚物(Zhangetal., 2020)。
木质素可提供植物机械支撑力,并可作为天然屏障抵御生物侵害,在植物生长过程中起重要作用(Voelkeretal., 2010)。木质素在木材产业上应用有利有弊:一方面木质素存在会降低传统造纸的出浆率和纸张质量;另一方面木质素可作为生物质固碳并在新能源开发上得到利用(Wengetal., 2008)。基于对木质素的不同需求,利用转基因技术调控木材中木质素含量具有重要意义(李少锋, 2019)。
木质素由对香豆醇、松柏醇和芥子醇3种单体按照一定比例组合而成。木质素生物合成过程中有很多酶参与催化反应(Boerjanetal., 2003),根据木质素生物合成途径中不同酶的作用部位,可通过调节酶活性实现对木质素生物合成的调控。肉桂酸-4-羟化酶(C4H)、香豆酸-3-羟基化酶(C3′H)、阿魏酸-5-羟基化酶(F5H)是3种木质素单体合成的内质网定位细胞色素P450单氧酶,分别催化3种单体前体的合成。C3′H、C4H、F5H是具有锚点或成核中心的拓扑结构特征的Ⅰ型膜蛋白,能与该途径中其他酶结合调控木质素代谢通路(Bassardetal., 2012; Scottetal., 2016)。在苜蓿(Medicagosativa)中降低C4H、C3′H表达能够减少木质素含量,而F5H表达的改变不影响木质素的含量(Reddyetal., 2005)。在木本植物毛果杨(Populustrichocarpa)中有参与促进P450单氧酶形成复合物的膜蛋白,可以确保酶活性以进行高效催化(Chenetal., 2011)。
膜相关孕激素受体(MAPRs)是一类具有非共价细胞色素b5-like类固醇结合域(cytochrome b5-like haem/steroid-binding domain,Cyt-b5)的小分子蛋白质,可以调节P450酶活性(Chenetal.,2011; Kimuraetal.,2012; Ryuetal., 2017; Mifsudetal., 2002)。膜类固醇结合蛋白(MSBP)AtMSBP1是植物中鉴定的第一个膜类固醇结合蛋白,属Cyt-b5蛋白超家族(Kaoetal.,2005; Mifsud et al., 2002),MSBP1是拟南芥(Arabidopsisthaliang)中响应甾体信号的关键蛋白,在负调节细胞伸长、响应盐胁迫和调控木质素合成等过程中发挥重要作用(Shietal., 2011; Witzeletal., 2018; Yangetal., 2008; Gouetal., 2018)。鉴于MSBP1对植物生长发育的多重影响,该蛋白被进一步认为是质膜衍生的内吞小泡,可以通过参与生长素转运蛋白PIN2的循环调节生长素的再分配(Yangetal., 2008),以及通过介导BR信号受体BAK1的内吞作用调节BR信号(Shietal., 2011; Songetal., 2009)。近期在拟南芥中发现,MSBP1可与其同系物MSBP2形成复合体,并与3种木质素单体合成P450单加氧酶相互作用,从而形成MSBP-P450酶蛋白复合物。MSBPs可以维持相互作用的P450酶的活性和稳定性,对于可溶性酚酯和苯丙烷类单体生物合成十分重要 (Shietal., 2011; Gouetal., 2018)。
杨树是研究木本植物生长发育的模式物种,揭示杨树木材形成机制可为林木育种提供重要理论支撑。基于MSBP调节植物生长发育的重要作用,研究其在杨树木材形成,尤其是木质素合成中的作用具有重要意义。鉴于此,本研究通过构建系统发育树和实时荧光定量PCR,从银腺杨84K(PopulusalbaXP.glandulosa‘84K’)克隆得到拟南芥AtMSBP1同源基因PagMSBP1/2a,进行理化性质和亚细胞定位分析后,进一步构建PagMSBP1/2a基因过表达载体,并通过农杆菌介导的遗传转化法获得过表达PagMSBP1/2a的转基因杨树,分析植株生长发育和木质素含量,为研究MSBP1对木本植物木质素合成的调控作用提供重要依据。
1 材料与方法
1.1 试验材料及试剂
试验材料为银腺杨无性系84K,由浙江农林大学亚热带森林培育国家重点实验室林木木材品质研究团队保存。试验所用杨树均由组培苗扩繁而来,并由组培苗移栽至培养土中继续培养。杨树幼苗培养条件为16 h/8 h(光照/黑暗),温度25 ℃。取不同组织样品用于基因表达组织特异性分析。
试验所使Gateway入门载体pDNOR207、带GFP的过表达载体PMDC43、大肠杆菌(Escherichiacoli)DH5α和农杆菌(Agrobacteriumtumefaciens)GV3101菌种均购自维地生物。总RNA提取试剂盒、DNA片段回收试剂盒购于北京天根生化科技有限公司,反转录试剂盒PrimeScriptTMRT reagent Kit购自TaKaRa公司,PCR高保真酶Prime STAR HS、DNA marker等购自南京诺唯赞生物科技有限公司,Gateway试剂盒购于Life Technologies公司。
1.2 杨树MSBP基因获取及结构分析
取拟南芥AtMSBP1序列在杨树基因组中进行Blast分析,获取杨树MSBP基因的氨基酸序列; 通过MEGA7软件基于遗传距离的邻近法构建拟南芥和杨树的MSBP基因系统进化树(Kumaretal., 2016)。根据拟南芥和杨树的MSBP基因序列信息,通过GSDS2.0在线分析拟南芥和杨树MSBP基因的内含子与外显子结构,并绘制基因结构图(Huetal., 2015)。
1.3 RNA提取、cDNA合成及组织特异性表达分析
将不同组织样品在液氮中研磨成粉,顶芽、根、幼茎(1~3节间)、老茎(6~8节间)、叶的RNA提取与cDNA的合成分别采用RNAprep Pure Plant Kit(TIANGEN)和PrimeScriptTMRT reagent Kit(TaKaRa)完成,具体方法参见试剂盒使用说明。qPCR使用CFX96 Real-time PCR Detection System(Bio-Rad, Hercules, CA),反应体系按照ChamQ SYBR qPCR Master Mix(Vazyme)说明书,每个样品4个重复。PCR程序: 95 ℃,30 s预变性; 95 ℃,5 s变性,55 ℃,30 s退火,72 ℃,30 s延伸,35次循环。内参基因为PtACTIN。杨树MSBP1/2基因定量引物(表1)使用在线引物设计软件BatchPrime(https:∥probes.pw.usda.gov/batchprimer3/index.html)设计得到。利用杨树资源数据库(AspWood, http:∥aspwood.popgenie.org)获取MSBPs在木材形成过程中的表达情况 (Sundelletal., 2017)。
1.4 杨树PagMSBP1/2a基因的克隆与结构分析
根据毛果杨中PtMSBP1/2a(Potri.015G139800)基因序列,在银腺杨84K基因组数据库(Qiuetal., 2019)中进行Blast分析以得到其同源基因PagMSBP1/2a,使用Primer3设计特异引物PagMSBP1/2a-F和PagMSBP1/2a-R,以84K杨cDNA为模板,利用高保真聚合酶Prime STAR HS进行PCR扩增,得到目的基因PagMSBP1/2a后进行测序验证。使用在线网站SWISS-MODEL(https:∥swissmodel.expasy.org/interactive)和TMHMM网站(http:∥www.cbs.dtu.dk/services/TMHMM/)对MSBP1/2a蛋白的三级结构和跨膜结构域进行分析预测。通过NCBI Blast(https:∥blast.ncbi.nlm.nih.gov/)和植物基因组网站(https:∥phytozome.jgi.doe.gov/pz/portal.html)对不同物种中MSBP1氨基酸序列进行查找获取。利用DNAMAN软件对多物种MSBP1蛋白氨基酸序列比对分析,获得保守结构域。

表1 84K杨PagMSBP1/2基因克隆和定量PCR引物Tab.1 Primer sequences for 84K poplar PagMSBP1/2 cloning and qPCR analysis
1.5 载体构建及遗传转化
使用特异引物gw-PagMSBP1/2a-F和gw-PagMSBP1/2a-R,利用Gateway技术将PagMSBP1/2a基因连接至入门载体pDNOR207上,后分别连接至带GFP荧光蛋白标签的载体pMDC43和过表达载体pMDC32,转化至大肠杆菌并测序验证。将构建好的35S∷GFP-PagMSBP1/2a转化至农杆菌GV3101中,并与内质网膜Marker SPAtWAK2-CFP-HDEL (cyan) (Zengetal., 2016)共同在野生型本氏烟草叶片下表皮细胞中表达,过夜暗培养后室温培养3天取样共聚焦显微镜观察。将构建好的过表达载体35S∷pMDC32-PagMSBP1/2a利用农杆菌介导的叶盘法转化84K杨(Zhouetal., 2019),将筛选的转基因植株提取RNA进行qPCR分析,选择表达量较高的2个过表达转基因株系进行试验。
1.6 组织切片染色及木质素含量测定
每个株系各选3株在营养土中生长9周的杨树苗,观察植株生长的表型变化。取其基部节间约1 cm长,用振动式切片机(LEICA VT1 200 S)进行切片,厚度40 μm,分别用0.05%的甲苯胺蓝O(TBO)染液和1%间苯三酚-HCL染液染色,镜检观察,拍照记录。
取生长5个月的PagMSBP1/2a过表达和对照84K杨茎部作为材料,样品80 ℃烘干至恒质量,粉碎后过40目筛,称取约5 mg(记为W),采用乙酰溴法测定总木质素含量(Barnesetal., 2017; Fosteretal., 2010),参照科铭生物的木质素含量检测试剂盒说明书进行,按照以下公式计算木质素含量:
木质素(mg·g-1干质量)=((ΔA-0.006 8)×Vt×
10- 3×T)/(0.069 4×W)。
式中: ΔA为测定管OD280读值-空白管OD280读值;Vt为反应总体积2.04 mL;T为稀释倍数;W为样本质量。
2 结果与分析
2.1 杨树MSBP基因的序列分析
以拟南芥MSBP1蛋白序列在毛果杨和84K杨基因组中进行同源序列比对,分别得到MSBP同源基因。利用MEGA7软件构建杨树和拟南芥MSBP基因的系统发育进化树(图1A),杨树中MSBP成员根据进化树中两物种间的对应关系进行命名,其中与AtMSBP1、AtMSBP2同源关系较近的基因有3个,分别命名为PtMSBP1/2a、PtMSBP1/2b和PtMSBP1/2c; 与AtMSBP3同源的基因有2个,分别命名为PtMSBP3a和PtMSBP3b; 与AtMSBP4同源的基因有2个,分别命名为PtMSBP4a和PtMSBP4b。84K杨中MSBP命名同理。同时利用GSDS2.0进行基因结构分析,构建结构图(图1B)。杨树中MSBP1同源基因PtMSBP1/2a与PtMSBP1/2b长度均为2 kb左右,而PtMSBP1/2c长度不到1 kb,在基因结构上3个基因都各包含2个外显子和1个内含子,与拟南芥MSBP1基因结构相比具有保守性,表明进化历程中结构相对保守。
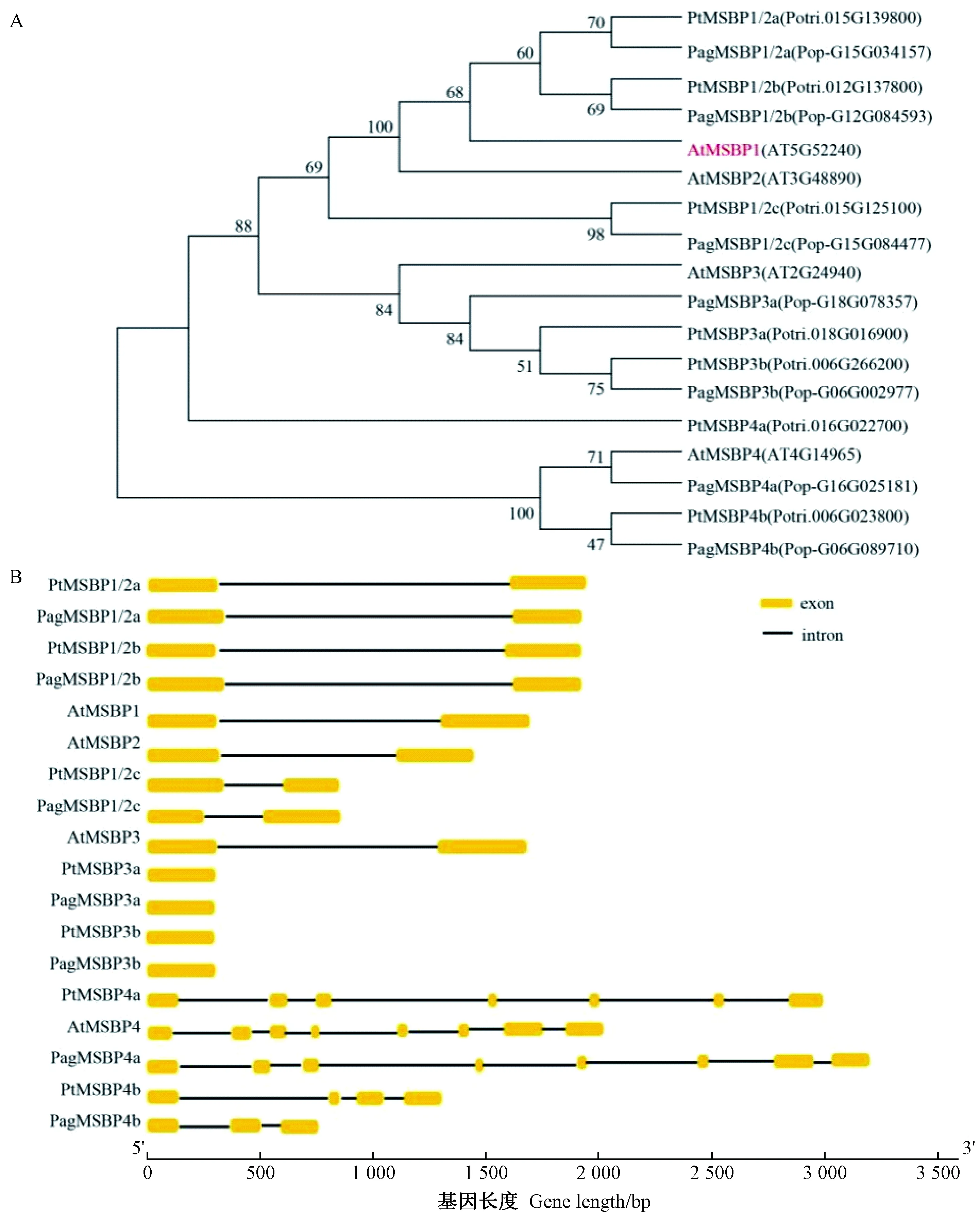
图1 拟南芥与杨树中MSBP基因分析Fig. 1 Phylogenetic tree and gene structure of MSBP from Arabidopsis and PopulusA.杨树MSBP与拟南芥同源蛋白的系统进化树分析 B.拟南芥与杨树MSBP基因结构分析。Pt:毛果杨; At: 拟南芥; Pag: 84K。A. Phylogenetic tree analysis of homologous proteins of Populus and Arabidopsis. B. Structural analysis of MSBP genes in Arabidopsis and Populus. Pt: Populus trichocarpa; At: Arabidopsis thaliana; Pag: 84K.
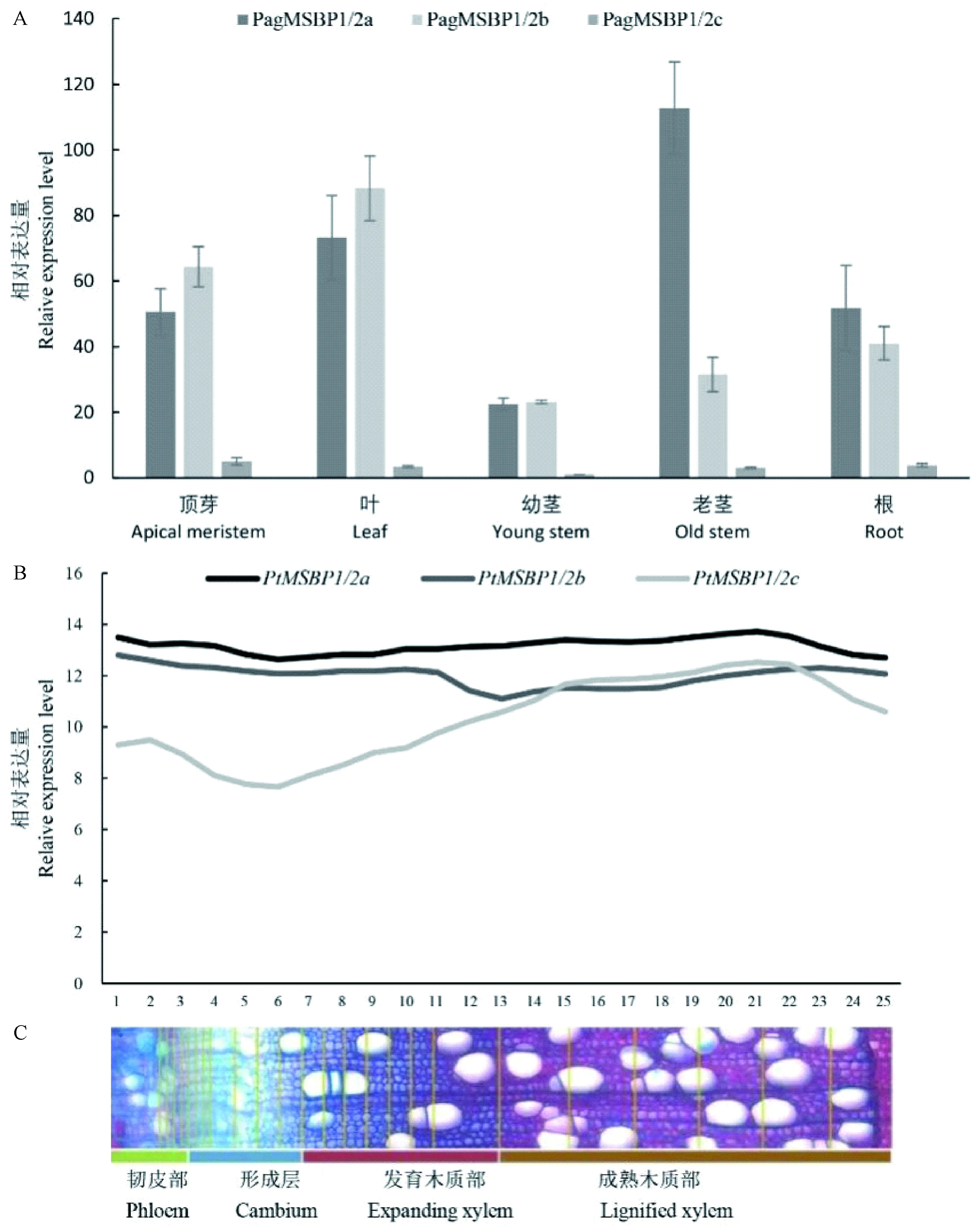
图2 PtMSBP1/2基因表达分析Fig. 2 Tissue-specific expression analysis of PtMSBP1/2A. qPCR分析PagMSBP1/2a、PagMSBP1/2b和PagMSBP1/2c基因在84K杨树顶芽、叶、幼茎、老茎和根中的表达; B.杨树PtMSBP1/2a、PtMSBP1/2b和PtMSBP1/2c基因在韧皮部、形成层及木质部中的表达。A. qPCR analysis of the expression of PagMSBP1/2a, PagMSBP1/2b and PagMSBP1/2c genes in 84K poplar top buds, leaves, young stems, old stems, and roots; B. Analysis of PtMSBP1/2a, PtMSBP1/2b PtMSBP1/2c gene in phloem, cambium and xylem expression.
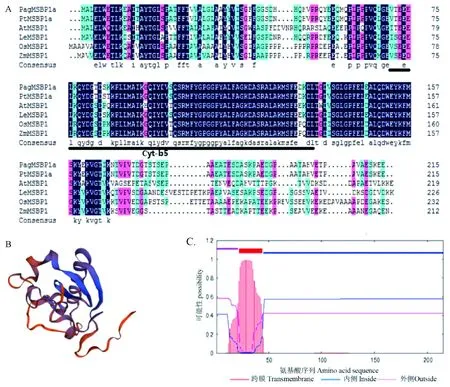
图3 PagMSBP1/2a序列结构分析Fig. 3 The sequence analysis of PagMSBP1/2aA. 6种植物同源MSBP1蛋白氨基酸序列比对,下划线代表Cyt-b5保守结构域; B.PagMSBP1/2a蛋白三级结构预测; C. PagMSBP1/2a氨基酸序列的跨膜结构域预测,峰值即代表跨膜结构域。A. Alignment of the amino acid sequence of MSBP1 protein with homologous genes of other species, the underline represents the Cyt-b5 conserved domain; B. PagMSBP1/2a protein tertiary structure prediction; C. The transmembrane domain prediction of PagMSBP1/2a amino acid sequence, the peak value represents the transmembrane domain.
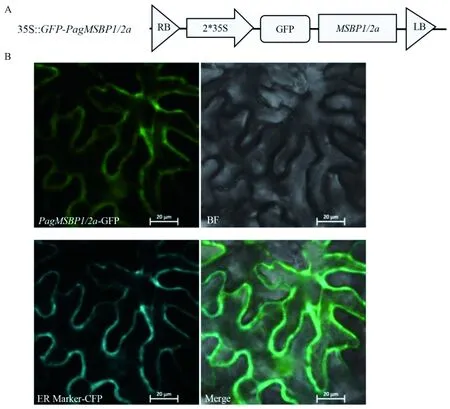
图4 PagMSBP1/2a亚细胞定位分析Fig. 4 Subcellular localization of PagMSBP1/2aA. GFP-PagMSBP1/2a亚细胞定位载体图; B.荧光共聚焦表明PagMSBP1/2a定位在内质网。A. PagMSBP1/2a subcellular localization vector map; B. GFP-PagMSBP1/2a localized on the endoplasmic reticulum by fluorescence confocal microscope tracking.
2.2 杨树MSBP基因组织表达分析
取土培6个月的84K杨的顶芽、叶、嫩茎、老茎和根提取RNA,qPCR分析PagMSBP1/2a、PagMSBP1/2b和PagMSBP1/2c基因表达情况,结果表明,3个基因在顶芽、叶、茎和根中均有表达,其中PagMSBP1/2c在各组织的表达量相对较低。PagMSBP1/2a在老茎表达量最高,为表达量最低的幼茎中的6倍,另外,PagMSBP1/2a在茎中的表达显著高于其他2个基因(图2A)。参考AspWood数据库(http:∥aspwood.popgenie.org)对3个基因在韧皮部、形成层及木质部中的表达谱进行数据分析,发现杨树MSBP1的3个成员均在成熟木质部表现出高表达,同时根据相对表达量分析,发现PtMSBP1/2a在木质部与韧皮部中的表达相对较高(图2B)。为此,本研究选择PagMSBP1/2a基因深入研究其在杨树木材形成中的调控作用。
2.3 PagMSBP1/2a基因克隆及序列分析
对84K杨中克隆出的PagMSBP1/2a基因分析发现,PagMSBP1/2a包含一个全长648 bp 的CDS,编码215个氨基酸,由2个外显子组成,分子量为2.33 kDa,理论等电点(PI)为4.53。利用DNAMAN软件进行多重序列比对表明,PagMSBP1/2a蛋白序列与毛果杨、拟南芥、水稻(Oryzasativa)、玉米(Zeamays)和番茄(Solanumlycopersicum)5种植物的MSBP1蛋白序列相似度较高,一些区域存在明显的保守序列。MSBP蛋白在单子叶和双子叶植物中具有一定保守性,且都有Cyt-b5结构域(图3A)。应用ExPAsy提供的SWISS-MODEL 对PagMSBP1/2a蛋白的三级结构进行预测,以人类膜相关孕激素受体组分1(PGRMC1)蛋白(4x8y.1.A)为模板,同源建模PagMSBP1/2a蛋白的完整氨基酸序列,预测所得模型见图3B,分析表明此蛋白具有类固醇结合结构域,是一个α+β混合结构,2对α-螺旋在β-折叠的一侧形成三明治口袋结构。采用TMHMM网站对MSBP1蛋白的跨膜结构域进行预测,结果表明,PagMSBP1/2a在N端第22~44位氨基酸形成跨膜结构域(图3C)。
2.4 亚细胞定位分析
将35S∷GFP-PagMSBP1/2a重组载体在烟草表皮细胞中瞬时表达,分析其在细胞中的定位。结果表明,在注射含有35S∷GFP-PagMSBP1/2a过表达载体农杆菌的本氏烟草叶片表皮细胞中,在488 nm波长的激发光下使用共聚焦荧光显微镜观察到的GFP荧光信号与内质网Marker重合,表明PagMSBP1/2a蛋白定位于内质网(图4)。
2.5 PagMSBP1/2a过表达载体的构建与遗传转化
为深入研究PagMSBP1/2a基因对杨树生长发育的作用,实验构建了35 S∷35S∷PagMSBP1/2a表达载体(图5A),采用农杆菌介导的叶盘法转化杨树,获得了过量表达的转基因株系(PagMSBP1/2a-OE)。通过PCR鉴定和qPCR检测表达情况后,获得PagMSBP1/2a过量表达的阳性转基因苗。选取2个表达量较高的株系#24、#33进行试验(图5C)。生长9周的过表达PagMSBP1/2a杨树株高生长明显低于对照(图5B)。统计结果显示,过表达PagMSBP1/2a杨树2个株系株高分别比对照株高减少21%、5%。
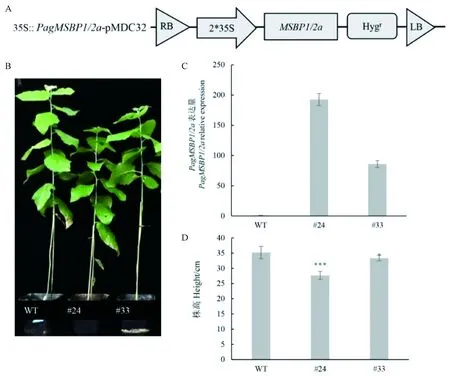
图5 PagMSBP1/2a过表达载体图谱及转基因植株Fig. 5 PagMSBP1/2a overexpressed vector and transgenic PopulusA.PagMSBP1/2a过表达载体图 B. PagMSBP1/2a过表达植株表型C.选定的过表达植株PagMSBP1/2a基因的相对表达量。A. Vector of PagMSBP1/2a overexpression. B. Phenotype of PagMSBP1/2a overexpressed Populus C. Relative expression of PagMSBP1/2a gene in selected overexpressed plants.
2.6 过表达PagMSBP1/2a对木质素合成的影响
MSBP1是调控木质素合成过程的重要支架蛋白,为研究PagMSBP1/2a基因在木质素合成中发挥的作用,对PagMSBP1/2a过表达株系#24和#33进一步解剖分析。观察茎横切面染色情况,发现过表达PagMSBP1/2a杨树比对照组间苯三酚-HCL染色浅,说明其木质素的含量有所降低。利用乙酰溴法测定木质素含量,在过表达PagMSBP1/2a株系#24和#33中,木质素含量与对照组相比分别减少34.6%、26.5%。
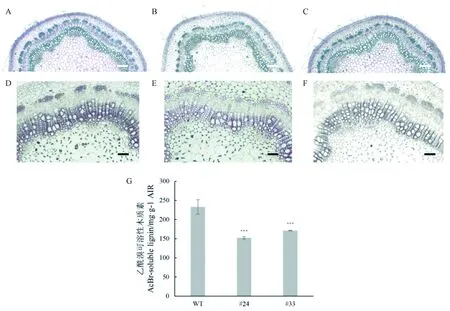
图6 过表达PagMSBP1/2a 影响木质素合成Fig. 6 PagMSBP1/2a affects lignin biosynthesis比例尺Scale bar in A-C is 200 μm; and in D-E is 80 μm.; C: 第7节间横切片甲苯胺蓝O染色(A: 对照植株,B-C: 转基因株系#24、#33); D-F: 第7节间横切片间苯三酚-HCL染色(D: 对照植株,E-F: 转基因株系#24、#33); G.乙酰溴法测定木质素含量。t检验P值用星号表示,***表示差异极显著(P<0.01)。A-C. 7th internode transverse section stained with TBO (A, control plants; B, C, transgenic plants #24, #33, respectively); D-F. 7th internode transverse section phloroglucinol-HCL staining (D, control plants; E, F, transgenic plants #24, #33, respectively); G. Acetyl bromide method to determine the content of lignin. The P value of t test is indicated by stars,*** indicate extremely significant difference (P<0.01).
3 讨论
木材形成是一个极为精细的过程,对于次生壁的合成机制已进行大量研究工作,目前木质素的生物合成及调控通路解析取得较大进展(Shietal., 2010)。最近表明,MSBP1作为动物膜相关孕激素受体PGRMC1(Falkensteinetal., 1996)同源基因在木质素合成中有重要作用(Shietal., 2011; Gouetal., 2018)。本研究对MSBP在杨树中的7个成员进行分析,包括进化关系、基因结构及其在杨树组织中的表达等。分析发现,杨树中MSBP蛋白结构相对保守,均具有Cyt-b5保守结构域(Mifsudetal., 2002),推测MSBP在功能上可能具有高度保守性。
结合组织定量分析以及AspWood在线分析表明,MSBP1/2a基因在成熟茎段的木质部表达水平高(图2)。拟南芥MSBP1以前被认定为质膜蛋白,可作为类固醇/BR激素信号成分负调控细胞伸长(Yangetal., 2005),后来荧光蛋白GFP介导的亚细胞定位研究表明,MSBP1主要是一种内质网驻留蛋白(Gouetal., 2018),本研究的PagMSBP1/2a亚细胞定位也证实了这一观点(图4)。MSBP1/2a的表达模式和定位分析表明MSBP1可能对茎的生长发育有影响,因此本文对84K杨中在木质部组织高表达的PagMSBP1/2a进行深入研究。
NST是NAC家族中参与调节次生壁加厚 (Mitsudaetal., 2007) 的重要转录因子,有研究证明在nst-1突变体中MSBP基因的表达降低了50%左右 (Gouetal., 2018),同时发现在杨树的发育木质部中MSBP同源基因表达水平较高 (Baoetal., 2013),这证明MSBP1和MSBP2与次生细胞壁合成和木质素的形成相关。在拟南芥突变体msbp1进一步的实验中,抑制MSBP2表达会致使C4H、C3′H和F5H蛋白水平的大幅下降以及C4H酶催化活性的降低,使植株呈现木质化程度低的表型(Gouetal., 2018)。采用间苯三酚-HCL对6周大的拟南芥基茎测定维管束和束间纤维的染色强度发现,双基因敲除系与野生型msbp2-1相比明显较弱,表明双基因敲除植株维管中木质素总量降低(Gouetal., 2018)。本研究通过过表达杨树PagMSBP1/2a基因,乙酰溴法测定转基因植株成熟茎段的木质素含量,PagMSBP1/2a过表达植株中的木质素含量明显低于对照植株(图6G)。本研究中过表达MSBP1导致木质素含量降低的结论,与已报道的减少MSBP1表达会降低木质素含量相矛盾。原因可能是植物在调节木质素合成途径中对MSBP1蛋白具有严苛的要求。MSBP1做为支架蛋白连接P450酶在内质网膜形成蛋白复合体,维持酶的活性和稳定性,MSBP1蛋白的增多极有可能打破了复合体的调控平衡,导致3个P450酶的排列及功能发生变化,从而直接影响复合体催化木质素的形成。有研究称在苜蓿中C3′H、C4H表达下降的植株中乙酰溴化木质素含量降低,而F5H表达降低不影响总含量。C4H、F5H的降低都提高了木质素G单体的比例,并且F5H的降低还增加了硫代酸解木质素的含量(Reddyetal., 2005; Stewartetal., 2009)。说明C3′H、C4H、F5H在木质素合成途径中存在精密的调控平衡中,MSBP作为木质素合成的关键调控因子,与相关酶的互作关系直接影响其调控作用的发挥,且PagMSBP1/2a与其他成员之间是否存在相互作用也有待进一步研究。本研究表明了该基因过量表达可以降低木质素的含量,但植株生长受到了较大影响。因此若采用该基因启动子进行木质部细胞特异表达,则可能实现PagMSBP1/2成员对木质素合成过程的精准调控。
4 结论
本研究克隆了84K杨PagMSBP1/2a基因,预测其编码膜蛋白,与其定位在内质网膜相一致。PagMSBP1/2a基因在成熟茎中表达量较高,尤其在木质部发育中表达量最高。通过高表达PagMSBP1/2a证明了其参与木质部发育过程,导致植株矮化,木质素含量减少。PagMSBP1/2a在木材形成过程中的调控作用,可以作为靶基因进行分子育种,调控木质素的含量。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
上海包装(2019年8期)2019-11-11 12:16:14
时代英语·高一(2019年1期)2019-03-13 10:29:48
生命科学研究(2018年1期)2018-05-29 01:12:47
作文评点报·低幼版(2017年24期)2017-06-19 14:59:13
喜剧世界(2017年10期)2017-06-01 12:39:29
上海农业学报(2017年3期)2017-04-10 12:39:14
天津造纸(2016年1期)2017-01-15 14:03:29
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
中国造纸学报(2015年1期)2015-12-16 19:35:25
