
基于气候和寄主因素的栗山天牛中国成灾和扩散风险评估
2022-10-16 07:47张宇凡党英侨王小艺
林业科学 2022年6期
张宇凡 党英侨 王小艺
(1.中国林业科学研究院森林生态环境与自然保护研究所 国家林业和草原局森林保护学重点实验室 北京 100091;2.广东省云浮市林业科学和技术推广中心 云浮 527300)
栗山天牛(Massicusraddei)隶属于鞘翅目(Coleoptera)天牛科(Cerambycidae)山天牛属(Massicus),是一种严重危害森林健康的蛀干害虫(陈世骧等,1959),现仅集中分布于东亚地区(中国、朝鲜半岛、日本、越南和俄罗斯远东地区),其中在我国分布范围较广,多省区市均见报道(孙永平,2001; 骆有庆等,2005; 国家林业局森林病虫害防治总站,2008;2010; 王直诚,2013; 国家林业和草原局森林病虫害防治总站,2019)。栗山天牛的寄主种类主要集中在壳斗科(Fagaceae)的栎属(Quercus)、栗属(Castanea)和锥栗属(Castanopsis),在我国北方危害栎类山区林木,在南方则主要危害板栗(Castaneamollisima)等经济林木(林家全等,2016)。在该虫的所有分布区中,仅辽宁和吉林省严重受灾,而在中国的其他省份和其他国家均不成灾(EPPO, 2018)。对于栗山天牛在东北林区的局部成灾机理,至今还未有明确定论,杨忠岐等(2011)和王小艺等(2012)认为东北林区栗山天牛种群有别于其他地理种群,其幼虫生长发育整齐且成虫表现出3年为周期的羽化行为,这符合前人对周期性昆虫的定义(Bulmer, 1977; Heliövaaraetal., 1994)。此外,前人研究表明,在相同昆虫种类中,周期性种群通过天敌避免效应(predator avoidance)或捕食者饱足效应(predation satiation)使其种群密度远高于每年均有成虫羽化的非周期性种群(Lloydetal.,1966; Martinetal.,1990)。Zhang等(2020)提出栗山天牛在东北林区局部成灾与其特殊的周期性羽化行为存在因果关系,并否定周期性羽化行为来源于与其他地理种群缺乏基因流动而造成的区域性种群隔离的可能,认为区域性生态因子、气候和寄主对该虫的适生等级为该虫周期性和局部成灾产生的主要原因。孙永平(2001)发现在辽宁宽甸林区内,不同寄主受栗山天牛危害的等级存在显著差异,蒙古栎(Quercusmongolica)和辽东栎(Quercuswutaishanica)的受害率最高,分别达到68.9%和48.6%; 槲树(Quercusdentata)和槲栎(Quercusaliena)的受害率次之,分别为14.1%和13.0%; 其他栎属和栗属树种的受害率较低或不受危害。EPPO(2018)也曾提及在栗山天牛严重危害的蒙古栎林旁栽植的板栗林不受危害的现象。因此,辽东栎和蒙古栎可能是栗山天牛最偏嗜取食的树种,槲栎和槲树可能为中等偏嗜取食的树种,而其他受害的栎属和栗属树种,包括麻栎(Quercusacutissima)、板栗、栓皮栎(Quercusvariabilis)、枹栎(Quercusserrata)、青冈栎(Quercusglauca)、锥栗(Castaneahenryi)与乌冈栎(Quercusphillyraeoides),为低偏嗜取食树种。以辽宁省宽甸县为代表的东北林区的栎林严重受灾可能受到了栗山天牛成虫周期性羽化行为和易感寄主树种成片分布的双重影响。
物种分布模型(species distribution models, SDM)是基于气候、寄主、地理和人为活动等多种因素预测物种潜在分布区的一种工具(许仲林等,2015)。目前常见的物种分布模型包括CLIMEX、DOMAIN、GARP、HABITAT和Maxent(Chejaraetal., 2010),其中最大熵模型Maxent(maximum entropy model)更具优越性,其对适生区的预测主要由物种分布地点、气候环境变量图层和软件参数设置3部分来决定,具有模拟精确度高、操作简便、运行稳定且所需时间短等特点(Elithetal., 2006; 张路,2015)。蛀干害虫幼虫隐蔽在树体内钻蛀取食,仅成虫期在树体外进行繁殖行为(Fengetal., 2016)。因此,只要温湿度为主的气候条件满足蛀干害虫基本生存所需,其扩散和定殖将更大程度上依赖于寄主树种的分布和人为协助的木材调运(Sobek-Swantetal., 2012; Shimetal., 2020; Dangetal., 2021)。
在前人对有害生物的潜在适生区研究中,通常仅基于其现有分布点利用气候因素来预测该害虫在未分布或非广泛分布国家和地区的潜在适生情况,而未考虑寄主植物的潜在影响(王茹琳等,2017; 宋光远等,2018; 范靖宇等,2019; Tangetal., 2019)。本研究的目的是基于栗山天牛在中国的现有分布点、气候变量和不同偏嗜取食程度的寄主树种分布数据,采用Maxent和ArcGIS评估该虫在我国不同地理区域的适生等级,结果与该虫实际分布和危害情况相对比,以明确寄主和气候因素对该虫局部成灾的作用,以及在我国未分布区域的扩散定殖风险,以期为栗山天牛成灾区的生态防控以及未分布区的检疫和早期预警措施提供理论依据。
1 研究方法
1.1 栗山天牛分布数据
Maxent模型模拟需要物种分布数据和环境数据,分布数据即为“存在”点的经纬度信息。栗山天牛的137个中国分布点通过以下2种方式获得: 1)文献整理(孙永平,2001; 王直诚,2003; 国家林业局森林病虫害防治总站,2008,2010; 王直诚,2013; 徐志华等,2013; 国家林业与草原局森林与草原病虫害防治总站,2019; 虞国跃,2020),共获得129个分布点数据(表1); 2)林间实地踏查和灯诱,共获得8个分布点数据(依次为山东烟台昆嵛山和栖霞方山蚕场、安徽合肥西庐寺和蜀山烈士陵园、河南西峡寺山森林公园、福建南平茫荡山、云南屏边大围山和文山州盘龙森林公园)(表1)。在excel中按照物种名称、经度和纬度整理分布点信息,并保存为csv格式用作后续Maxent建模分析(Phillips, 2017)。本文依据文献报道点来进行栗山天牛潜在适生区预测,利用实际调查分布点来检验预测结果的准确性。

表1 栗山天牛中国境内分布和危害情况Tab.1 Distribution and infestation conditions of M. raddei in China
1.2 寄主树种分布数据
在中国,栗山天牛的寄主树种共11种(依次为蒙古栎、辽东栎、槲栎、槲树、麻栎、板栗、枹栎、乌冈栎、锥栗、青冈栎与栓皮栎)(EPPO, 2018),各分布点主要通过全球生物多样性信息网(Global Biodiversity Information Facility, GBIF, https:∥www.gbif.org/)获得,并通过地球在线(https:∥www.earthol.com.)验证分布点的准确性。通过删除相同、信息不全和位于海上的点,最终共获得寄主树种在中国的分布点2 548个。基于栗山天牛对每种寄主树种的喜食程度进行赋值,预测栗山天牛的中国潜在适生区。
1.3 地图数据与气候数据
本研究所使用的中国国界及省界1∶400万地图来自国家基础地理信息网(National Geomatics Center of China, NGCC, http:∥ngcc.sbsm.gov.cn/)。1970—2000年的19个气候因子(bio01~bio19)的平均值栅格数据在WorldClim数据集(http:∥www.worldclim.org.)下载,数据空间分辨率为2.5弧分。使用ArcGIS掩膜提取出中国范围内的气候变量TIFF格式文件,并转换成ASCII格式后保存。将栗山天牛的分布数据和ASCII格式的气候变量数据导入ArcGIS中,利用空间分析工具箱提取采样,将采样结果导入SPSS 20.0版本进行皮尔森相关性分析(Pearson correlation analysis),对19个变量进行两两相关性检验,得到气候变量间的相关系数矩阵。相关系数绝对值大于0.9定义为高度相关。Maxent模型初次构建时拟合1次,得到19个气候因素对栗山天牛潜在分布区的贡献率大小。去除高相关性的变量组中贡献率小的变量,最后选择出独立且具有较大生物意义的8个环境变量,用作Maxent后续建模分析。
1.4 适生区预测
1.4.1 仅基于气候因素的适生区预测 将栗山天牛中国分布点的csv文件和筛选出的气候变量一起导入MaxEnt软件中,软件运行的参数设置中选用刀切法Jackknife来检验各气候因子的权重,随机抽取分布数据的25%作为测试集,75%为训练集,其他参数为软件默认。模型拟合10次,以10次拟合结果的平均值作为最终结果(Phillips, 2017; Tangetal., 2019)。
1.4.2 仅基于寄主现有分布的适生区预测 为了将栗山天牛寄主植物的分布数据更加精确地对应到每个县区内,将筛选得到的栗山天牛寄主植物在中国的分布数据和中国县界地图数据一起导入ArcGIS中进行空间连接,连接后导出txt文件并加载到excel中分析和整理。依据孙永平(2001)报道的辽宁宽甸林区内不同寄主树种的受危害程度,每个县/区中存在敏感树种(蒙古栎和辽东栎)的分布点赋值为10,存在中适树种(槲树、槲栎)的分布点赋值为5,仅存在低适树种(麻栎、板栗、栓皮栎、枹栎、青冈栎、锥栗和乌冈栎)的分布点赋值为2,无栎树分布点赋值为0。计算得到各个县/区寄主赋值总量n后,按照n=0、2、5和10将所有县/区划分为1~4级(依次为非适生区、低适生区、中适生区和高适生区)。将整理得到的新csv文件重新导入ArcGIS中,利用Join工具将该文件与县界地图相关联,并生成shp文件。之后利用ArcGIS的转换工具将该文件转成TIFF栅格文件,value值选择事先设置好的1~4等级,得到寄主树种风险等级图。采用ArcGIS内的转换工具将TIFF文件转为ASCII文件后,将其作为寄主因素与栗山天牛中国分布点的csv文件导入Maxent中,随机抽取分布数据的25%作为测试集,75%为训练集,其他参数为软件默认。模型拟合10次,以10次拟合结果的平均值作为最终结果,进而得到基于寄主因素的栗山天牛在中国的潜在适生区ASCII文件。
1.4.3 基于气候和寄主因素的适生区预测 栗山天牛成虫羽化和繁殖活动受到温湿度限制,过低温度和过高湿度将会降低羽化成虫数量(杨忠岐等,2011); 在树体内隐蔽生活的幼虫也将受到环境温度和寄主适生等级的影响,当树体内的低温低于幼虫过冷却点或树种本身表现出对幼虫生长发育的物理或者化学抗性防御,均将造成该蛀虫在此分布区域的非适生性(DeSantisetal., 2013; Shimetal., 2020)。因此本文认为气候条件、寄主种类和分布对栗山天牛的潜在分布概率的作用大小是相同的,基于寄主或气候因素获得的栗山天牛潜在分布图显示的是相关变量影响下每个地区栗山天牛的潜在分布概率。采用ArcGIS中的Plus和Divide功能将这2个ASCII文件叠加,以求得每个地区仅基于气候因素和仅基于寄主因素的2个栗山天牛潜在分布概率的平均值,将平均值作为每个地区在寄主和气候因素的共同影响下的栗山天牛潜在分布概率,叠加得到的ASCII文件即为基于寄主和气候因素的栗山天牛适生区预测图。
1.5 模型精确度检验
本试验采用受试者工作曲线(receiver operating characteristic curve, ROC)分析法中的曲线下面积值(area under curve, AUC)及遗漏率(omission rate, OR值)评价模型的准确性。AUC检验值显示的是模型预测能力,AUC>0.9说明模型拟合良好。遗漏率计算的是适宜值低于指定阈值的测试地点的比例。其中,最小训练集阈值遗漏率(minimum training presence threshold omission rate, ORmtp)指的是模型任何训练位置预测的最小值,用来表明训练点内最不适合的环境; 相似地,10%训练集阈值遗漏率(10% training threshold omission rate, OR10)指的是排除了训练适宜值中最低的10%后使用的最小值。在理想模型中,ORmtp和OR10的预期值分别为0和0.1,实际值高于预期值时说明模型过拟合,否则说明模型拟合良好(Radosavljevicetal., 2014; Arthuretal., 2019)。
1.6 适生区划分和面积计算
将Maxent软件输出的和图层叠加后的ASCII格式文件导入ArcGIS中,利用空间分析工具的再分类功能(Reclassify)中的自然间断法(Natural Breaks)自动将适生程度划分为4个等级(非适生区、低适生区、中适生区与高适生区)。采用ArcGIS软件中的投影与转换工具箱内的栅格投影功能将地理坐标系(GCS_WGS_1984)转为投影坐标系(WGS_1984_Web_Mercator_Auxilialy_Sphere),依次计算基于气候因素、寄主分布和2个因素的栗山天牛不同适生等级区域所占比例,然后在Excel 2019中作图。
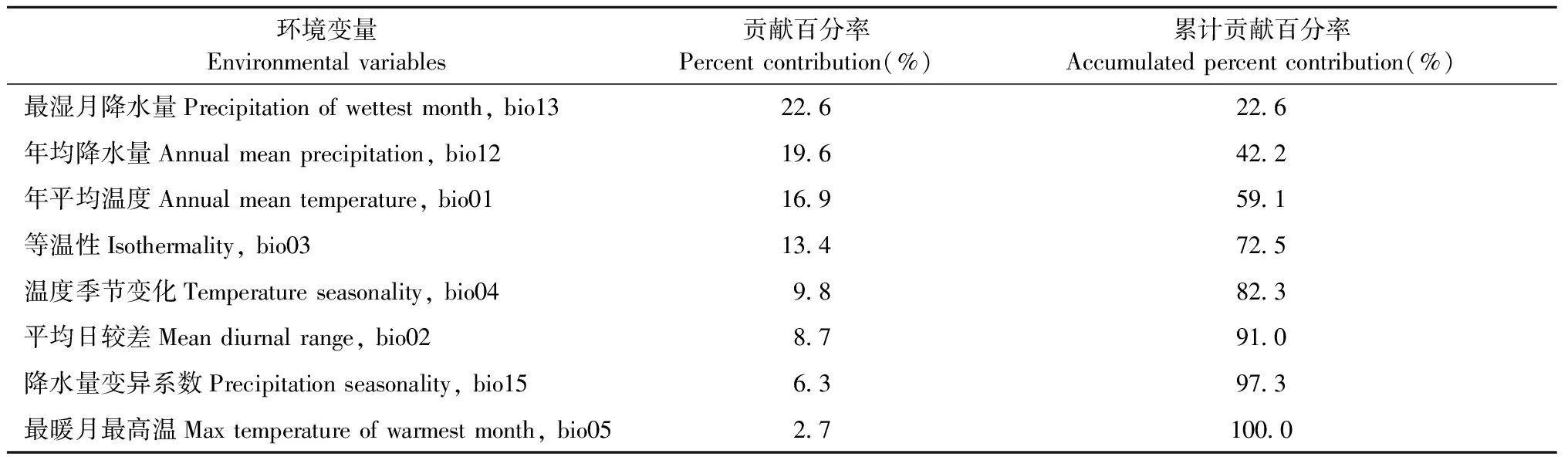
表2 影响栗山天牛分布的环境变量贡献率和累积贡献率Tab.2 The accumulated contribution of selected environmental variables to the potential distribution of M. raddei defined by Maxent model
2 结果与分析
2.1 仅基于气候因素的栗山天牛潜在适生区
基于气候变量的栗山天牛中国适生区预测模型AUC值为0.921,ORmtp和OR10值分别为0.048和0.250,虽然ORmtp和OR10值均略高于预测值0和0.1,存在一定程度上的过拟合现象,但考虑到AUC值大于0.9,综合来看模型拟合较好(图1)。用作栗山天牛中国潜在分布区预测的气候变量共8个,包括bio13(最湿月降水量)、bio12(年平均雨量)、bio01(年平均温度)、bio03(等温性)、bio04(温度季节性变化标准差)、bio02(平均日较差)、bio15(降水量变异系数)与bio05(最暖月最高温度),其贡献率依次为22.6%、19.6%、16.9%、13.4%、9.8%、8.7%、6.3%与2.7%(表2)。其中前3个气候变量对栗山天牛潜在分布概率的响应曲线均呈现先上升再下降的趋势,而昼夜温差与年温差比值(等温性)对栗山天牛潜在分布概率的响应曲线呈现下降、上升再下降的趋势。最湿月降水量为200 mm、年平均雨量为750 mm、年平均温度为4 ℃和等温性为22 ℃下最适合栗山天牛分布和生存(图2)。
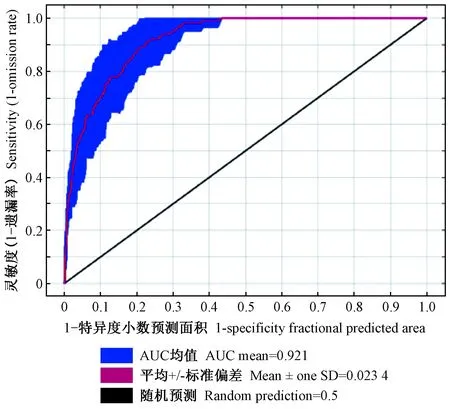
图1 模型适用性检验AUC值Fig. 1 AUC value of test on applicability of the Maxent model
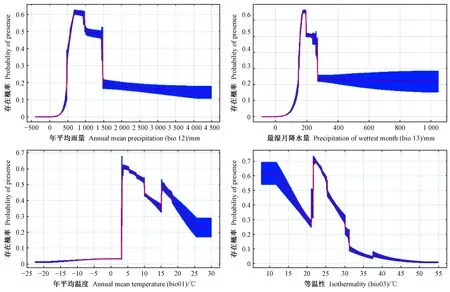
图2 Maxent模型中栗山天牛存在概率随主导环境因子的响应曲线Fig. 2 Response curves of leading climatic factors to presence probability of M. raddei in Maxent model
基于气候因素的栗山天牛潜在适生区结果显示,栗山天牛在除了新疆和青海以外的中国大部分地区均表现出适生性。其中高适生区主要集中在辽宁和吉林大部分地区、河北北部和浙江北部,绝大多数南方省份和黑龙江南部地区为低、中适生区,内蒙古大部分地区均表现为非适生性,仅与辽宁省和吉林省接壤的地区为低适生区。然而这一预测结果与实际调查得到的栗山天牛的分布情况存在差异,即福建省南平市茫荡山、云南省屏边大围山和文山州盘龙森林公园等多个实际调查到的分布点并未涵盖在预测的适生区范围内(表3、表6)。
2.2 仅基于寄主因素的栗山天牛潜在适生区
基于寄主因素的栗山天牛潜在适生区结果显示,栗山天牛的潜在适生等级表现为南北地区明显的地理分化格局,最喜取食的寄主树种蒙古栎和辽东栎主要分布于我国北方大部分省份(黑龙江、内蒙古、吉林、辽宁、北京、河北、山西、陕西、宁夏、甘肃)和四川北部,为栗山天牛的高适生区; 南部各省主要分布有其他次要喜食的寄主树种,为栗山天牛的中、低适生区,与仅基于气候因素的栗山天牛潜在适生区结果相似,除了新疆和青海外,其他省份均存在栗山天牛的适生区(表4、表6)。
2.3 基于气候和寄主因素的栗山天牛潜在适生区
联合寄主因素和气候因素叠加而成的栗山天牛潜在适生区结果显示,栗山天牛适生总面积变大,其中,高适生区范围有所缩减,主要集中于辽宁、吉林、河北、北京,但低、中适生区范围有所增加。预测区涵盖了栗山天牛所有实际调查得到的分布点,结果与栗山天牛目前在我国的危害情况更加吻合(表1、表5、表6、图3)。
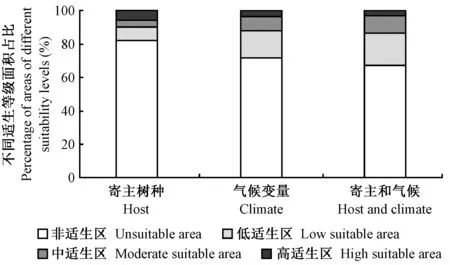
图3 基于寄主因素、气候因素、寄主和气候共同影响下的栗山天牛在中国不同适生等级面积所占百分比Fig. 3 Percentage of areas of different suitability levels for M. raddei based on host distribution, climatic variables and the combination of two factors

表3 仅考虑气候因素的栗山天牛中国适生区①Tab.3 Potential distribution of M. raddei based on climatic factors in China
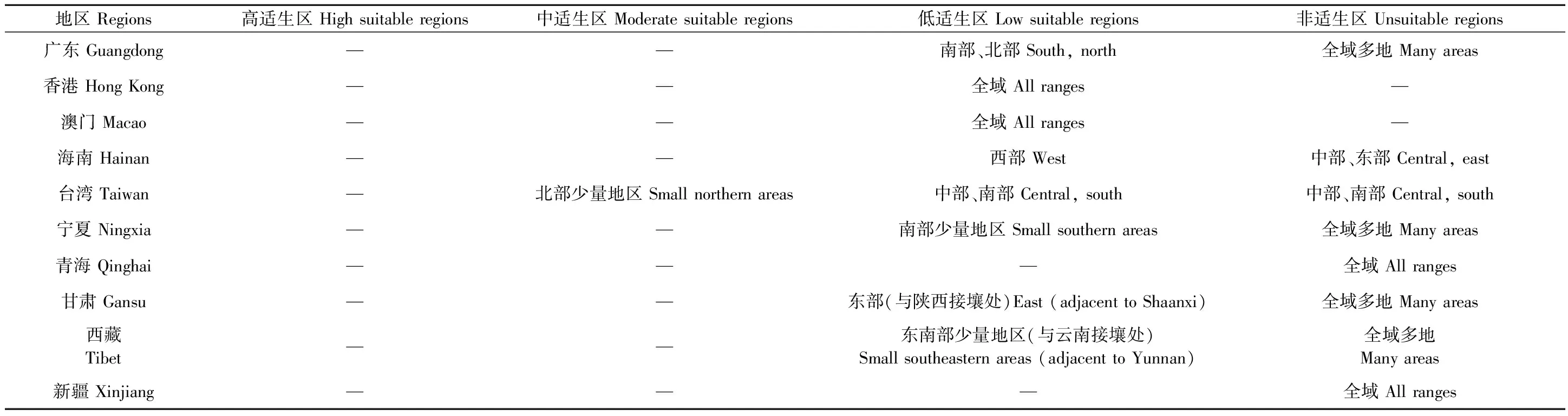
续表3 Continued

表4 仅考虑寄主分布的栗山天牛中国适生区Tab.4 Potential distribution of M. raddei based on host distribution in China

续表4 Continued

表5 同时考虑寄主和气候因素的栗山天牛中国适生区Tab.5 Potential distribution of M. raddei based on the combination of climatic factors and host distribution in China

续表5 Continued
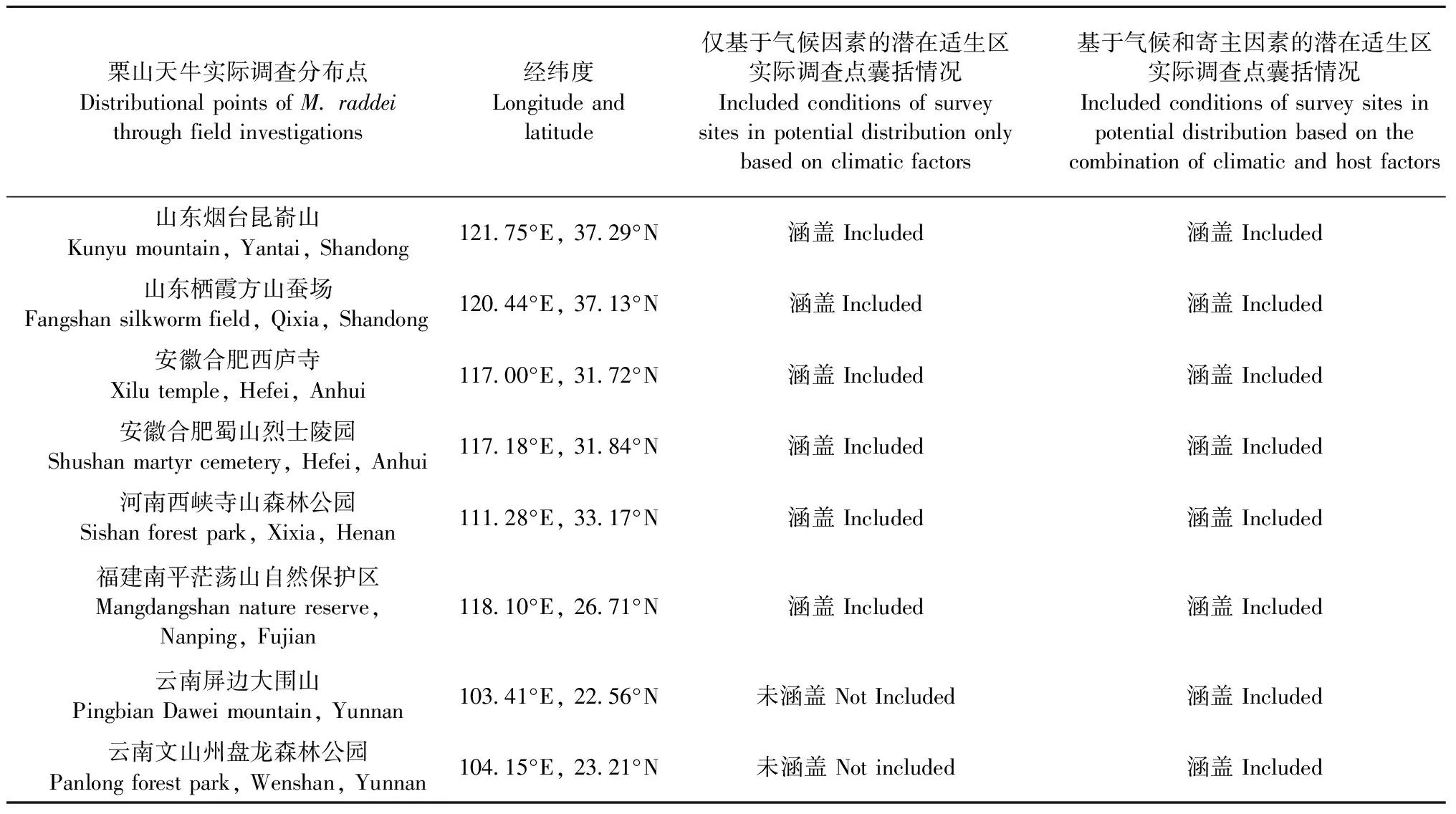
表6 栗山天牛实际调查分布点和模型准确性评估结果Tab.6 Distributional locations of M. raddei obtained from field surveys and the model accuracy assessment results
3 讨论
本研究基于寄主和气候因素对栗山天牛在中国不同地理区域的扩散和成灾风险进行了评估。敏感树种蒙古栎和辽东栎的广泛分布是东北林区为栗山天牛高适生区的主要原因,也是导致该虫在此地暴发成灾的生态因素之一。本研究还发现即使在气候不适宜的地区,一旦分布有寄主树种,栗山天牛便可在此地定殖发展。因此,在该虫暴发成灾的东北林区里,以非寄主或次要取食寄主为主的混交林取代蒙古栎和辽东栎纯林可作为长期生态防控策略; 在该虫的未分布省份和区域,应严格管控栎树相关的木材贸易,以防我国栗山天牛发生和成灾范围的进一步扩大。
3.1 生态因子分析
影响栗山天牛潜在适生区的主导气候因素(贡献率超过10%)依次为最湿月降雨量、年平均雨量、年平均温度和等温性。由于幼虫在树体内蛀食,只要外部环境因素可满足成虫羽化和繁殖活动,便有利于栗山天牛定殖。温湿度对于栗山天牛成虫羽化和后续的繁殖行为均很重要,22~26 ℃间的林间温度和50%~80%的林间湿度将有利于成虫活动,但过高的湿度将导致成虫羽化数量减少(唐艳龙,2011; 杨忠岐等,2011)。此外,温湿度也决定着寄主树种的土壤水源供给、生长发育和生产力提高(殷晓洁等,2013; 李腾等,2014),将间接影响栗山天牛的分布情况。本研究也发现栗山天牛潜在分布概率随最湿月份降雨量、年平均雨量和年平均温度的增加表现为先上升后下降的趋势。此外,等温性对栗山天牛潜在分布概率的响应曲线表现为下降、上升再下降的趋势,西藏、青海和新疆的过高海拔将会导致较大的昼夜温差,也是这几个省份在气候上表现为栗山天牛非适生区的主要原因。然而,王志刚等(2003)报道光肩星天牛(Anoplophoraglabripennis)可以在西藏南部海拔3 000 m以上地点定殖,因此考虑到蛀干害虫的隐蔽生活习性,栗山天牛扩散到上述高海拔地区后也存在定殖并成灾的可能性。
3.2 栗山天牛成灾和扩散风险评估
栗山天牛东北林区的局部成灾除与羽化同步性导致的成虫高种群密度有关外(Williamsetal.,1995; 王小艺等,2012; Zhangetal., 2020),本文提出高适寄主和适宜气候也有助于种群发展和扩张。相似地,Morewood等(2004)和Faccoli等(2016)均发现光肩星天牛在敏感树种上生存时表现出更高的产卵量和幼虫存活率,认为光肩星天牛的抗性寄主栾树(Koelreuteriapaniculata)和豆梨(Pyruscalleryana)分别依靠物理防御(树体流胶)和化学防御(树体内次生化合物)阻碍该天牛幼期生长,进而抑制天牛种群数量增长和进一步成灾; 亚洲本土白蜡(Fraxinusspp.)树种对白蜡窄吉丁(Agrilusplanipennis)的抗性普遍比北美栽培树种的抗性强,这也是该虫在原产地密度较低不成灾,而在北美入侵地快速成灾的原因之一(Rebeketal., 2008; 路纪芳等,2012)。相似地,栗山天牛在我国南部省份每年均有成虫羽化,气候条件也适宜该虫种群定殖发展,然而栗山天牛对南部地区主要分布的板栗、青冈栎寄主种类表现出次要喜食程度和低适生性,骆有庆等(2005)曾报道我国南部省份的栗山天牛种群密度比北部种群的更低,不成灾。在本研究中,同时考虑寄主和气候因素获得的栗山天牛中国适生区相对于仅依据寄主分布或仅依据气候条件的栗山天牛中国适生区表现为高适生区面积的缩小和低、中度适生区面积的增加,说明各地的现有寄主适生性等级是限制栗山天牛分布和成灾的主导因素之一。因此,完善成灾区林分结构,以非寄主树种或非偏嗜取食寄主为主的混交林取代蒙古栎和辽东栎纯林可作为成灾林区长期生态防控手段。此外,在栗山天牛分布的非成灾林区内,应该避免因人为种植或定期间伐而产生大面积的易感寄主蒙古栎和辽东栎纯林,以防加速该虫的蔓延危害和进一步的暴发成灾。
寄主树种对于物种分布特别是生活隐蔽、受环境条件波动性小的蛀干害虫的分布发挥着关键作用(Fletal., 2015; Shimetal., 2020; Dangetal., 2021)。在气候条件可能不适宜栗山天牛生长发育的地区,当有适宜树种分布时,栗山天牛也可能定殖,云南文山州和屏边大围山实际调查分布点有力佐证了这一点。相似地,Dang等(2021)报道了敏感白蜡树种对白蜡窄吉丁潜在分布区的重要影响,即使在依据气候因素预测的不适生区域内,只要存在适宜的寄主树种,也可能发现白蜡窄吉丁的发生和危害。Lee等(2020)发现了光肩星天牛在韩国本土分布地二次扩散蔓延至韩国其他未分布区域的生态现象。类似地,栗山天牛虽仅在我国东部诸多省份广泛分布,但仍存在于内蒙古、宁夏、青海、西藏、新疆、贵州、广东、甘肃、山西、云南、广西、四川、海南等未报道分布或者仅存在少数分布点的地区,且上述多地表现为在气候上对栗山天牛定殖的适宜性,因而该虫同样存在向这些地区蔓延定殖的可能。近些年来,上述地区的苗木调运、植树造林等人为活动已经导致多种蛀干害虫的扩散和定殖,威胁当地的生态安全,造成较大的经济损失(韩玺等,1995; 骆有庆等,1999; 王志刚等,2003; 才旺计美等,2007; 高瑞桐等,2009; 岳朝阳等,2011; 阿里木等,2013)。同时,本研究中的寄主分布信息显示在西藏林芝市已经有麻栎分布,在内蒙古、甘肃、山西和宁夏多地分布有易感树种蒙古栎和辽东栎,青海和新疆虽然未发现有寄主树种分布,也仍然存在该虫由周边分布省份随原木或木质包装材料传播从而定殖并危害转主寄主的可能性。Williams等(1995)指出入侵未分布地区的昆虫可能表现出过渡周期羽化行为(即在昆虫生命周期内,其中1年的虫口密度明显大于其余年间),或者进一步的周期性羽化行为。因此,栗山天牛若扩散至我国未分布地后,将可能产生类似东北林区的成虫同步性羽化并暴发成灾的现象。总之,未分布地的早期检疫和预警措施是必要的。
3.3 预测模型可行性评估
本研究结果显示基于寄主和气候因素所获得的栗山天牛潜在适生区准确度高于由单一因素获得的预测结果,适生区面积囊括了所有实际调查分布点,且与现阶段该虫不同地理种群危害等级更吻合。此外,本研究中假设寄主和气候因素对该虫的潜在分布概率的贡献率是相同的,即均为50%,从试验结果来看,这2大因素对该虫分布概率为叠加影响效应的假设是正确的,但是两者贡献率是均为50%还是数值一大一小仍有待进一步确定。总之,在对栗山天牛这类营隐蔽方式生活的蛀干害虫进行潜在适生区预测中,综合考虑气候适宜性和寄生植物可利用性是必要的。Shim等(2020)基于寄主和气候因素预测了光肩星天牛在韩国的潜在适生区,其结果相对仅依据气候因素的预测分布区更加符合光肩星天牛在本地的实际危害情况。不同的是,该研究认为在气候不适宜或未分布适生寄主的地区(即相应预测结果的潜在分布概率为0),害虫将不会在此处分布定殖,基于本文结果,认为此结论是过于绝对的,其忽视了适生性寄主植物为蛀干害虫提供抵御不适生长环境的营养物质价值,以及转主寄主危害的可能性。因此,本文也提出一种针对栗山天牛在内的蛀干害虫扩散和成灾风险评估的新方法,可更精确地筛选出害虫易成灾地区,以便采取针对性的检疫和预测预报措施。
4 结论
1)偏嗜寄主植物蒙古栎及辽东栎和气候的高适生性,是造成栗山天牛在东北林区表现为高度适生并能局部成灾的重要生态因素,同时该虫在我国的未分布地区内均存在扩散和定殖风险。以非寄主树种或者非偏嗜寄主为主的混交林逐渐取代蒙古栎和辽东栎纯林,可作为成灾林区的长期生态防控策略。此外,建议相关地区的造林绿化和植物检疫部门予以重视,加强监管和早期预警措施,以防栗山天牛在我国发生和成灾面积进一步的扩大。
2)在栗山天牛成灾和扩散风险评估中应综合考虑气候适应性和寄主植物可利用性,这种新型的预测方法精确度更高,有助于筛选出易成灾区并及时采取早期预防措施。
猜你喜欢
风景园林(2022年1期)2022-03-07
意林·少年版(2020年17期)2020-10-12
发明与创新·中学生(2020年4期)2020-04-17
农民致富之友(2019年11期)2019-05-23
幼儿教育·父母孩子版(2018年8期)2018-12-28
故事作文·低年级(2018年10期)2018-10-25
少儿科学周刊·儿童版(2018年12期)2018-01-26
少儿科学周刊·少年版(2018年12期)2018-01-26
少儿科学周刊·儿童版(2017年7期)2017-09-29
作文与考试·小学低年级版(2015年11期)2015-07-17
