
广西家蚕僵病流行趋势及其主要病原遗传规律研究
2022-10-11 12:51唐亮胡文娟蒋满贵黄深惠董桂清李标唐名艳赵烨芸何骥潘志新
南方农业学报 2022年7期
唐亮,胡文娟,蒋满贵,黄深惠,董桂清,李标,唐名艳,赵烨芸,何骥,潘志新
(广西蚕业技术推广站/广西蚕业科学研究院,广西南宁530007)
0 引言
【研究意义】蚕病是在家蚕饲养环节中制约蚕茧产量、质量及蚕农经济效益的主要因素之一(祁广军,2013)。其中,僵病是养蚕过程中全龄易发的真菌蚕病,且病源多样,常经风媒传播或直接接触蚕体而感染致病,若防控不及时极易暴发流行,给蚕桑产业造成重大经济损失(廖华珍等,2009;唐亮等,2013)。因此,掌握家蚕僵病的发病规律及病原特性,建立科学的防控策略和措施,对有效降低家蚕僵病危害、保障蚕业稳定发展及促进蚕农增收均具有重要意义。【前人研究进展】牟志美等(1999)研究报道,常规年份山东家蚕的僵病危害在5%~8%,与饲养品种、发育时期及野外昆虫的发生密切相关;朱方容(2008)、廖华珍等(2009)调查发现广西2008—2009年连续2年春季家蚕僵病暴发流行,造成当地蚕茧产量锐减,其主要病原可能来源于飞机喷施森林防虫用的白僵菌。引起家蚕僵病的病原除白僵菌属外,还有拟青霉属(朱方容等,1997;苘娜娜等,2015)、曲霉属、莱氏野村菌属(王长军,2000)及蜡蚧属(苘娜娜等,2015)等多种真菌病原,存在持续的感染能力(余金勇等,2007),但不同菌株间对宿主的毒力存在明显差异(Saito and Brownbridge,2016)。Wang等(2005)利用简单序列重复(ISSR)分子标记研究39个白僵菌属分离株的遗传多样性,结果表明ISSR分子标记能成功将3个密切相关的物种与球孢白僵菌()区分开来,并证实球孢白僵菌分离株与其地理来源存在一定关联,但这些分离株与其昆虫宿主间无明确的相关性。吕思行等(2012)采用微卫星分子标记对广东和广西蚕区的白僵菌居群之间和居群之内的遗传多态性进行分析,结果发现两广白僵菌居群之间的基因分化系数()为0.0590,多态位点百分率(PPL)为97.73%,Nei's基因多样性指数()为0.1896,Shannon信息多样性指数()为0.3165,表明两广家蚕来源的白僵菌居群间遗传分化较小。唐亮等(2013)通过研究宿主湿度环境对球孢白僵菌致病力的影响,发现球孢白僵菌在病蚕体中宿存60 d内对家蚕的致死率仍然很高,因此及时清除病蚕尸体并进行尸体病原性灭活,可降低白僵菌的二次感染。陈雪(2014)通过ISSR遗传多样性数据及菌株对家蚕的致死数据研究全国各地球孢白僵菌种群关系,结果表明家蚕白僵病的发生与松毛虫白僵病的自然流行或人工释放球孢白僵菌杀虫剂均无直接关系。王婷婷(2019)在柞蚕僵病病原分子鉴定的研究中发现,野外柞蚕僵蚕、僵蛹及室内人工接种僵蚕的内转录间隔区(ITS)序列相似性存在明显差异,与已知基因序列(KM205065.1)的相似性在96.01%~100.00%。王露露等(2022)从山东和广西僵蚕及生防菌中筛选获得对家蚕具有高毒力的白僵菌,并优化其发酵条件,为制备僵蚕提供了优质的菌种资源。Wang等(2022)通过对比、、、、及共6个基因序列,以评估中国、泰国15株白僵菌的系统发育位置,结果筛选出3株具有生防潜力的白僵菌株。【本研究切入点】查明广西家蚕僵病发生状况、流行规律,掌握病原特性及其遗传规律等是有效防控该病的基础,但目前尚缺乏对广西家蚕僵病的系统性研究。【拟解决的关键问题】通过分析2008—2021年广西主要蚕区蚕病调查数据,结合病原菌形态学与分子生物学研究,旨在掌握广西主要蚕区的僵病流行规律及其病原特性,为精准防控家蚕僵病提供科学依据。
1 材料与方法
1.1 蚕病数据
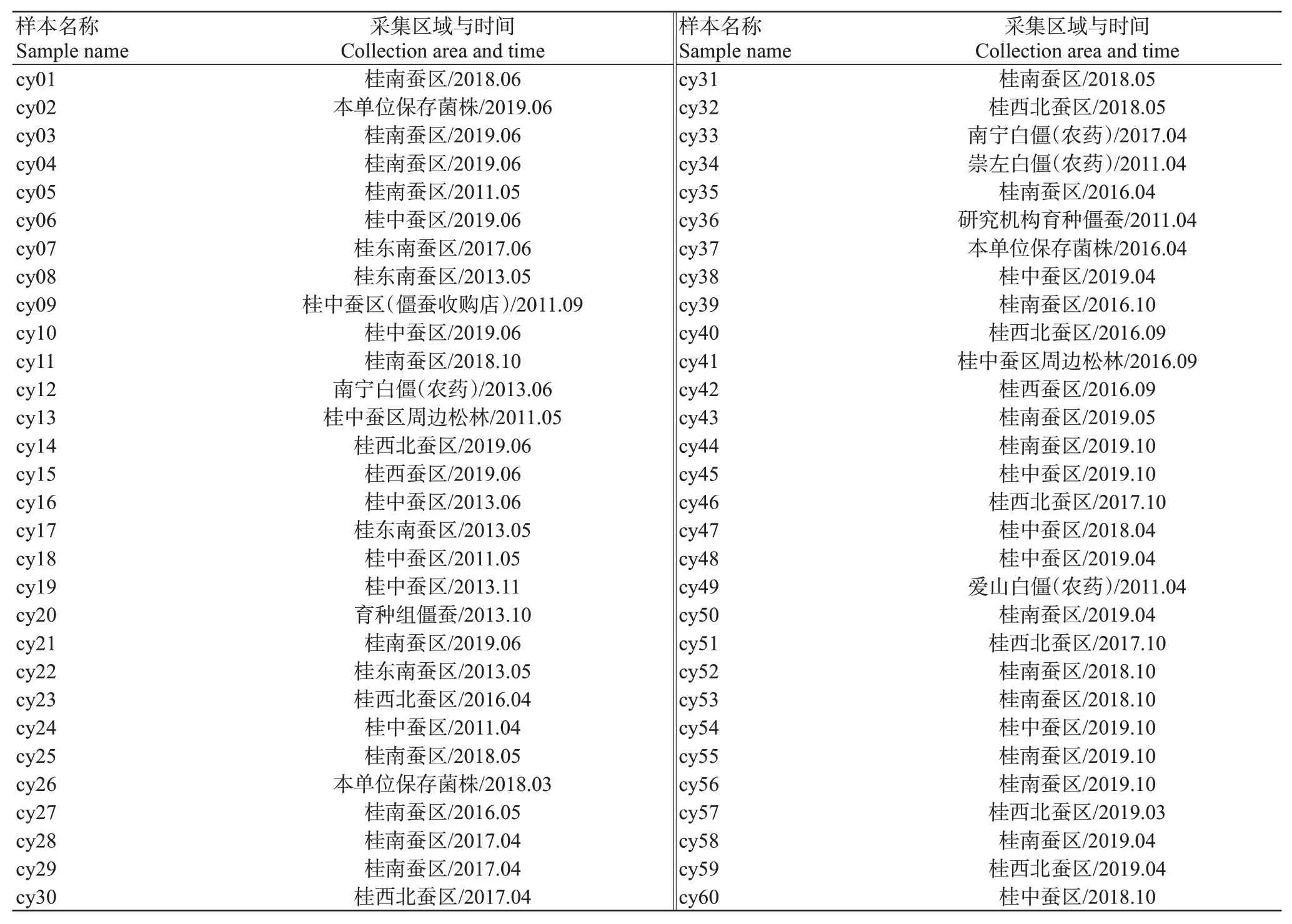
广西家蚕僵病发病原始数据由广西蚕桑病虫害监测网的常规蚕病监测数据和相关科技项目蚕病调查数据组成;数据汇总年份为2008—2021年,月份为每年3—10月。每批次全龄期蚕病调查数据为三龄起蚕、五龄起蚕、上蔟后第2 d的僵病发生率总和。
1.2 供试病原
家蚕僵病病原采自桂中、桂南、桂西、桂西北、桂东南等广西主要桑蚕主产区的僵蚕,清洁处理后接种于萨氏培养基上,划线分离纯化获得单菌落,经显微形态鉴定和产孢测定后,保存于广西蚕业技术推广站蚕桑病虫害研究室菌种库。
1.3 供试菌丝及分生孢子制备
将供试菌株接种至萨氏培养基,培养箱(25±1)℃倒置培养10 d左右,待大量产生菌丝和分生孢子后,刮取表层菌丝及分生孢子,经液氮速冻后,分别置于-20和-80℃冰箱保存备用。
1.4 病原菌对家蚕致病力测试
采用滤纸接触法(唐亮等,2013)添毒两广二号的二龄起蚕,置于(25±1)℃、相对湿度在90%以上的环境下饲养,连续观察7 d内家蚕发病情况。重复3次。
1.5 病原菌DNA提取
参照朱衡等(1994)的方法提取病原菌总DNA,以1.5%琼脂糖凝胶电泳检测DNA质量;采用核酸测定仪测定DNA浓度后稀释至20 ng/μL,-20℃保存备用。
1.6 病原菌ITS序列分析
采用真菌通用引物ITS4(5'-TCCTCCGCTTAT TGATATGCITS-3')/ITS5(5'-GGAAGTAAAAGTCG TAACAAGG-3')组合进行PCR扩增(陈立杰等,2008),PCR反应体系50.0μL:DNA模板2.0μL,dNTP Mixture(2.5 mmol/L)2.5μL,ITS4/ITS5引物(20μmol/L)各1.5μL,10×ExBuffer(MgPlus)5.0μL,Ex酶(5 U/μL)0.2μL,ddHO补足 至50.0μL。扩增程序:94℃预变性3 min;94℃1 min,55℃1 min,72℃2 min,进行30个循环;72℃延伸5 min。PCR扩增产物采用1.5%琼脂糖凝胶电泳进行检测,回收目的条带,经PCR Purification Kit纯化后以扩增引物进行双向测序,所得测序结果拼接后输入GenBank,通过BLAST进行比对分析。
1.7 病原菌ISSR多态性分析
从测试样本中选取4个样本对20对引物进行测试,筛选出10个扩增条带清晰、多态性好的引物用于ISSR-PCR反应。反应体系25.0μL:DNA模板约40 ng,10×Bufer 2.5μL,DNA聚合酶1.25 U,引物(10 ng/μL)1.0μL,dNTPs(2.5 mmol/L)0.4μL,MgCl(1.5 mmol/L)2.0μL,ddHO补足至25.0μL。扩增程序:94℃预变性5 min;94℃45 s,55~62℃45 s,72℃1.5 min,进行35个循环;72℃延伸10 min,4℃保存。PCR扩增产物以2.0%琼脂糖凝胶电泳进行检测。
1.8 统计分析
基于ITS4/ITS5引物进行PCR扩增,并将双向测序结果输入GenBank进行BLAST比对分析。ISSRPCR扩增产物在琼脂糖凝胶电泳的某个相同迁移率位置上有DNA条带记为1,无DNA条带则记为0,形成ISSR表型数据矩阵,然后使用NTsys计算遗传相似系数,并基于遗传相似系数进行UPGMA聚类分析。同时,使用PopGene32计算供试材料的遗传多样性指数(安丹丹等,2021;周春宝等,2021),并以SPSS 25.0进行单因素方差分析(One-way ANOVA)。
2 结果与分析
2.1 不同年份家蚕僵病的发生规律
2008—2021年广西主要蚕区的家蚕僵病平均发病情况如图1所示。广西家蚕僵病发病率整体上呈波动下降趋势,连续14年的平均发病率为5.19%。其中,2008年的发病率为10.08%,显著高于其他年份(<0.05,下同);发病率最低的年份是2019年,发病率仅为3.16%。此外,广西家蚕僵病年均发病率曲线在2012年呈现一个转折点,2008—2012年整体上呈快速下降趋势,2012—2021年则现出上下波动,但总体维持在较低水平,年均发病率仅为4.08%。
2.2 不同月份家蚕僵病的发生规律
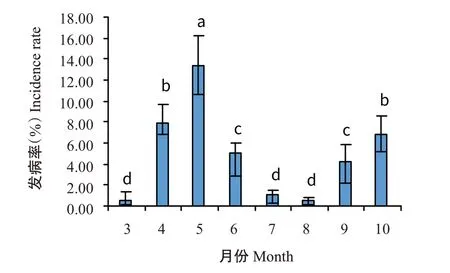
以月为单元进行分析,结果显示广西家蚕僵病以5月的发病率最高(图2),达13.37%,与其他月份的差异均达显著水平;次高峰出现在4月和10月,对应的发病率分别为7.91%和6.87%,二者差异不显著(>0.05,下同)。广西家蚕僵病发病高峰和次高峰时段与广西温润多雨气候存在高度重叠,推测温湿度对家蚕僵病的发生有较大影响。
2.3 不同蚕区不同地形家蚕僵病的发生规律

据2008—2021年桂中、桂南、桂西、桂西北和桂东南五大蚕区的调查数据显示,各蚕区平缓丘陵地形、山区地形的家蚕僵病发生情况如图3所示。由图3可知,山区地形的家蚕僵病发生率普遍高于平缓丘陵地区。平缓丘陵蚕区的家蚕僵病发病率排序为桂中(13.55%)>桂西北(12.66%)>桂南(11.56%)>桂西(11.05%)>桂东南(9.53%),可能与桂中、桂西北在春季低温阴雨天气偏多,湿度偏高有利于真菌萌发有关。值得注意的是,5月是各蚕区家蚕僵病的发病高峰期,但进入6月中下旬家蚕僵病发生率迅速下降,可能是平缓丘陵地区气温上升迅速、湿度快速下降,超出了真菌发育的最适温湿度而表现出家蚕僵病发病率急剧下降。
山地蚕区的家蚕僵病发病率排序为桂中(18.91%)>桂西北(17.96%)>桂西(16.01%)>桂西(14.90%)>桂东南(11.61%),可能是桂中、桂西北山区养蚕多位于河谷地区,相对日照较短,下午16:00后的太阳照射盲区及夜间会出现局部相对湿度偏高而有利于真菌萌发和感染传播。相对于平缓丘陵地区5月家蚕僵病发病高峰过后次月发病率骤降的情况,山地蚕区的家蚕僵病发病率下降趋势略显缓慢,尤其是桂中、桂西、桂西北地区6月的家蚕僵病发生率仍维持在7.80%~10.28%,提示山地蚕区的家蚕僵病危害时间更严重且更持久。

2.4 病原菌形态学观察及其对家蚕毒力测试结果
本研究收集了桂中、桂南、桂西、桂西北及桂东南5个蚕桑主产区多年发生的僵蚕、僵虫,以及部分白僵菌(农药),以此为基础分离获得60株病原单菌落(表1)用于后续测试。分离获得的单菌落接种至萨氏培养基上,26℃培养14 d,连续观察其生长情况并统计菌落的产孢情况,综合评价病原菌的危害。然后挑取菌丝和分生孢子制片,在显微镜下观察分生孢子形状、分生孢子梗分支情况及产孢方式等真菌分类特征,初步鉴定分离菌株即为白僵菌(图4)。
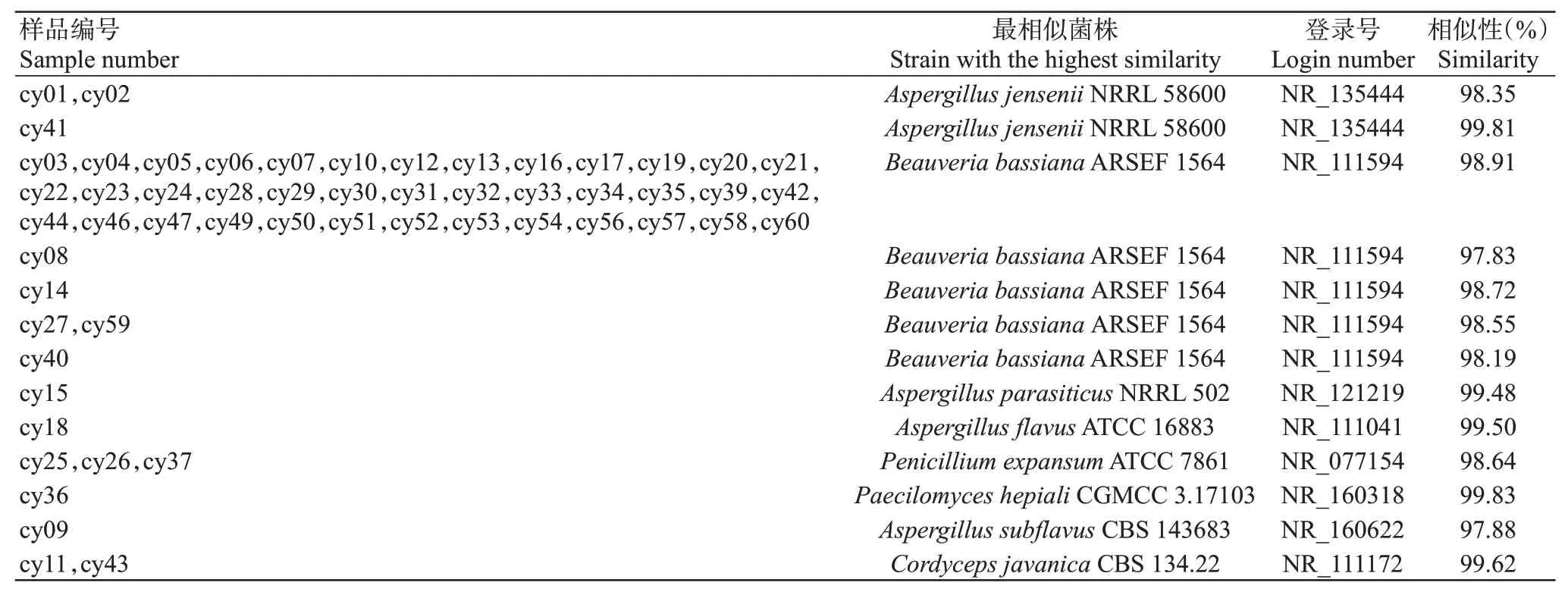
筛选广西各蚕区及不同来源农药白僵菌株产孢量第一或前两名的菌株,统计分析其中的10株菌株(表2),产孢量排名前3的菌株是崇左白僵菌株(农药)、爱山白僵菌株(农药)和桂南蚕区白僵菌株,对应的平均产孢量分别为3.4×10、2.8×10和2.6×10分生孢子/mm,其中前两者是森林防虫用农药白僵菌,而桂南蚕区白僵菌是从白僵蚕分离获得,说明农药源白僵菌在产孢量方面具有明显优势。其中,桂南蚕区白僵菌株来源于广西蚕业技术推广站品种保育过程的僵蚕,由于广西蚕业技术推广站地处南宁市区,养蚕环境独立、饲养技术规范,与其他蚕区病原基本无交集,故推测该菌株是1株相对独立的野生白僵菌,但其产孢量在所有测试菌株中最低。
同时测试上述10株白僵菌株对家蚕的致病力,结果(图5)显示,这些白僵菌株对两广二号二龄幼虫的致死速率存在一定差异,但7 d累计致死率均达100.00%,致病力最强的为崇左白僵菌株(农药),其攻毒48 h后即出现家蚕吐水现象,72 h累计致死率达100.00%。72 h累计致死率排名第一梯队为3种农药源白僵菌株:崇左白僵菌株(农药)(cy34)100.00%、南宁白僵菌株(农药)(cy33)98.89%、爱山白僵菌株(农药)(cy49)97.78%;其次是蚕区病蚕分离的白僵菌株和松林僵虫白僵菌株:桂中松林白僵菌株(cy13)94.44%、桂南2白僵菌株(cy28)93.33%、桂中2白僵菌株(cy16)93.33%、桂西北白僵菌株(cy51)92.22%、桂中1白僵菌株(cy24)91.11%、桂东南白僵菌株(cy22)83.33%,即蚕区病蚕白僵菌株致死率在83.33%~93.33%,低于农药源白僵菌株对家蚕的致死率;最后为独立的桂南1白僵菌株(cy20)8.89%,明显低于农药用白僵菌株。经显著差异分析发现,除爱山白僵菌株(农药)与桂中松林白僵菌株、桂南2白僵菌株、桂中1白僵菌株、桂中2白僵菌株及桂西北白僵菌株的差异不显著外,南宁白僵菌株(农药)、崇左白僵菌株(农药)均显著高于蚕区白僵菌株的致病力,松林白僵菌株显著高于桂东南蚕区和桂南独立蚕区的白僵菌株。即10株白僵菌株对家蚕的致病力总体上呈现出农药源白僵菌株>松林白僵菌株>蚕区病蚕白僵菌株的趋势,同时存在桂南、桂中、桂西北蚕区白僵菌株感染力高于桂东南、桂西蚕区的白僵菌株。可见,桂南、桂中、桂西北蚕区受白僵菌农药影响的可能性较大,即蚕桑主产区应慎用白僵菌农药。



2.5 病原菌ITS序列分析结果
以最初冻存的病原菌丝和分生孢子为样本,提取60株病原单菌落总DNA,经1.5%琼脂糖凝胶电泳检测合格(图4-A)后,利用ITS4/ITS5引物进行目的片段扩增,PCR扩增电泳结果(图4-B)显示60株病原样本均能扩增获得预期大小的目的条带,可用于后续的ITS序列分析。
将PCR扩增产物送至生工生物工程(上海)股份有限公司进行双向测序,测序结果输入GenBank进行BLAST比对分析,结果显示扩增的ITS序列存在高度保守性,有多个序列比对结果完全一致。在60株病原菌中有56条序列匹配到8个已知序列,序列相似性在97.83%~99.83%(表3)。其中,来自桂中蚕区2019年的3个样本和桂南蚕区2019年的1个样本,经2次重复提取DNA均未能成功测序,故不计入后续分析。在56株匹配的菌株中,有45株为蚕区采集病原株,且其中39株为球孢白僵菌()。可见,导致蚕区家蚕白僵病的主要病原为球孢白僵菌,占比达86.67%;诱发家蚕僵病的病原还有曲霉属、拟青霉属和虫草菌属等,表明蚕区家蚕僵病病原复杂多样。
2.6 白僵菌遗传多样性及聚类分析结果
在ITS序列分析的基础上,选择45株蚕区白僵菌株样本、4株农药源白僵菌株样本及1株松林白僵菌株样本共计50株菌株进行多态性分析。首先选用cy03、cy14、cy40、cy60等4个样本进行ISSR引物筛选,结果从20条引物筛选出10条扩增条带清晰、条带数较多、多态性良好,且易于扩增的引物(表4)。利用筛选出的10条ISSR引物对50株菌株样本的DNA进行ISSR-PCR扩增,扩增产物以2.0%琼脂糖凝胶电泳进行检测,并根据ISSR-PCR扩增结果以Quantity One绘制电泳模式图,结果显示,10条ISSR引物从50株菌株样本中扩增出152条清晰条带(表5),其中多态位点147个,多态百分数为87.5%~100.0%,平均为96.71%。各引物扩增获得的条带数在13~19条,平均为15.2条。从单条引物扩增得到的多态位点来看,以引物P8和引物P9扩增得到的多态位点最多,均为19个。10条引物扩增得到的多态位点平均为14.7个,表明这10条引物能很好地区分白僵菌株多样性。利用PopGene32计算供试材料的遗传多样性指数,结果显示,50株菌株样本的等位基因数()为1.8750~2.0000,有效等位基因数()为1.2726~1.5625,Nei's基因多样性指数()为0.1740~0.3257,Shannon信息多样性指数()为0.2840~0.4880,均表现出极高的遗传多样性,尤其以引物TL-P8的扩增效果最优(图7)。

根据“0,1”矩阵数据,分别计算50个供试材料两两间的遗传距离并构建遗传相似矩阵,结果显示,cy13与cy16的遗传相似系数(GS)最大,为0.9737,表明二者的遗传距离非常近;而cy33(白僵属)与cy25(拟青霉属)和cy26(拟青霉属)的GS最小,均为0.4276,表明其遗传距离较远。蚕区来源白僵菌株间的GS波动幅度在0.5921~0.9671,说明蚕区病原遗传多样性较高,同时中位区间在0.8000~0.8999,占56.41%,预示着超过半数的蚕区病原基因相关度较高,GS大于0.8000的比例高达78.00%,则提示区域内的菌株变异较小或区域间存在频繁交流。其中,蚕区病原的GS最高值(0.9737)出现在2013年的2株桂中蚕区病原菌株间(cy16与cy19),最小值(0.5921)则出现在2013年育种僵蚕菌株cy20与同年桂西北蚕区的僵蚕病原菌株cy23间,表明相同地域的白僵菌株会表现出更高的亲缘关系,而跨区域菌株间的基因相关度较低。



根据50株菌株两两间的遗传距离进行UPGMA聚类分析,结果(图8)显示,蚕区病原菌株与农药源白僵菌株聚类在相邻位置,提示二者具有较紧密的遗传关系。如cy28(桂南蚕区,2017年)与cy33[南宁白僵(农药),2017年]、cy51(桂西北蚕区,2011年)与cy49[爱山白僵(农药),2011年]、cy22(桂东南蚕区,2013年)与cy12[南宁白僵(农药),2013年]、cy24(桂中蚕区,2011年)与cy34[崇左白僵(农药),2011年]间的遗传距离分别为0.9276、0.9342、0.9605和0.9568,预示着桂南、桂西北、桂东南、桂中等蚕区的病蚕白僵菌株与农药源白僵菌株存在非常近的亲缘关系,进一步证实广西各蚕区白僵病的发生流行与施用白僵菌农药存在密切关系。
3 讨论
2008—2021年广西主要蚕区的家蚕僵病平均发病率为5.19%,与牟志美等(1999)研究报道山东家蚕僵病常规年份的年均发病率在5%~8%相仿。广西属于高温多湿的亚热带季风气候,山东属于温带季风气候,两地温湿度差异明显,但两地的家蚕僵病发病率相近可能与广西气温偏高,适宜白僵菌繁殖和浸染的时间范围较窄有关。在2008和2009年广西蚕区周边环境引入大量森林防控白僵菌(朱方容,2008;廖华珍等,2009)的情形下仍取得良好防控效果,预示着在没有大量外源病原菌影响的情况下僵病发病率可能会更低。自2009年广西蚕区大面积引入外源性白僵菌后,2010和2011年广西蚕区周边未见大量施用白僵菌的报道,但2010年的家蚕僵病发病率显著高于之后的多年平均发病率(4.08%),广西家蚕僵病年均发病率曲线在2012年呈现一个转折点。说明人为引入大量白僵菌株后,对当地家蚕僵病的影响可能会维持2年左右,第3年则呈显著下降趋势,与余金勇等(2007)在研究白僵菌防松毛虫持续致病力得出的结论相似。本研究通过对广西主要蚕区及周边地区收集的白僵菌株进行ITS序列分析,进一步明确广西家蚕僵蚕的病原种类,包括白僵菌属、曲霉属、拟青霉属和虫草菌属,表明广西家蚕僵病病原呈多样性。其中,拟青霉属病原与朱方容等(1997)的研究结果相似。广西家蚕僵病的主要病原仍是球孢白僵菌(占86.67%),说明广西蚕区家蚕僵病的病原高度保守。

本研究对广西五大蚕桑主产区家蚕僵病病原、4株生物防控白僵菌(农药)、1株松林白僵分离株和1株研究机构育种白僵菌进行毒力测试,结果显示生物防控白僵菌株>松林白僵菌株>主要蚕区白僵菌株>研究机构育种白僵菌。生物防控白僵菌株(农药)对家蚕的致病力显著高于蚕区白僵菌株,提示其具备区域性危害的基础条件,与石美宁和朱方容(2010)、刘吉平等(2011)的研究结果相似。本研究同时利用ISSR-PCR分析广西主要蚕区白僵菌病原间的基因相关度,结果显示同一地域相同年份的病原菌株具有非常近的遗传距离,如cy16与cy19的GS最大(0.9737);相同区域经过多年后遗传变异较大,如cy28与cy58的GS最小(0.7895)。广西桑蚕主产区家蚕僵病病原的GS中位数区间在0.8000~0.9000,约有78.00%的GS大于0.8000,暗示广西白僵菌居群的遗传多样性水平相对较低,与吕思行等(2012)的研究结果相似。此外,通过对比生物防治用白僵菌株(农药)与广西部分蚕区白僵菌株,发现其遗传距离非常近,同时蚕区周边松林分离白僵株与前两者存在极高的遗传关联度,预示着松林害虫可能是白僵菌(农药)感染家蚕的中间宿主(陈雪,2014),即广西蚕区白僵病的流行与施用白僵菌农药存在密切联系。
4 结论
广西家蚕僵病危害高峰期为5月,球孢白僵菌仍是最主要的病原菌,且ITS序列高度保守。不同来源白僵菌对家蚕致病力的排序为农药源白僵菌株>松林白僵菌株>蚕区僵蚕菌株,但三者相互间存在非常高的基因关联性,提示广西家蚕僵病的病原遗传背景复杂,且蚕桑主产区应慎用白僵菌农药。
猜你喜欢
广西植物(2022年8期)2022-09-07
健康体检与管理(2022年2期)2022-04-15
科学导报(2020年32期)2020-05-26
中学生物学(2019年7期)2019-10-17
中学生物学(2017年7期)2017-08-23
科技资讯(2016年32期)2017-03-31
现代农业科技(2017年1期)2017-03-06
现代园艺(2016年9期)2016-07-16
江苏农业科学(2015年1期)2015-04-17
