
一株新型重配H9N8亚型禽流感病毒的分子进化分析及其对小鼠的感染性研究
2022-10-06 05:53崔鹏飞谷文丽张元成衡武昌王丛丛邓国华陈化兰
中国预防兽医学报 2022年7期
颜 成,崔鹏飞,邢 鑫,谷文丽,张元成,衡武昌,王丛丛,邓国华,陈化兰
(中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室/农业农村部动物流感重点开放实验室,黑龙江 哈尔滨 150069)
自1966 年在美国的火鸡中首次报道以来[1],H9亚型禽流感病毒(Avian influenza virus,AIV)已在世界范围内呈地方性流行,特别是在亚洲、中东以及北非和中非部分地区的家禽中广泛流行[2],并且H9亚型AIV 在自然界的野生鸟类中也广泛分布。1994年,在我国广东鸡群中首次分离到H9N2 亚型AIV,随后H9 亚型AIV 在我国多地出现,呈现蔓延趋势,目前已成为我国鸡群中流行的最主要亚型AIV[3-4]。当前,H9 亚型AIV 的HA 基因可分为7 个进化分支,其中G1-like 和BJ94-like 是我国流行的主要H9 亚型AIV[2]。H9 亚型AIV 感染可引起家禽产蛋量下降,当与细菌混合感染时常致家禽较高的致死率,给家禽养殖业造成了重大经济损失[5]。H9 亚型AIV 也可以跨越种间屏障感染人类,1998 年郭元吉等首次在广东省流感患者咽喉样品中分离到H9 亚型AIV[6],截止2019 年7 月,全球已有59 例人感染H9N2 亚型AIV病例[7]。此外,血清学数据表明在部分地区的职业暴露人群中,H9N2 特异性抗体的血清检测率超过20%[8-9]。
H9 亚型AIV 有多种NA 亚 型组合,其中H9N2 是最常见的亚型,约有98%的H9 亚型AIV 属于H9N2亚型。到目前为止,H9N8 亚型AIV 出现的频率极低,GenBank 中仅查询到8 条数据。2020 年5 月,本研究在广西某鸭场中分离到一株H9N8 亚型AIV,为了解该病毒的进化起源,本研究对其进行了分子进化分析,并以BALB/c 小鼠为实验动物模型评价了其对哺乳动物的感染性,为H9 亚型AIV 的流行病学调查提供数据支持。
1 材料与方法
1.1 病毒株与实验动物本研究所用H9N8 亚型AIV A/duck/Guangxi/S10590/2020(H9N8)株,简称DK/GX/S10590/2020 株,由国家禽流感参考实验室分离、鉴定和保存;10 日龄SPF 鸡胚购自国家禽类实验动物资源库;6 周龄BALB/c 雌性小鼠购自北京维通利华实验动物有限公司。
1.2 主要试剂病毒RNA 提取试剂盒购自天根生化科技(北京)有限公司;M-MLV 反转录酶及RNA 酶抑制剂购自美国Promega 公司;EasyTaqDNA 聚合酶购自北京全式金生物技术有限公司;PCR 纯化试剂盒购自美国OMEGA 公司;测序试剂盒Big Dye Terminator 3.1 购自美国ABI 公司。
1.3 病毒纯化与鸡胚半数感染量(EID50)的测定将病毒原液用PBS 10 倍倍比稀释,取106~109病毒稀释液接种10 日龄SPF 鸡胚,每个稀释度接种4 枚鸡胚,37 ℃孵化48 h 后收取最高稀释度且血凝价最高的鸡胚尿囊液,按照上述方法连续传3 代后得到纯化病毒,将F3 代病毒液用PBS 10 倍倍比稀释至1010,105~1010每个稀释度接种5 枚10 日龄SPF 鸡胚,37℃孵化48 h 后检测各稀释度血凝阳性的鸡胚数目,并根据Reed-Muench 方法计算病毒的EID50。
1.4 病毒全基因组测序与分子进化分析利用RNA提取试剂盒提取F3 代病毒的RNA,并利用12 bp 流感病毒反转录通用引物(5'-AGCAAAAGCAGG-3')将其反转录为cDNA。采用文献中PB2、PB1、PA、HA、NP、NA、M、NS 基因节段特异性引物分别扩增病毒的这8 个基因节段序列[10],PCR 产物纯化后利用Big-Dye Terminator 3.1 测序试剂盒测序,利用DNAStar 的Seqman 软件包拼接序列,并对其进行BLAST 比对分析。对测序结果利用MEGA 7.0 中的邻近法(bootstrap值设为1000)构建表面基因(HA、NA 基因)的进化树;利用贝叶斯[BEAST(1.10.4版)]端点定年法估算该病毒8 个基因节段的最近共同祖先时间(tMRCA),BEAST 中所使用的最佳核苷酸替代模型由IQ-TREE中的ModelFinder 计算得出,所使用的分子钟和树先验(Tree prior)分别为不相关的对数正态分布宽松分子钟和恒定大小的溯祖。并分析分离病毒的特殊氨基酸位点。
1.5 病毒对BALB/c 小鼠的感染性试验将F3 代病毒稀释至106EID50/50 μL,以50 μL/只剂量感染8 只6周龄的BALB/c 雌性小鼠;同时设立5 只接种PBS 的空白对照组小鼠。感染3 d 后,感染组随机剖杀3 只小鼠,无菌采集其脑、鼻甲、脾脏、肾脏和肺脏,利用鸡胚进行病毒滴定,检测病毒在小鼠体内的复制情况;同时,感染后每天观察并记录感染组和对照组小鼠的临床表现、体质量变化和死亡情况,直至感染后第14 d。
2 结 果
2.1 病毒纯化及EID50测定结果有限稀释法连续纯化3代后获得F3代纯化病毒,利用Reed-Muench 方法检测计算得到病毒的EID50为7.28 EID50/0.1 mL。
2.2 病毒的全基因组测序及同源性分析结果利用BigDye Terminator 3.1 测序试剂盒对DK/GX/S10590/2020 株的全基因组测序,将测得的全基因组序列输入NCBI 数据库进行BLAST 比对分析,筛选与DK/GX/S10590/2020 各基因节段核苷酸同源性最高的参考株序列。结果显示:与分离株8 个基因节段核苷酸同源性最高的病毒有H1N1、H3N8、H4N8、H6N1、H7N3、H7N7 和H9N2亚型AIV,基因来源呈现明显的多样性(表1),提示该病毒可能是由多种LPAIV 在病毒传播过程中经复杂重配而成。
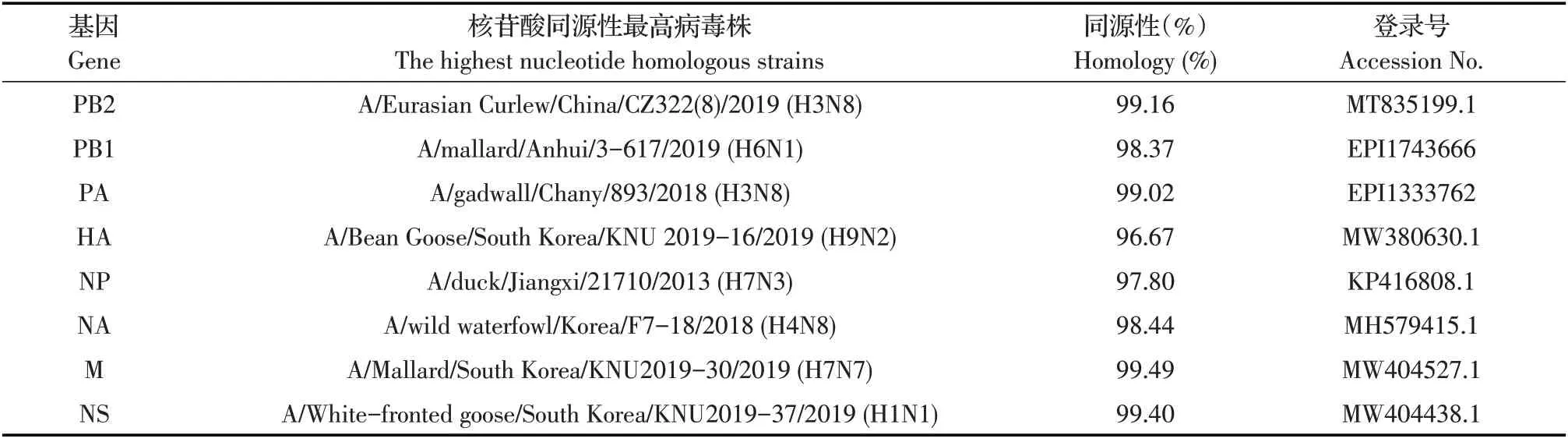
表1 与DK/GX/S10590/2020株各基因节段同源性最高的病毒株Table 1 Virus with the highest nucleotide identity to each gene segment of DK/GX/S10590/2020
2.3 病毒表面基因的遗传演化分析结果对该病毒分离株表面基因进行遗传演化分析,HA 基因进化树显示:DK/GX/S10590/2020 株与我国目前流行的BJ94-like 和G1-like H9N2 AIV 处于不同的两个分支,而与西伯利亚、韩国、日本和孟加拉国来源的H9N2 AIV HA 基因亲缘关系较近,提示其HA 基因可能来源于东亚-澳大利西亚候鸟迁徙线上的外来源H9N2 AIV;NA 基因进化树显示,该病毒株的NA 基因与NCBI 数据库中H9N8 AIV 的NA 基因位于不同的进化分支,而与多株阿拉斯加野鸟源H3N8 AIV 亲缘关系较近,提示东亚与北美洲的N8-NA 基因库发生了明显的基因漂移(图1)。以上结果表明,DK/GX/S10590/2020 株可能是由东亚-澳大利西亚候鸟迁徙线上的野鸟引入广西鸭群。

图1 分离株DK/GX/S10590/2020 HA和NA基因的遗传进化树Fig.1 Phylogenetic trees of the HA and NA gene segments of DK/GX/S10590/2020
2.4 病毒起源的进化分析结果为了进一步推算DK/GX/S10590/2020 株的产生时间,利用贝叶斯端点定年法对分离病毒的8 个基因节段进行分析。结果显示,其NP 基因的最近共同祖先来源于中国的鸭源AIV,其余7 个基因的最近共同祖先均来源于东亚地区的野鸟源AIV(表2),表明该H9N8 AIV可能由东亚地区的野鸟源AIV和我国的鸭源AIV重配而成。该病毒的PB2、PB1、PA、NA、M 和NS 基因的tMRCA为2017 年5 月 至2018 年12 月,HA 和NP 基 因 的tMRCA分别估算为2015年12月和2015年6月(表2),表明DK/GX/S10590/2020 株的重配过程主要发生在2017年~2018 年,可能在2018 年末完成重配。

表2 分离株DK/GX/S10590/2020(H9N8)的tMRCATable 2 Time of most recent common ancestors for DK/GX/S10590/2020(H9N8)
2.5 病毒基因组特殊氨基酸位点分析结果对DK/GX/S10590/2020 株的特殊氨基酸位点分析,结果显示,HA 蛋白裂解位点氨基酸位点为PAASNR/GL,仅含有一个碱性氨基酸,为明显的LPAIV 分子特征。本研究中该病毒的HA 蛋白受体结合区具有155T 和226Q的分子特征;NA 蛋白茎部区无缺失。该H9N8 分离株的PB2 氨基酸序列中具有增强对小鼠毒力的504V 分子特征[11]。此外,还在该分离株中发现许多可增强H9亚型AIV对鸡或哺乳动物组织嗜性的突变,如PB1蛋白的L13P、PB1-F2 蛋白的L82S、PA 蛋白的K26E 和V160D、NP 蛋白的V105M 和K398Q、以及M1 蛋白的H110Y和A166V[2]。但未发现该病毒具有药物抗性的突变(如NA 蛋白的E119A/G 和H274Y/R;M2 蛋白的L26P、V27A、A30T 和S31N),表明该分离株对流感病毒的抑制药物仍敏感。以上结果表明,DK/GX/S10590/2020 为一株LPAIV,但其存在感染鸡和哺乳动物的潜在风险。
2.6 病毒对BALB/c 小鼠的感染性试验结果以BALB/c 小鼠作为动物模型,评估DK/GX/S10590/2020株对哺乳动物的感染性。结果显示:仅在2 只小鼠的鼻甲内检测到了病毒低水平复制,其病毒滴度均为1.25 log10EID50/mL,在小鼠的肺脏、脑、脾脏和肾脏内均未检测到该病毒复制(图2)。感染后14 d 观察期内,感染组小鼠无明显的临床症状,但相比于对照组,感染组小鼠整个实验期体质量总体呈波动缓慢增长趋势,但在感染后第4 d~6 d 小鼠的体质量显著下降(P<0.05)(图3),此结果可能是因为感染后第4 d 该病毒在小鼠体内的复制达到最高水平。以上结果表明,DK/GX/S10590/2020 株对小鼠呈低致病力。

图2 感染后第3 d小鼠脏器病毒滴定结果Fig.2 Viral titers in the organs of mice at 3 days post inoculation
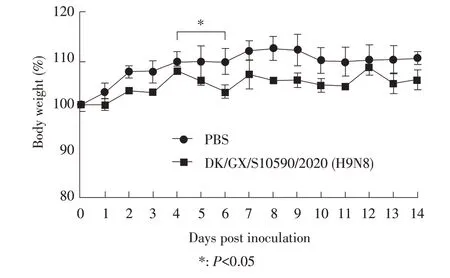
图3 小鼠感染病毒14 d内的体质量变化Fig.3 Changes in of body weight of in mice infected with the virus over in 14 days
3 讨 论
由于自然地理屏障的存在,AIV 的基因组可以分为两个独立进化的谱系,即欧亚谱系和北美谱系。有时这种障碍可以被一些长距离迁徙候鸟(如针尾鸭)所消除[12]。以往研究表明,欧亚谱系和北美谱系的AIV 之间存在部分基因节段的交换,或通过候鸟的迁徙进行AIV 的扩散传播[13-14]。阿拉斯加作为东亚-澳大利西亚迁徙路线和北美迁徙路线的主要重叠区,是AIV 洲际间基因重组最为频繁的地区之一[15-16]。本研究中DK/GX/S10590/2020 株的NA 基因与阿拉斯加野鸟源H3N8 AIV 的NA 基因位于同一进化分支,表明二者可能有共同的基因来源。
流感病毒HA 蛋白受体结合区的特殊氨基酸位点是决定其宿主范围的关键因素。研究表明155T(以H3的HA 编号)能够显著增强H9 AIV 对α-2,6 人源唾液酸受体的结合能力[17],本研究中的DK/GX/S10590/2020 株HA 蛋白具有该分子特征。NA 茎部缺失能够增强流感病毒对小鼠的毒力,并与流感病毒从野生水鸟跨物种感染家养陆禽后的适应性有关[18],但本研究中该H9N8 分离株NA 蛋白茎部无缺失。PB2 蛋白中的一些突变(Q591K、E627K、D701N)可显著增强AIV 对哺乳动物的适应性[19-21],本研究中的DK/GX/S10590/2020 株未发现这些氨基酸突变,但其PB2 蛋白存在504V 分子特征,该位点可以增强AIV 对小鼠的毒力[11]。虽然在该病毒株内部蛋白上发现多个增强对鸡或哺乳动物组织嗜性的突变,如PB1 蛋白的L13P、PB1-F2蛋白的L82S、PA蛋白的K26E 和V160D、NP蛋白的V105M 和K398Q、以及M1 蛋白的H110Y 和A166V[2],但本研究小鼠感染性试验结果显示,该病毒仅在小鼠的鼻甲内低水平复制,而不能在小鼠的肺脏复制,说明该病毒株还未完全适应哺乳动物,但不排除该病毒株可以感染家鸡的可能性。
候鸟迁徙在AIV 的传播过程中起着关键作用。目前已知全球共有9 条主要的候鸟迁徙路线,其中有3 条途经我国,即中亚迁徙路线、西亚-东亚迁徙路线和东亚-澳大利西亚迁徙路线。本研究的病毒分离自广西的一个鸭养殖场,恰好位于东亚-澳大利西亚迁徙路线上;贝叶斯进化分析结果表明该病毒可能是由东亚地区的野鸟源AIV 和我国鸭源AIV 重配而成后,由东亚-澳大利亚迁徙线上的野鸟引入广西鸭群。在我国家鸭大多数处于开放式饲养状态,缺乏有效的生物安全防护措施,与野鸟接触密切,为AIV 在家鸭与野鸟之间交流重组提供了便利条件。因此,为了有效预防AIV,尤其是防止高致病性AIV由野鸟传入家禽,应当进一步加强水禽饲养环节的生物安全防护措施,同时提高水禽禽流感疫苗的免疫覆盖率和免疫合格率,阻断AIV 的侵入,从而减少因为水禽的养殖和流通带来的AIV 传播风险。
猜你喜欢
铁道建筑技术(2022年10期)2022-10-28
中国典型病例大全(2022年12期)2022-05-13
中国生物制品学杂志(2022年3期)2022-05-13
中国典型病例大全(2022年9期)2022-04-19
皮肤病与性病(2021年3期)2021-07-30
求学·理科版(2020年7期)2020-07-14
参花·青春文学(2020年3期)2020-03-18
森林与人类(2016年2期)2016-03-07
筑路机械与施工机械化(2014年3期)2014-03-01
