
茶多糖结构特征研究进展
2022-09-30 07:49:54陆安霞叶玉龙
食品科学 2022年17期
陈 丽,陆安霞,刘 飞,叶玉龙,*
(1.宜宾学院农林与食品工程学部,四川 宜宾 644000;2.四川省农业科学院茶叶研究所,四川 成都 610066;3.精制川茶四川省重点实验室,四川 成都 611130)
茶多糖不同于茶叶中的纤维素、半纤维素、淀粉等均一性多糖,是一类由单糖、蛋白质和矿质元素等单元组成的具有生理活性的复合植物多糖。因其具有抗氧化、降血糖、抗癌等多种生物活性,受到国内外茶叶研究者的广泛研究,已有大量报道对茶多糖的生物活性进行了详尽的总结,但其结构特征仍缺乏系统的综述。
茶多糖的结构是其呈现生物活性的基础,组成十分复杂,具有品种多样性、生育期多样性和茶类多样性等。作为一类生物大分子,其层次的分类沿用了蛋白质的分类法,也可分为1~4级结构,本文主要综述了茶多糖在茶中的含量、分子质量、组成单元、糖苷键的类型、糖链分支的位置等一级结构特征以及茶多糖分子三维结构、微观形貌等高级结构,以寻求其中规律。
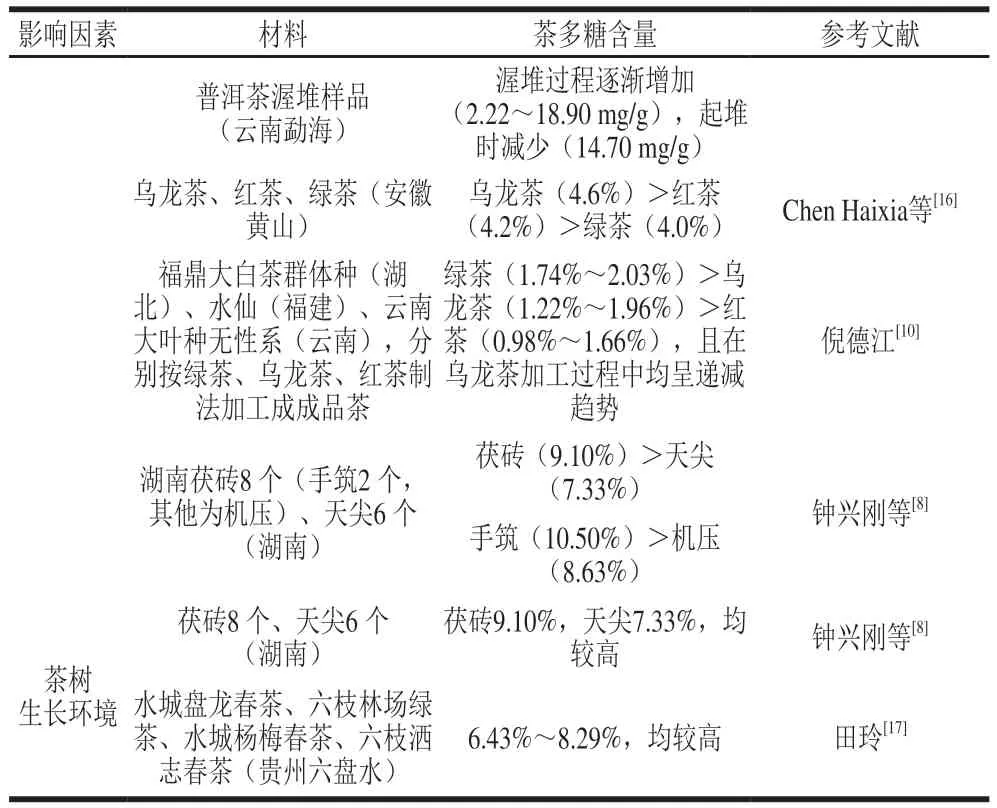
1 茶中茶多糖的含量
茶多糖在茶中的质量分数在0.14%~10.73%之间,除了样品本身的影响以外,与提取、纯化和定量方法有很大的关系。茶中杂质较多且茶多糖分子质量差异较大,茶多糖定量尚欠缺精准方法。一些研究用蒽酮-硫酸法定量,此方法测定的结果为可溶性总糖的含量,粗多糖中单糖、二糖和寡糖的存在会使检测结果偏高;对于结合有蛋白质、无机元素等成分的茶多糖,该方法测定结果偏低。需要注意的是,与蒽酮-硫酸反应后,茶多糖产物的最大吸收波长与常用的葡萄糖(glucose,Glu)标准品不同,故使用Glu标准品会导致检测结果偏低,宜用半乳糖(galactose,Gal)、阿拉伯糖(arabinose,Ara)或一定比例的混合多糖作为标准品。也有研究者通过茶多糖提取率来反映其含量,粗多糖中的色素、蛋白等杂质的存在同样会导致结果偏高,而提纯后的茶多糖因分离纯化过程中的损失导致结果偏低。
在不同的茶树品种中,倪德江分析了9 个茶树品种茶多糖的含量,揭示了茶多糖的品种多样性,赵雪丰发现大叶种乌龙茶多糖含量高于中、小叶种。在不同生育期茶叶中,多数研究结果表明随着茶叶成熟度的增加,参与缩合的单糖种类可能增多,分子质量变大,茶多糖含量随之增加,但倪德江的研究结果与之相反,可见茶多糖含量与茶叶成熟度并非完全呈正相关,造成这种差异的原因目前尚未有相关报道说明。在茶树的不同器官中,茶花的茶多糖含量高于茶果皮和茶叶。在同一片茶叶中,叶片正面通常比背面具有更高的茶多糖含量。在不同的茶类中,黑茶的茶多糖含量普遍较高,普洱茶多糖含量在渥堆过程中逐渐增加,起堆时降低,茯砖茶多糖含量高于天尖茶多糖含量,手工筑包茯砖茶多糖含量高于机械压包茯砖茶多糖含量。一是因为黑茶的原料较老,黑茶原料本身含有较多的茶多糖;二是微生物在渥堆过程中起到了重要作用,普洱渥堆产生的曲霉和茯砖发花产生的冠突散囊菌等菌属能够促进束缚态茶多糖的分解,产生游离态茶多糖,同时也能够促进茶叶中不溶性的碳水化合物、蛋白质等分解,与其他物质络合,增加各种可溶性茶多糖的含量,而手工筑包的紧实程度、通气性更有利于提高微生物活性,黑茶起堆时酵母菌呼吸的糖消耗量大于糖生成量,导致茶多糖含量降低。对于其他的茶类,学者们普遍认为发酵茶多糖含量高于不发酵茶,但也有研究结果与之相反,即绿茶高于乌龙茶和红茶,且在乌龙茶加工过程中多糖含量均呈递减趋势,这可能是品种和发酵工艺的差异导致的。如上所述,发酵过程中束缚态的茶多糖同样向游离态转化,使茶多糖含量上升,而茶多糖水解程度同样很大,同时也会发生非酶促褐变反应,生成相应的醛类和吡咯类、吡嗪类等含氮化合物,使茶多糖含量下降,成品茶中茶多糖的含量取决于各因素综合影响程度。适宜的茶树生长环境也有利于茶多糖的积累。钟兴刚检测到湖南茯砖和天尖茶多糖含量均较高,这是因为研究原料产自湘中和湘西茶区,该地区气候温暖,降水量适中,土壤以红、黄壤为主,日照时间短,昼夜温差大,环境适宜;同样,田玲检测到贵州六盘水的4 种茶样茶多糖含量也较高,这也与六盘水冬无严寒、夏无酷暑、雨量充沛、雨热同季、昼夜温差大的环境特点有关。
综上所述,茶多糖含量受茶树品种、成熟度、茶树器官、茶叶加工方法、茶树生长环境等多种因素的综合影响,各因素对茶多糖含量的影响见表1。

表1 不同因素影响下的茶多糖含量Table 1 The polysaccharide content of tea varies depending on various factors
续表1
2 茶中茶多糖的平均分子质量
分子质量是茶多糖最重要的物理特性之一,茶多糖属于大分子化合物,其纯品指的是一定分子质量范围的相对均一多糖组分,所以茶多糖经过分离纯化后,需进行纯度的鉴定。常用于测定多糖分子质量的方法包括凝胶渗透色谱法(gel permeation chromatography,GPC)和凝胶过滤色谱法(gel-filtration chromatography,GFC)。在不同茶叶样品中发现的茶多糖重均分子质量在2.56~3 900.00 kDa之间(表2)。
影响茶多糖分子质量大小的因素有提取纯化方法、茶树器官部位和加工工艺等。茶多糖分子质量会受洗脱液的影响,用蒸馏水洗脱的茶多糖是一种非均相多糖,用NaCl溶液洗脱,随着NaCl洗脱浓度的升高,多糖的分子质量降低且更均匀。在茶树不同器官中,茶花多糖的分子质量最大,为2.56~1 460.00 kDa,茶叶多糖和茶籽多糖的分子质量分别为3.67~758.00 kDa和3.66~961.00 kDa。不同茶类中,绿茶(9.2~251.5 kDa)比乌龙茶(5.3~100.9 kDa)的分子质量大,红茶最低(3.8~32.7 kDa),这可能是发酵过程中碳水化合物被酶水解导致的,发酵程度越深,茶多糖分子质量越小。白茶属于不发酵或轻发酵茶,分子质量为29 kDa。而普洱茶中茶多糖的分子质量比绿茶多糖GTPS(251.5 kDa)大10 倍以上,这与渥堆过程中茶多糖含量增加的原理相同。
3 茶中茶多糖的组成单元
3.1 中性单糖与糖醛酸
中性单糖和糖醛酸是茶多糖的主要组成成分,以下3 种方法可以同时检测二者的组分:1)与1-苯基-3-甲基-5-吡唑酮柱前衍生后用反相高效液相色谱(reversed-phase high performance liquid chromatography,RP-HPLC)分析;2)用三氟乙酸水解茶多糖的糖苷键后,对分解得到的单糖进行乙酰化衍生再用气相色谱(gas chromatography,GC)或气相色谱-质谱(gas chromatography-mass spectrometer,GC-MS)分析;3)通过离子交换色谱(ion exchange chromatography,IEC)直接定性定量,IEC法比GC-MS法更方便、准确。咔唑-硫酸法和间羟基联苯法常被用来测定糖醛酸的总量,有研究表明中性己糖和戊糖会对此方法的检测结果造成干扰,间羟基联苯法可以排除中性糖对检测结果的影响。
目前在茶多糖中主要发现了8 种中性单糖,分别是Glu、鼠李糖(rhamnose,Rha)、Ara、甘露糖(mannose,Man)、核糖(ribose,Rib)、木糖(xylose,Xyl)、Gal和岩藻糖(fucose,Fuc),在样品中含量普遍较高的是Gal、Glu、Rha和Ara,其中93.75%的茶多糖含有Gal,90.62%的茶多糖含有Ara。糖醛酸只有半乳糖醛酸(galacturonic acid,GalA)和葡萄糖醛酸(glucuronic acid,GluA)两种,含糖醛酸的茶多糖仅有15.62%,这些茶多糖中,糖醛酸占总糖的12.14%~63.11%,部分中性单糖和糖醛酸的结构如图1所示。
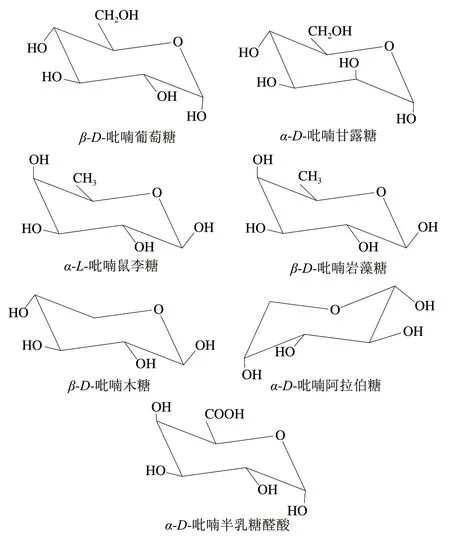
图1 茶多糖中典型的中性单糖和糖醛酸结构Fig. 1 Typical monosaccharide and uronic acid structures in tea polysaccharides
茶多糖根据中性单糖和糖醛酸的比例不同可分为中性茶多糖和酸性茶多糖,Wang Yuanfeng等测得中性茶多糖含有82.7%的总糖,其中糖醛酸只占12.9%,而酸性茶多糖含有85.5%的总糖,糖醛酸占39.8%,但二者的界定范围尚不明确。它们的单糖组成种类有一定的差异,中性茶多糖中的单糖主要是Gal(67.6%),酸性茶多糖主要含有Rha、Ara、Gal、GalA以及少量Rib。
1.3 判定标准 将原始得分转化为百分制,评分高于60分者视为健康素养合格;参加孕妇学校分为定期参加、偶尔参加及未参加3个等级,课程为期3个月,2次/周,共24次课程;喂养方式:母乳喂养,婴儿仅食用母乳,未使用任何其他事物;混合喂养,同时给予婴儿母乳与加工食品;人工喂养,完全未食用母乳,一直予以奶粉等加工食品喂养。
中性单糖与糖醛酸的组成和含量同样与实验方法、鲜叶的成熟度、茶树不同器官、茶叶加工方法、茶树生长环境等多种因素有关。
沸水提取的茶多糖含有除Fuc和Rib以外的其他6 种中性茶多糖,酶法辅助提取的茶多糖缺少Xyl和Man;纯化过程中,用蒸馏水洗脱得到的茶多糖富含GluA,是一种典型的酸性多糖,用NaCl洗脱的组分均是中性茶多糖;Scoparo等用碱法提取绿茶和红茶中的茶多糖,得到茶多糖粗提取物,再经过透析、冻融和离心分离出可溶性和不可溶性茶多糖两部分,可溶性多糖的中性单糖主要是Ara、Gal,而不溶性多糖主要含有Xyl。茶树不同器官中,茶叶、茶籽和茶花中的茶多糖均含有Rha、Ara、Gal、Glu、Xyl、GalA和GluA,茶叶多糖还含有Man和Rib,茶籽多糖也含有Man。茶花多糖的中性糖(62.84%)质量分数高于茶叶多糖(59.62%)和茶籽多糖(47.58%)质量分数;在Wei Xinlin等的研究中,茶花多糖的糖醛酸质量分数最低,为14.32%,茶叶和茶籽多糖质量分数分别为24.32%、22.78%;Wang Yuanfeng等在茶花多糖中发现了GalA,未发现GluA。以上研究结果表明,茶花多糖是一类中性茶多糖。针对茶树生育期对单糖含量的影响,学者们的研究得出了不同的结果,招钰等认为随着茶叶成熟度的增加,中性糖含量增多,而糖醛酸含量变化不大,而倪德江却发现随着鲜叶的老化,多糖中性糖和糖醛酸含量均呈先增加后减少的趋势,峰值出现在第2叶。发酵过程中存在的强烈酶促氧化会使中性糖、糖醛酸含量都降低,多糖中的糖链和肽链均被显著降解。招钰等用同一种提取检测方法,分析得出绿茶及乌龙茶的糖醛酸含量明显高于黑茶和红茶,尤其是红茶糖醛酸的含量最低,约占茶多糖总量的1.95%;乌龙茶多糖中性糖质量分数最高(63.09%),其次为黑茶(61.25%)、绿茶(48.01%)和红茶(31.31%),这也与乌龙茶及黑茶原料较粗老有关。陈浩用酵母菌对云南普洱大叶种晒青绿茶进行人工发酵,发酵前期普洱茶多糖中糖醛酸质量分数为20.08%,发酵40 d后糖醛酸质量分数增长至27.41%;对贮藏1、3 年和5 年的普洱茶分析发现,3 种普洱茶多糖都是酸性多糖,且糖醛酸含量随着茶叶贮藏年份的增加而增加,这是因为黑茶加工过程中微生物的作用往往大于热分解和自身酶促氧化,使糖醛酸含量上升。
3.2 蛋白质与氨基酸
许多研究发现,茶多糖经重复脱蛋白后仍含有少量蛋白质,表明茶多糖是蛋白质结合的多糖,进一步的研究表明蛋白质通过N—或O—共价键与糖链结合在一起。前人对茶多糖结合蛋白和组成蛋白质的氨基酸展开了一系列研究,与游离蛋白质和氨基酸的检测方法一样,蛋白质的鉴定主要采用紫外光谱法,含量分析采用考马斯亮蓝法(Bradford)、凯氏定氮法以及福林-酚法;氨基酸主要通过酸水解后用HPLC或氨基酸分析仪进行分析。
茶多糖结合的蛋白质在茶叶中的比例为1.87%~38.00%。Lu Xinshan等用Bradford法测定得出黄山毛峰绿茶多糖蛋白质量分数为7.1%;Wang Yuefei等同样用Bradford法测定了从乌龙茶中分离出的TTPS、FTPS和DTPS 3 种茶多糖中蛋白质量分数分别为5.57%、7.68%和9.30%;Scoparo等在可溶性绿茶和红茶多糖中发现了12%和19%的蛋白质;普洱茶多糖的蛋白质量分数为20.0%。由此并未发现茶多糖中蛋白质的存在规律。此外,并不是所有的茶多糖都含有蛋白质,Lee等从绿茶中得到酸性多糖组分中未检测到蛋白质。
在茶多糖中共检测到了18 种氨基酸。Chen Haixia等用HPLC法在绿茶多糖中检测到了15 种氨基酸,以甘氨酸(glycine,Gly)、谷氨酸和天冬氨酸(aspartic acid,Asp)为主,其质量比为1.16∶0.94∶0.50。Wang Dongfeng等用氨基酸分析仪对安徽茶样的茶多糖氨基酸进行研究,发现了17 种氨基酸,含量较高的也是谷氨酸(24.23%)、Asp(18.27%)和Gly(17.89%)。可溶性绿茶多糖中组成蛋白质的氨基酸以羟脯氨酸(hydroxyproline,Hyp)、丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和半胱氨酸(cysteine,Cys)为主;在可溶性红茶多糖的蛋白质中,含量最丰富的氨基酸是Hyp、Ser、Thr和谷氨酸,值得注意的是,Cys在绿茶多糖中的相对含量为9.6%,而红茶多糖中没有检测到Cys。福建白茶多糖的蛋白质包含17 种氨基酸,以谷氨酸、脯氨酸(proline,Pro)、苯丙氨酸(phenylalanine,Phe)、亮氨酸(leucine,Leu)、Ser和Asp为主。西湖地区茶树鲜叶多糖中含有15 种必需氨基酸,总含量为68.06 mg/g,蛋白质含量为58.49 mg/g,说明茶多糖中除了组成蛋白质的氨基酸还含有独立连接糖链的氨基酸。
茶多糖分子质量及单糖、蛋白质和糖醛酸的组成见表2。
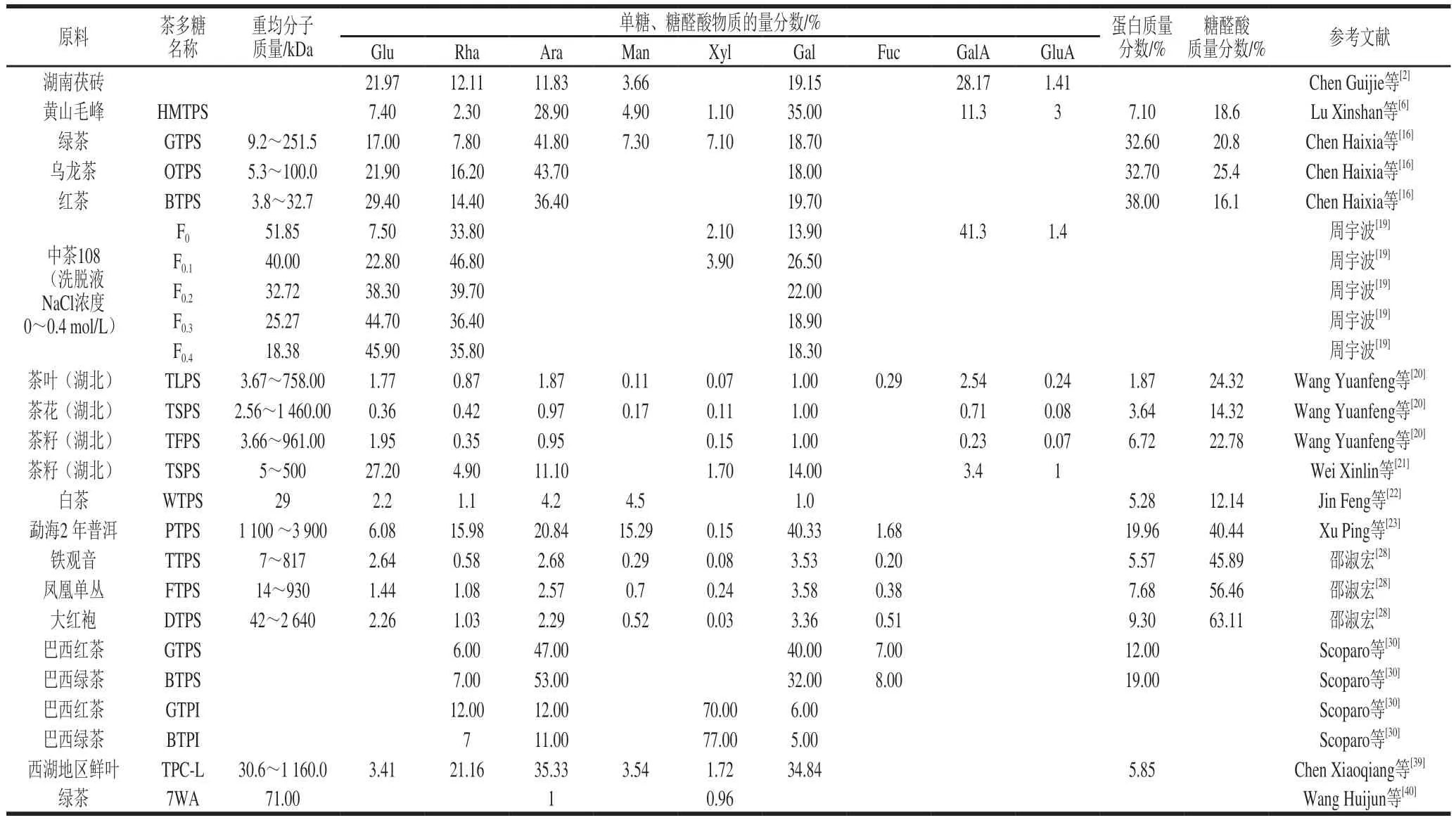
表2 茶多糖分子质量及单糖、蛋白质和糖醛酸的组成Table 2 Molecular mass, monosaccharide composition, and protein and uronic acid contents of tea polysaccharides
3.3 无机元素
在茶多糖中也发现了铁、镁、锌、硒和稀土元素(rare earth,REE)等无机元素,无机元素在茶多糖中的含量主要由原子荧光光谱(atomic fluorescence spectroscopy,AFS)或电感耦合等离子体发射光谱仪(inductively coupled plasma emission spectrometer,ICP)测定。富硒茶的茶多糖硒含量同样较高,Wang Yuanfeng等报道了富硒绿茶中分离到的Se-TPS、Se-TPS1、Se-TPS2、Se-TPS3 4 种茶多糖分别含硒1.987、0.970、0.440 μg/g和0.340 μg/g。稀土微肥在我国广泛应用,以提高包括茶叶在内的作物的产量和质量,因此茶多糖中也存在稀土元素,Wang Dongfeng等在茶多糖检测到1 mg/g REE,包括La、Ce、Nd、Pr和Y共5 种,它们是自然界中最常见的REE,占总量的90%左右。
4 茶中茶多糖的糖链结构
茶多糖糖链结构研究包括异头物的构型、糖苷键类型、单糖序列、糖链有无分支、分支的位置和长短等。能有效解析糖链结构的方法有化学方法、物理方法和生物方法,其中最为常用的方法包括红外光谱法、高碘酸氧化法、部分酸水解法、Smith降解法、甲基化分析法和核磁共振(nuclear magnetic resonance,NMR)法等。
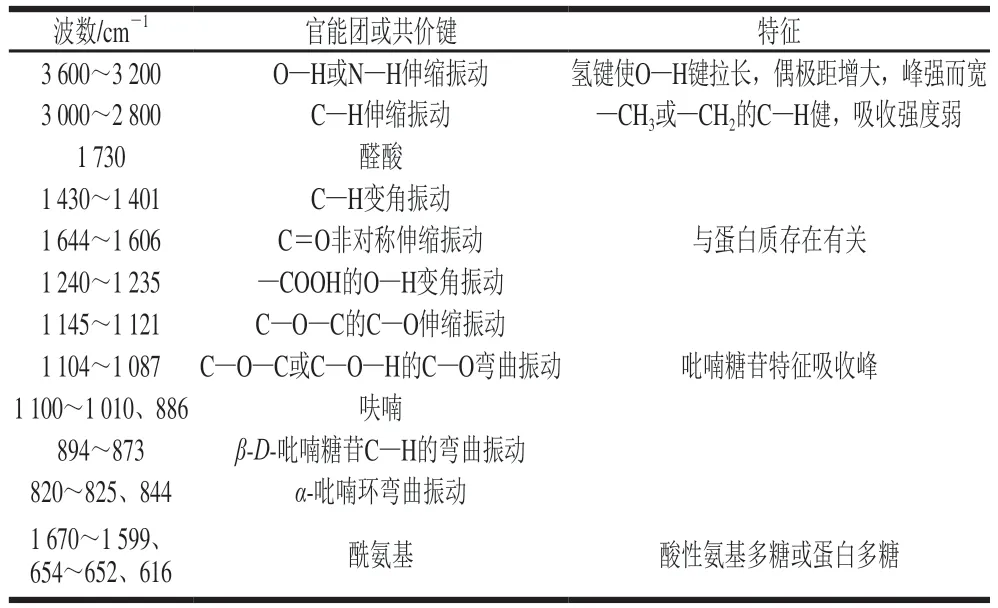
表3 茶多糖的红外光谱分析[10,19,22,28,43]Table 3 Infrared spectral analysis of tea polysaccharides[10,19,22,28,43]
高碘酸氧化可选择性断裂糖分子中的连二羟基或连三羟基之间的C—C键,生成相应的多糖醛、甲醛或甲酸。每开裂1 个C—C键消耗1 分子高碘酸,通过测定高碘酸的消耗量及甲酸的释放量,可以推断糖苷键的位置。若1 mol糖残基消耗2 mol高碘酸的同时释放1 mol甲酸,证明该单糖残基中含有连三羟基,推测是以1→6或1→键合的吡喃己糖;若1 mol糖残基只消耗1 mol高碘酸,证明该单糖残基中含有连二羟基,推测是以1→2、1→4、1→2,6、1→4,6键合的吡喃己糖,或1→2、1→4键合的吡喃戊糖,或1→5键合的吡喃戊糖;若糖残基不消耗高碘酸,证明该单糖残基中无连羟基,推测是以1→3键合的吡喃己糖、吡喃戊糖、呋喃戊糖,或1→2键合的呋喃戊糖。Smith降解是将高碘酸氧化产物还原成稳定的多羟基化合物后酸水解,根据水解产物的种类,如甘油或赤鲜醇,推断糖苷键的位置;再根据产物的量并结合高碘酸反应的结果进一步推测单糖残基类型。→2)---葡萄糖-(1→的Smith降解反应式如图2所示。
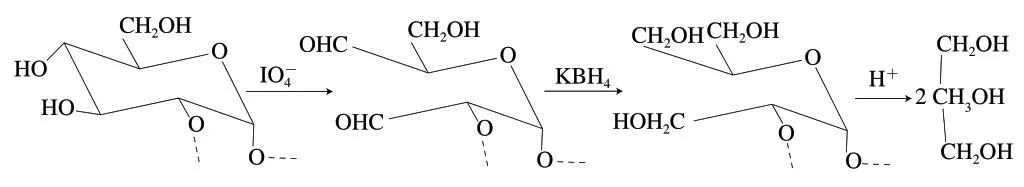
图2 →2)-α-D-葡萄糖-(1→的Smith降解反应Fig. 2 →2)-α-D-Glcp-(1→ Smith degradation reaction
部分酸水解可以将茶多糖大分子裂解成小片段,其作用包含两个方面:1)判断单糖在糖链上的位置,对多糖进行分级水解时支链上的单糖较骨架上的单糖更易水解;2)帮助判断糖苷键和单糖残基的类型,1→6糖苷键对酸水解相对稳定、呋喃型糖残基较易水解、己糖醛酸糖苷键可抵抗酸水解。
甲基化反应可用来判断糖苷键的连接位点,发生反应时糖基的游离—OH全部被—CH取代,经三氟乙酸水解后产物的—OH所在位置即为单糖残基的连接点,吡啶-乙酸酐衍生得到甲基化糖醇乙酸酯后进行GC-MS分析,用于确定糖苷键的位置和单糖的物质的量比。
H和C二维NMR被用来解析复杂的茶多糖结构信息,包括通过C、H原子化学位移来判断单糖的类型;通过耦合常数推测茶多糖分子中各原子之间的连接关系;通过信号强度估计单糖残基的重复数量;通过对比糖残基C原子与单糖C原子化学位移确定糖基相连位置等。
茶多糖的糖链结构见表4。可以看出主链主要由Gal、Glc、Rha组成,其中Gal最普遍,Ara多在侧链上,Man和Fuc常在糖链末端,侧链通过O-3、O-4、O-6与主链连接,Gal既有-型又有-型,Rha往往以-型存在。酸性茶多糖的糖醛酸多位于主链;同一种原料制作的红茶和绿茶多糖糖链结构相似;可溶性茶多糖和不溶性茶多糖结构差异很大,水溶性茶多糖以→3)-Gal-(1→和→6)-Gal-(1→为主,侧链中存在-Araf分支,这与Wang Huijun等检测发现的水溶性茶多糖7WA结构相似;而不溶性茶多糖则以少见的→4)--Xylp-(1→为主。
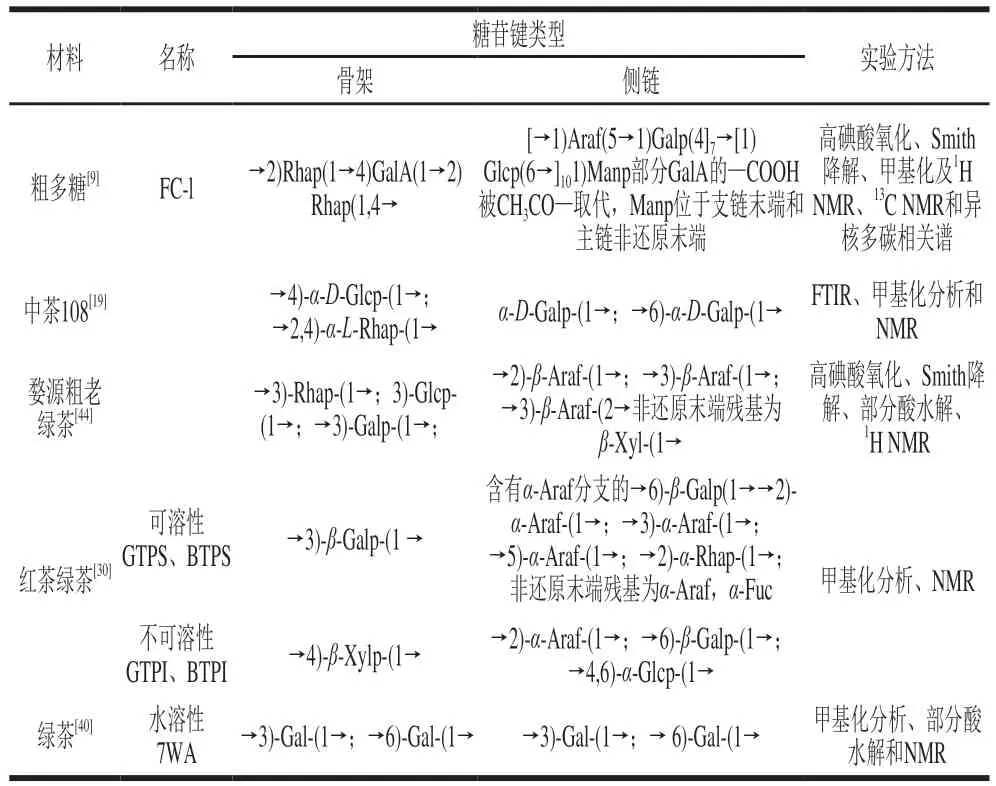
表4 茶多糖的糖链结构Table 4 Sugar chain structures of tea polysaccharides
5 茶中茶多糖的高级结构
茶多糖的微观形貌特征采用原子力显微镜(atomic force microscopy,AFM)、扫描电子显微镜(scanning electron microscopy,SEM)和圆二色光谱(circular dichroism,CD)等方法观察。
AFM主要是观察茶多糖聚集体的形状和大小,它利用细微的探针感知针尖端和样品表面原子之间的相互作用力,通过原子力的变化反映多糖聚集体表面的起伏变化。茶多糖单链的直径一般为0.1~1.0 nm,而AFM下观察到的茶多糖直径要大得多,这是因为多糖分子并非为单个糖链分子,而是由多个糖链缔合而成,羟基和糖醛酸羧基极易本身或与另外一糖链形成分子内或分子间氢键,导致多个糖链缠绕,形成线团状或者箱式结构,这也是同一种多糖可测定到不同的分子质量的原因。茶多糖聚集体的形状和大小受提取方法、加工工艺和酸碱度的影响。其分子的平均直径、长度随着NaCl洗脱液浓度的增加逐渐减小;从挤压茶中提取的多糖表面呈块状,坚固且不平整,有较大空腔,而未挤压样品的茶多糖表面呈光滑的片状,说明在挤压加工中,热能和机械能的结合可以破坏多糖的微观结构,使之形成特定的形状;不同茶类多糖随着茶叶发酵度的增加,茶多糖的长度缩短,绿茶多糖和乌龙茶多糖的形状呈分枝线状,而红茶多糖呈球状,三者长度分别为620~1 526、430~1 296 nm和430~1 130 nm;乌龙茶叶多糖在溶液中呈直径0.2~0.5 μm、长0.3~0.6 μm的短棒形不均一的聚集体,在酸性环境下呈分散的短棒状,在碱性环境下形成更大的聚集体,因为茶多糖的聚集状态跟自身的化学组成有关,羧基的存在使茶多糖含负离子,而载体云母是一种硅酸铝,同时带负电荷,长度较短、COO—含量高的茶多糖受云母排斥力影响较大。也有研究表明温度和离子强度等外界因素对茶多糖构象的稳定性起着重要作用。
SEM利用电子束与样品间的相互作用激发出各种信号,通过这些信号的接受、转化、放大和显示成像,可在荧光屏上直接微观成像,用于研究茶多糖分子的外形长度、伸展状态、柔顺性大小以及空间构象。SEM下观察到的茶多糖分子呈颗粒状,结构松散,随着糖醛酸和蛋白质含量的增加,分子间相互作用位点增多,作用力增大,颗粒间紧密结合,因各茶多糖分枝情况不同,形成的分子间氢键的位置和作用力大小也不同,就呈现出多层次片状、无规则团状、串珠状、螺旋状等多种立体结构。利用SEM在低倍镜下还能观察到乌龙茶多糖的折叠卷曲结构,而在高倍镜下则呈现平整的图像,这一观察结果与绿茶多糖的SEM图是完全不同的,这是在乌龙茶加工过程的酶促降解作用下,糖链和肽链变短,分子质量减小,分子之间的氢键作用增强导致的。
CD利用不对称分子对左、右偏振光吸收的不同来分析茶多糖分子在水溶液中的绝对构型、构象等三维结构信息。因为茶多糖尤其是中性茶多糖缺乏在一般紫外区的光谱活性,难以直接由CD分析得到结构信息,通常先将茶多糖进行分子修饰或与刚果红(CHNOSNa)反应后进行CD测定推测茶多糖的构象,具有三股螺旋结构的茶多糖可以与刚果红形成络合物使最大吸收波长移向长波。周鹏等将婺源粗老绿茶多糖与刚果红反应后,刚果红的最大吸收波长由230 nm移向了490 nm(样品与刚果红混合物),说明样品在溶液中呈现螺旋构象;张艳用刚果红-NaOH鉴定发现购买的粗茶多糖样品中不存在三螺旋结构;江和源等分析了3 个茶多糖纯化组分的Cotton效应,并拟合计算了这3 个组分所含蛋白质的二级结构相对含量,发现这些糖蛋白质存在-折叠、-螺旋和无规卷曲。
茶多糖的结晶性较差,不适用于常规X射线单晶衍射,利用纤维衍射结合立体化学信息与计算机模拟,可以准确确定茶多糖构型。也有研究者借鉴并运用了部分高分子领域分析方法,如激光扫描共聚焦显微镜(laser scanning confocal microscopy,LSCM)、激光光散射-尺寸排阻色谱联用法,对不同分子质量茶多糖的空间结构信息进行了初步探讨。倪德江利用LSCM首次发现茶多糖聚集体带有强烈的荧光,产生的原因是否同构象相关,尚需进一步证实。
6 结 语
茶多糖结构特征的研究尚不透彻,一方面是因为茶多糖结构复杂,需要结合多种仪器的分析结果进行准确解析,目前这些贵重仪器普及范围仍然不广,部分实验人员也缺乏相关仪器的专业操作训练,这给茶多糖结构的研究带来了很大的阻力;另一方面,原料和实验方法不同都会导致研究结果不同,这就需要研究人员控制单一变量,探明提取纯化检测等方法及茶树品种、茶树器官、加工工艺等多种因素对茶多糖结构的影响。此外,对于产生这些影响的更深层原因,即茶多糖合成及降解机理鲜见详细报道,仅有学者对糖链片段的化学合成进行了一些尝试,对于茶多糖的生物合成途径、热裂解和酶解的机理需进一步深入研究。
茶多糖的生物活性与其结构特征密切相关,现有的茶多糖构效关系的相关报道包括4 个方面:一是茶多糖的分子质量是影响其生物活性的一个重要因素。一些学者认为分子质量大的茶多糖由于水溶性较差、表面黏度大,难以通过组织屏障进入细胞内部,限制了其抗氧化活性。也有一些学者有相反的看法,认为高分子质量茶多糖的己醛酸含量较高,金属螯合能力较低,清除自由基和抑制脂质过氧化活性更高;二是茶多糖的生物活性明显与单糖组成和糖醛酸含量有关。Scoparo等发现红茶和绿茶多糖即使具有相似的糖链,但二者的GalA残基含量差异导致其抗败血症的能力不同;Wang Yuefei等发现乌龙茶多糖的抗氧化性和-葡萄糖苷酶抑制活性明显与糖醛酸含量成正比;三是结合无机元素和蛋白质的茶多糖具有更高的生物活性。Chen Haixia等发现蛋白含量较高的茶多糖组分具有更强的抗氧化活性;Wang Yuanfeng等发现富含无机元素Se的紫阳富硒茶多糖可以显著减少剧烈运动引起的氧化应激,具有更高的抗氧化活性;四是修饰茶多糖结构可以提高其生物活性,如梁进等对茶多糖分别进行硫酸化、乙酰化和羧甲基化修饰后,多糖主链和支链的空间结构发生改变,导致活性基团暴露,茶多糖的负电荷以及溶解度增大,从而使其抗凝血活性进一步增强。进一步深入分析阐明茶多糖的分子结构,有助于茶多糖构效关系和生物活性的分子机制研究,从而为茶多糖作为药物临床应用或作为天然产品在功能性食品中的应用提供新的见解,通过修饰茶多糖结构来拓展其生物功能也正在成为一个新的研究方向。
猜你喜欢
生物化学与生物物理进展(2022年10期)2022-10-25 04:20:06
医药前沿(2020年18期)2020-12-03 04:42:50
天然产物研究与开发(2019年1期)2019-03-01 05:41:20
生物技术进展(2018年6期)2018-11-28 09:45:08
高等学校化学学报(2016年9期)2016-11-03 02:10:08
天然产物研究与开发(2016年1期)2016-06-05 10:29:25
现代食品(2016年24期)2016-04-28 08:11:48
特产研究(2016年3期)2016-04-12 07:16:26
中南民族大学学报(自然科学版)(2015年2期)2015-12-16 12:11:10
食品工业科技(2014年15期)2014-03-11 18:17:19
