
荞麦脱壳性状的研究进展
2022-09-29 11:16:06马贵芳辛海波孙朝霞
中国农学通报 2022年24期
马贵芳,辛海波,修 莉,孙朝霞,张 华
(1北京市园林绿化科学研究院,绿化植物育种北京市重点实验室,北京 100102;2山西农业大学,山西晋中 030801;3沈阳市浑南区城乡建设综合执法中心,沈阳 110000)
0 引言
荞麦(Fagopyrum esculentumMoench.)隶属蓼科,自花授粉、生育期短、适应性广、种质资源丰富,是一种原产于中国且广泛分布的一年生或多年生双子叶草本植物,主要种植在亚洲和东欧地区[1]。荞麦基因型多样,用于农业生产上主要有甜荞和苦荞[2]。荞麦作为一种药食同源作物,含有丰富的淀粉、维生素、脂肪及微量元素钾、钙、镁等,同时含有多种人体必须氨基酸和抗氧化化合物,这些物质在疾病预防或治疗中发挥着重要的作用[3-5]。近年来,荞麦作为一种生产营养保健品和功能食品的健康作物受到越来越多的关注,以荞麦为原料的产品越来越多,如荞麦壳枕头、荞麦茶、荞麦酒、荞麦饼等,这使得荞麦在全球的产量日益增加[6-8]。荞麦籽粒难脱壳的性状,极大地限制了荞麦产业的发展。常见的荞麦脱壳方法有撞击式脱壳和碾压脱壳,这些方法在很大程度上使荞麦的外观受到严重破坏,且脱壳效率低、产率低,均无法达到规模化生产[9-10]。此外,荞麦的脱壳处理方式还有采用水浸泡、蒸制、烘干后再进行脱壳处理,这样的脱壳方式势必破坏苦荞麦仁的营养成分,且工序繁杂,出仁(胚)率低,难以得到理想的产品原料[11]。改良荞麦种子裂壳性状,提高荞麦加工效率、减少营养流失,是目前育种的主要目标之一。近年来,研究表明荞麦壳性状受遗传和发育的调控,荞麦壳细胞的排列方式、细胞壁降解相关酶的含量、胚乳与壳之间蜡质角质层的合成及细胞壁合成相关转录因子的调控均发挥着重要作用,这使果壳主要组成物质发生差异代谢,呈现出壳厚度的差异性状。因此,本研究进展从以上几个方面概述了荞麦壳性状相关基因及代谢物的变化,以期为薄壳荞麦育种提供全面的理论依据。
1 荞麦分类及果壳的成分和细胞结构
根据脱壳的难易程度,荞麦可分为稻谷型和非稻谷型两种。非稻谷型外壳较厚,且紧紧附着在胚乳上,去除难度大;而稻谷型外壳较薄且松散,不附着在胚乳上,容易去除(如图1所示)。相比稻谷型荞麦,非稻谷型荞麦在卵巢中脉附近的表皮下出现了一层薄薄的细胞(图1c),在衰败花的子房中,细胞层更厚,且木质素含量更高(图1d,e),这是外壳和胚乳之间存在附着力的原因之一。同时研究发现该细胞层出现在花芽长轴长度在0.8~1.8 mm时,这表明此类细胞的分裂发生在卵巢发育早期(图2)[12-14]。研究者对稻谷型荞麦与非稻谷型荞麦的杂交后代分析发现,稻谷型易脱壳性状可能是由一个隐性纯合基因控制,但该基因尚未被报道[15]。研究表明果壳成分包含有木质素、纤维素、半纤维素和果胶等,其含量决定了果壳的厚度和机械性能是否良好。如长角果在成熟过程中木质素和纤维素的比例对果荚发育和开裂至关重要,木质素的积累导致内壁组织变厚,从而促进果荚的开裂[16]。大豆在成熟时,背部和腹部的豆荚会形成一个脱落层,并且有1,4-β-葡聚糖酶和内聚半乳糖醛酸酶积累在在裂解区,在成熟过程中其含量逐渐增多,这是导致豆荚破裂的原因之一[17-19]。此外,有研究表明脱落现象发生时,除了需要酶作用之外,也需要小蛋白(15k~25 kD)和蜡质的生物合成和分泌,其在分离细胞的表面形成了可扩展的细胞外基质,有助于果荚开裂及籽粒脱壳[20]。

图1 非稻谷型(NT)品种(Hokkai T8和Manten-Kirari)和稻谷型(RT)品种(Mekei T28)的卵巢和果实的组织学特征
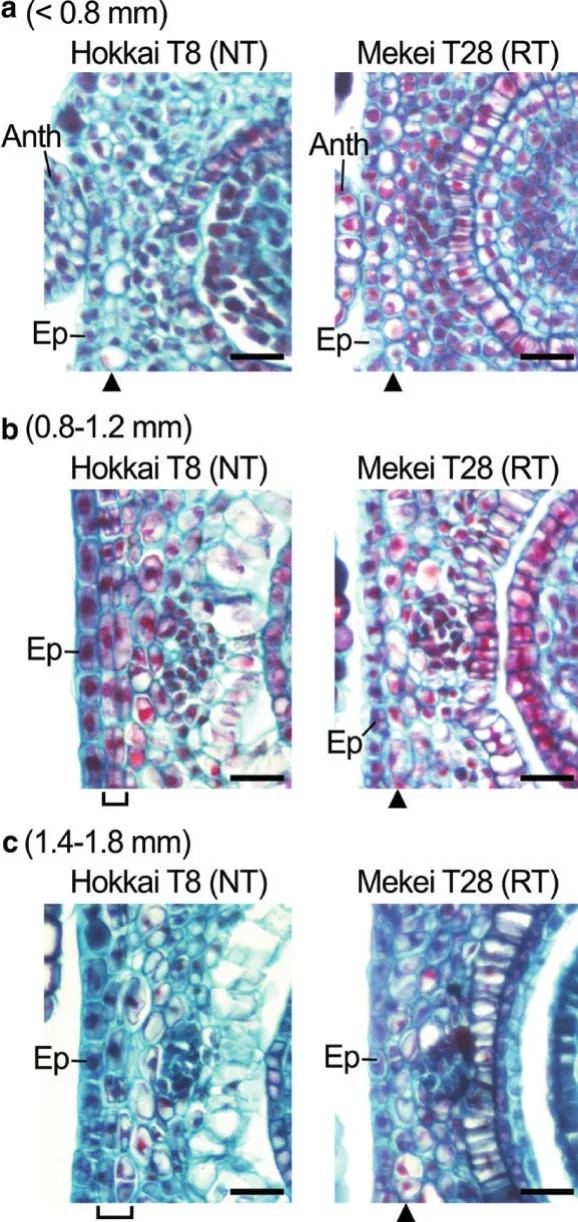
图2 荞麦非稻谷(NT)品种和稻谷(RT)品种花芽细胞长轴长度及子房中脉表皮下周壁细胞组织学特征
果壳作为种子的重要组成部分,由子房壁发育而来,对胚乳和胚起到保护作用[21]。如表1所示,不同品种壳的成分大致相同[22-23]。其中木质素在植物细胞壁的结构完整性方面发挥重要作用,纤维素在细胞壁拉伸强度发挥着至关重要的作用[24-26]。木质素积累量越多,其细胞壁越容易开裂,而纤维素含量越多,其细胞壁的韧性越强,越不容易开裂[27-28]。吴朝昕[29]研究表明荞麦厚壳和薄壳不同品种间纤维素、半纤维素及木质素含量之间存有明显的差距。SONG等[30]研究发现在荞麦种子发育成熟过程中,木质素和纤维素的含量均增加,但难脱壳品种在后期发育过程中,木质素含量下降,纤维素含量增加。SUANUM等[31]在红豆中也同样发现了此规律。此外,荞麦壳的厚度及是否易脱壳与其细胞类型及排列密切相关。李淑久等[32]发现苦荞壳除内表皮、外表皮外,在内外表皮间有多层厚壁组织细胞;而甜荞在内外表皮间由3层厚壁组织细胞组成;野生荞麦品种在内外表皮间由一层厚壁组织细胞和一层石细胞构成。吴朝昕[29]研究发现难脱壳苦荞品种‘晋荞2号’和易脱壳苦荞品种‘小米荞’的果壳均由外表皮、横向厚壁组织、纵向厚壁组织和内表皮四部分组成,其中易脱壳品种果壳厚壁组织较薄,外表皮和横向厚壁组织的纤维素含量较低,木质素含量较高。在荞麦壳生长发育形成期,LI等[33]发现纤维素及半纤维素合成酶基因在难脱壳品种中高表达,易脱壳品种中低表达,这也可能是造成果壳薄厚差异的关键因素。
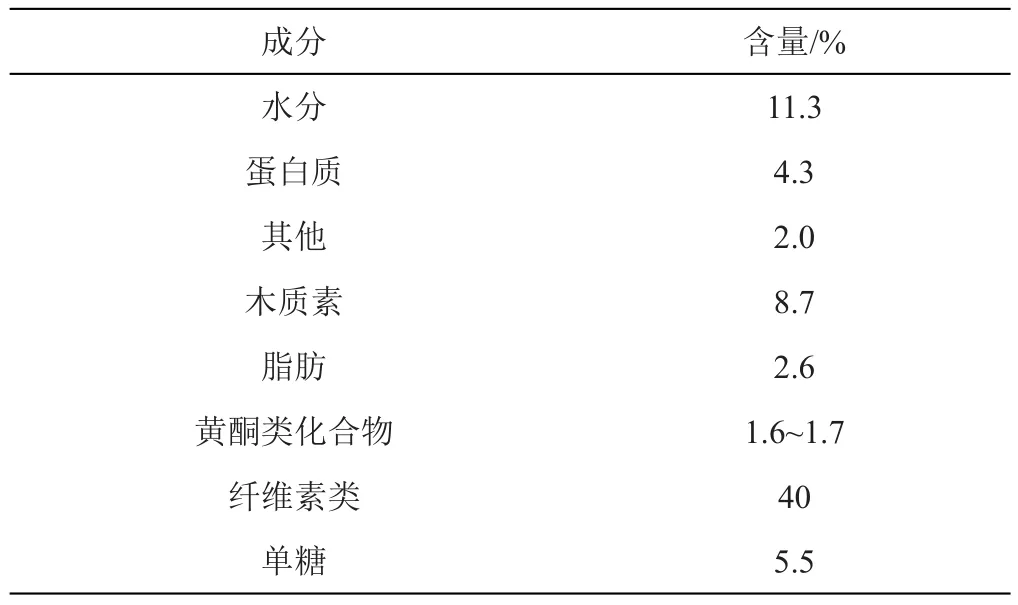
表1 荞麦壳主要成分
2 酶在种壳开裂过程中的功能
果壳壳裂性状被认为与叶子及果实脱落具有共同的特征,均发生在精确的位置,且伴随着细胞壁的崩解和细胞的分离[34]。脱落过程分为4个时期:脱落区分化、响应脱落信号、脱落激活(该时期细胞壁和胞外基质发生变化诱导器官分离)和保护层形成及脱落后横向分化(图3)[35-36]。在此过程中,细胞壁松弛相关的、编码木葡聚糖转糖基酶/水解酶(XTHs)、膨胀素(EXPs)等基因发生特异表达,这将有利于细胞壁降解酶进一步发挥作用[37-38]。细胞壁主要成分纤维素被纤维素酶(CELs)水解为单糖,果胶被多聚半乳糖醛酸酶(PGs)水解为β-半乳糖醛酸,这使得细胞内聚力丧失,从而产生裂缝[39-40]。研究表明,在果荚开裂时检测到与脱落相关的酶如PGs、CELs活性均有所增加[41-43],这在大豆及拟南芥中均被证明[17-38]。如图4所示,在成熟绿色荚果的背侧缝合线处,薄壁细胞构成了大豆荚开裂区(DZ),在腹侧缝合线下面由薄壁细胞组成裂缝区,且DZ细胞呈空泡化增加;在成熟黄色荚果,背侧阀瓣边缘已经分开,并且仅通过纤维帽细胞彼此连接,裂口区的细胞全部塌陷,腹侧瓣膜分离,DZ细胞坍缩,有效地分离了瓣膜边缘。目前尚未有人测定荞麦籽粒形成过程中薄壳及厚壳荞麦品种果胶酶及纤维素酶含量差异。
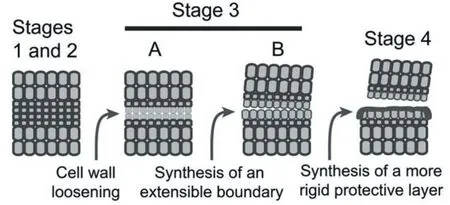
图3 脱落区形成和器官分离的模型图
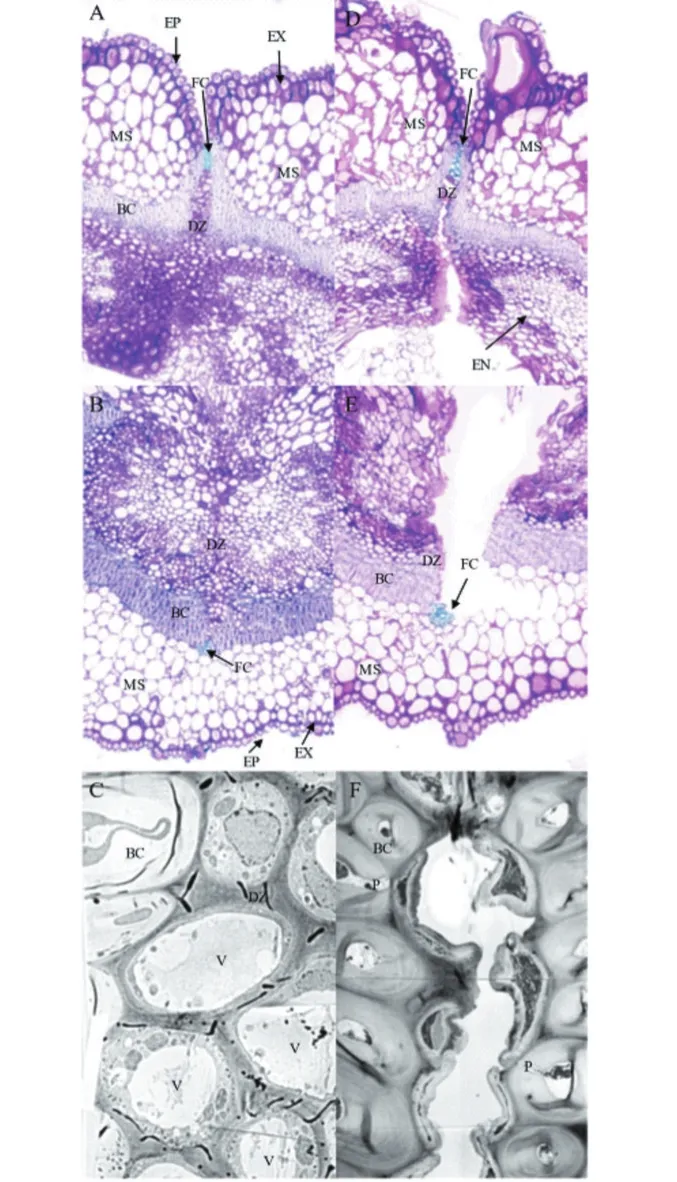
图4 大豆荚成熟期两个阶段背侧和腹侧荚果开口区横切面的光学电子显微图
3 蜡质角质层在种子外壳脱落过程中的功能
在外壳脱落过程中,控制蜡质角质层类物质合成和分泌的基因表达上调是普遍特征,这表明果皮表皮上产生的脂质与外壳和颖果间的粘连是相关的[44]。大豆、番茄和拟南芥的转录组测序数据显示,在脱落发生早期,蜡质角质层合成的相关基因表达上调早于细胞壁降解酶编码基因[45]。近些年来研究发现,蜡质角质层成分变化比细胞壁分解酶的改变对器官分离的调控更加显著,如番茄蜡质缺陷突变体表现出花器官融合,推测这是由于植物表皮细胞的脂质可以提供一层防水的保护层,防止发育过程中组织异常粘连,而失去这层保护层后导致花器官发生融合[46-48]。研究表明,脂质合成后被分泌的部位对于脱壳性状至关重要,如调控大麦脂质生物合成代谢的基因为Nud基因,属于乙烯响应因子(ERF)家族,该转录因子蛋白可激活外种皮里特定脂质的产生,跨过果皮层分泌到外表皮,不能起到分离果实和壳的作用,造成外壳和颖果粘连,因此有稃大麦颖果难以脱壳,而裸大麦易脱壳(见图5)[35,49]。水稻颖果中Os06ERF基因(Nud的同源基因)在花后7天达到最高水平,早于大麦Nud(花后14天),但其在花后不到21天就停止了表达[50]。这种表达的时间偏差可能导致Os06ERF不能激活脂质合成途径,从而不能控制水稻壳性状。将大麦Nud基因在自身启动子驱动下转化水稻,发现其仍然提早停止表达(花后21天),类似水稻内源Os06ERF的表达模式,无法作用于水稻脂质合成途径(图6)。由此可见,Nud基因在水稻和大麦颖果中的转录调控机制可能不同,也可能是水稻脱壳性状主要受细胞壁代谢等途径而非脂质合成途径调控。拟南芥WIN1/SHN1基因(Nud的同源基因)被证明主要在脱落和裂开区等细胞分离区域表达,对脂质生物合成途径和发育过程中组织的分离起着重要的作用[51-53]。目前荞麦籽粒发育过程中蜡质角质层的变化尚未被报道。
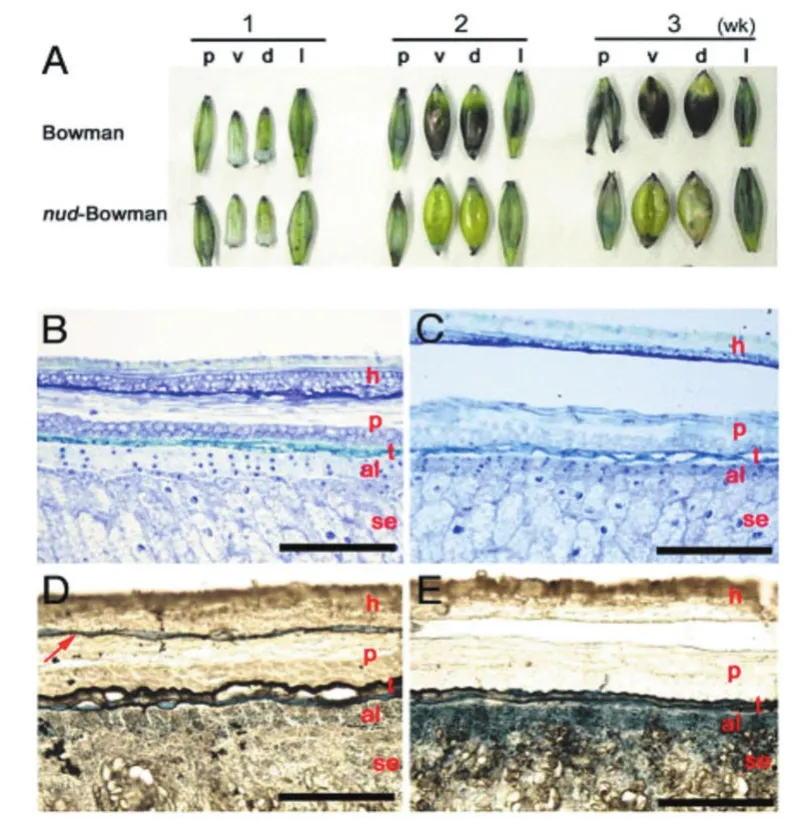
图5 大麦颖果横切面的光学电子显微图
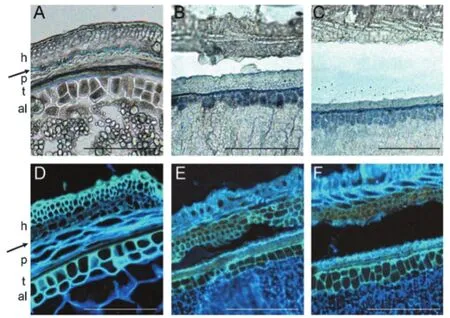
图6 颖果横切面的光学电子显微图
4 转录因子对壳发育的调控
转录因子(Transcription Factors,TFs)对基因的调控作用目前已被广泛研究[54]。转录因子通过调控细胞壁主要成分纤维素和木质素合成相关基因的表达从而调控壳裂性状。植物细胞壁主要由初生壁、次生壁和胞间层三部分组成,具有机械支持、保护和物质输导的作用,其中次生壁增厚在植物的花药、果荚开裂中发挥重要作用,调控作用的研究多集中在次生壁合成方面[55-56]。拟南芥次生细胞壁结构及其生物合成转录调控网络如图7所示[57]。
4.1 NAC转录因子
NAC蛋白是植物特异性转录因子家族,参与调节多种生物过程,包括抗逆性、木质部的形成[58]。AIDA等[59]首先报道了并命名了NAC结构域。之后NAC转录因子被克隆并其抑制顶端分生组织的分化[60]。2005年,MITSUDA等[61]研究发现拟南芥中的花药开裂可能是由两个密切相关但复杂冗余的NAC转录因子调控的,一个是NST1,它仅在花药壁中少量表达,另一个是NST2,它在花药中表达强烈但活性有限,这两个基因的表达导致拟南芥花药内膜次生壁增厚,从而导致花药不能开裂缺陷,这显示NAC转录因子可能对次生壁增厚具有一定的作用。2007年,MITSUDA等[62]又证明NST1和NST3通过调节拟南芥木质组织形成来影响其次生壁的形成。ZHONG等[63]同样证明了NST1、NST2和NST3是调节拟南芥花序茎和下胚轴的次生壁形成的关键调控因子,且三者存在功能冗余作用。荞麦薄壳壳裂性状与花药开裂性状具有相似性,LI等[33]对厚壳和薄壳品种的荞麦籽粒发育4个时期(开花后5、10、15、20天)的差异表达基因分析发现,NAC转录因子是调控荞麦壳厚的关键基因。WANG等[64]在拟南芥中过表达FtNAC16,发现其抑制S型木质素合成来降低木质素总量,从而负调控拟南芥荚果的开裂。

图7 拟南芥次生细胞壁结构及其生物合成转录调控网络
4.2 bHLH转录因子
bHLH转录因子被报道参与了果实开裂、心皮和表皮发育以及胁迫反应的转录调控[65-67]。KIM等[68]研究发现bHLH转录因子参与调控拟南芥脱落酸介导的反应,这进一步表明bHLH参与植物果实开裂的调控。在花药发育过程中,只有小孢子外的胼胝体被降解,小孢子才能被释放到花药室中并继续发育成成熟的花粉粒[69]。宋垚等[70]研究表明bHLH可调控胼胝质降解,这对于花药发育是至关重要的。RAJANI等[71]发现ALC基因编码与bHLH转录因子相关的蛋白质,促进木质化细胞层之间的不稳定非木质化细胞分化,使细胞分离,果荚开裂。AtbHLH40、AtbHLH73均属于ALC基因编码的蛋白,在拟南芥果实开裂过程中,它能促进开裂部位的非木质细胞分化,是果实开裂的必要调控因子[72]。SUN等[73]研究发现FtbHLH70是AtbHLH40的同源基因,其在‘小米荞’籽粒中表达量最高且在果实发育过程中逐渐增加,这被判断可能作为关键转录因子来调控荞麦脱壳性状。
4.3 MADS转录因子
转录因子在植物的生长发育过程中发挥着重要作用[74]。研究表明,MADS转录因子可参与调控果实开裂[75]。在拟南芥中已经发现,SHP基因(MADS基因家族)的表达对果实开裂是至关重要的。在拟南芥果实成熟过程中,SHP1和SHP2控制开裂区细胞的分化和木质化[76]。双突变体shp1/shp2植株的果实没有开裂特征[77]。在此基础上,LIU等[14]对苦荞MADS基因家族的SHP基因挖掘中发现了3个潜在靶基因FtpinG0009028100.01、FtpinG0009028000.01和FtpinG0002495300.01,通过对易脱壳荞麦和难脱壳荞麦的比较发现FtpinG0009028000.01的表达水平越高,果壳开裂概率越大,反之则相反,在未开裂的果壳中该基因的表达基本没有变化,这与拟南芥调控果实开裂的SHP基因表达模式一致,因此,研究者将FtpinG0009028000.01作为研究苦荞脱壳特性的潜在靶点。
4.4 MYB转录因子
MYB转录因子含有高度保守的重复序列,在植物体内发挥重要作用[78-79]。MYB转录因子参与植物生长发育、反应应激及物质代谢的调控等过程[80-84]。在植物细胞壁的生物合成过程中,MYB转录因子调控苯丙素和木质素合成途径中的作用目前已得到了深入的研究,同时研究发现一个基于NAC(NAM/ATAF/CUC)-MYB的转录调控网络控制次生壁的生物合成[85]。目前关于MYB转录因子的研究多集中在对花序茎及茎秆次生壁中木质素合成的调控,直接调控种壳厚度的研究尚少[86]。STEINER等[87]研究发现在myb26拟南芥突变体中,花药虽然发育正常,但在发育后期花粉成熟,花药壁收缩导致花粉粒不能释放,从而导致雄性不育。YANG等[88]进一步研究发现,MYB26通过调控NST1和NST2转录因子来影响花药次生壁的木质素含量,从而调控花药开裂。
5 展望
荞麦作为中国主要的杂粮作物之一,其籽粒及加工产品具有极高的营养价值,但其规模化生产主要受到落粒性、易倒伏、难脱壳以及结实率低等性状的限制。其中,荞麦脱壳性状与籽实生产和加工具有最直接的关系。目前,有关荞麦籽粒脱壳性状的生理和分子机制研究较少。本研究进展从果壳组成及细胞排列、酶、蜡质角质层和转录因子方面,探讨了荞麦难脱壳这一农艺性状的形成原因。早在1990年YANG等[89]研究发现荞麦的薄壳性状可能是由一对隐形基因所控制,但是由于技术的限制,没有克隆到该基因。随着近年来测序技术的飞速发展和基因组学、代谢组、转录组等多组学学科的深度交叉,研究人员定位和克隆荞麦脱壳性状调控基因的工作量及难度可能会逐步减小。对于荞麦难脱壳的性状,在以下几方面的研究可能会加速荞麦产业的发展:
(1)在前人的研究中已明确荞麦的易脱壳性状与细胞壁主要成分纤维素和木质素的含量及比例相关,但是具体调控纤维素及木质素生物合成的机制尚未明确。因此,明确荞麦籽粒发育过程中纤维素、木质素等的积累规律及相关基因的表达,将为进一步揭示荞麦脱壳的分子机制奠定基础。
(2)果胶酶和纤维素酶作为细胞壁降解的关键酶,直接影响脱壳的难易,在荞麦籽粒形成过程中薄壳及厚壳荞麦品种果胶酶及纤维素酶含量差异有待深入研究。
(3)蜡质角质层形成部位及其分泌量决定外壳与颖果间是否粘连,是影响籽粒脱壳的关键因素之一。荞麦籽粒发育过程中蜡质角质层变化的研究有待进一步深入。
目前,关于果壳的研究多集中在主要粮食作物上,如豆类、油菜类作物的果荚开裂,且均表现为易脱壳,这些研究表明,荚裂性状是由多个基因共同调控的,在分子水平上解析荞麦壳裂性状的形成对荞麦易脱壳性状的研究及荞麦产业的发展,乃至对植物果实外壳形成、开裂、脱落等研究十分重要,鉴于目前对这方面的研究基础较为薄弱,育种家在荞麦薄壳形成分子机制的研究中仍然存在着巨大的挑战。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:19:24
渔业致富指南(2020年12期)2020-12-20 00:46:32
家庭医学(下半月)(2019年8期)2019-09-25 09:02:04
今日农业(2019年14期)2019-01-04 08:57:40
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
黄河·黄土·黄种人(华夏文明)(2017年3期)2017-08-22 04:01:29
山西农经(2016年3期)2016-02-28 14:23:49
汉语世界(2014年4期)2014-04-06 05:23:40
食品工业科技(2014年15期)2014-03-11 18:17:34
