
北部湾中华鲎(Tachypleus tridentatus)幼鲎生态位及种间竞争分析
2022-09-25 09:46郭清扬左娅林吴颖肖晓波赵心忆李银康应紫薇周传江颉晓勇
海洋学报 2022年10期
郭清扬,左娅,林吴颖,肖晓波,赵心忆,李银康,应紫薇,周传江,颉晓勇*
( 1. 中国水产科学研究院南海水产研究所 广东省渔业生态环境重点实验室,广东 广州 510300;2. 河南师范大学 水产学院,河南 新乡 453007;3. 南方海洋科学与工程广东省实验室(广州),广东 广州 511458;4. 惠东县水生动物防疫站,广东惠州 516300;5. 深圳爱栖自然生态科技有限公司,广东 深圳 518112;6. 广西生物多样性研究和保护协会,广西 南宁 530012)
1 引言
鲎隶属于肢口纲(Merostomata)剑尾目(Xiphosura),是一种浅海底栖节肢动物[1],因其具有广食性,在近岸海洋生态系统中参与能量转移、物质循环及信息交流[2]。近年来中华鲎(Tachypleus tridentatus)数量锐减[3],2019年中华鲎在世界自然保护联盟濒危物种红色名录中被正式更新为濒危(EN)[4],2021年2月被列入国家二级重点保护野生动物名录。
生态位是一个种群在生态系统中,在时间和空间上所占据的位置及其与相关种群之间的功能关系与作用的总和,物种生态位宽度及生态位重叠指数体现各物种资源利用情况及种间竞争关系强弱[5-6]。种间联结体现群落内部竞争物种的替代关系[7],种间联结测度分析表达群落各物种间相互作用程度及群落动态[8]。生态位和种间联结被用于探索多种生物关系,如中国花鲈(Lateolabrax maculatus)种群特征[9]、龙头鱼(Harpodon nehereus)等游泳动物间生态关系[10]、湿地浮游植物优势种生态位研究等[11]。中华鲎所处底栖生境内生物多样,食物网关系较为复杂[2],明确该物种生存竞争将进一步深化鲎栖息地保护工作所需的科学理论认识。
北部湾海域位于广东省雷州半岛、海南省和广西壮族自治区及越南之间,是当前现存最适中华鲎栖息的区域之一[12]。作为存续4.75亿年的海洋“活化石”,中华鲎在一定程度上能够反映水域生态系统的健康程度[13-14],截至目前,尚无中华鲎幼鲎在底栖动物群落内竞争强度和种间竞争情况的相关研究。本文以北部湾滨海湿地中华鲎幼鲎生境调查资料为基础,探索中华鲎幼鲎与栖息地主要生物种类间的生态位关系,计算群落总体关联性并检验种间联结性显著程度,进行种间竞争动态模拟,以期填补中华鲎幼鲎与环境底栖动物中部分优势种种间竞争关系理论空白,为中华鲎幼鲎栖息地保护提供生态学基础理论依据。
2 材料与方法
2.1 数据组成及其来源
在前期北部湾沿岸幼鲎种群分布调查基础上[4],本研究于2018年7月选择中华鲎幼鲎典型分布区域开展幼鲎调查及潮间带大型底栖动物调查,调查地点分别为西背岭、竹林盐场、三娘湾、坡尾底、沙田及榕根山(图1)。在各调查地点依地形特点分设2~6个500 m×500 m的样方,其中西背岭地区、沙田地区和榕根山地区分别设2个样方,竹林盐场地区设6个样方,三娘湾地区设4个样方,坡尾底地区设3个样方,采用样线调查法调查样方内出现中华鲎幼鲎的密度和频度。
大型底栖动物定量采样按照《海洋监测规范》(GB 17378.1-2007)和《海洋调查规范》(GB/T 12763.1-2007)进行,各调查地点设1条采样断面,每个断面按照高、中、低潮区设站点进行随机取样。本研究取样站布设时在高潮区布设2站、中潮区布设3站、低潮区布设2站,因滩面沉积物类型较为一致,生物分布较为均匀,故每站取4个样方,样方大小为25 cm×25 cm,采样深度为30 cm,岩石岸区域每站取2个样方,若生物栖息密度很高且分布较均匀则采用10 cm×10 cm定量框取样。取样时选用40目筛网,孔径为1 mm,过滤收集底栖生物。将同一潮区的平行样混合装瓶,得到高、中、低潮位共计3份样品,用酒精进行固定保存。
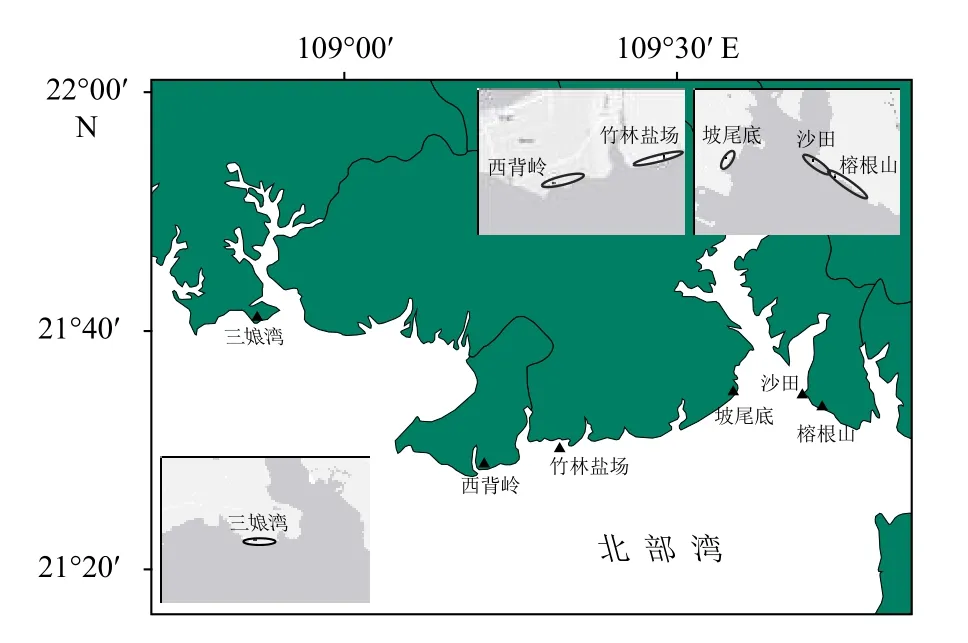
图1 采样站位Fig. 1 The location of sampling stations
各调查断面采集1份定性样品,尽可能将该地附近出现的生物种类收集齐全,以记录该地区蕴藏的底栖生物种类。所得样品用75%酒精固定5 d后进行种类鉴定,并使用精度为0.01 g的电子天平称重。依据取样面积,将个体数和湿重换算成栖息密度(单位:ind./m2)和生物量(单位:g/m2)。
2.2 数理统计方法
优势度指数计算公式为

式中,Y为优势度指数;N为样品中个体总数;ni为第i种个体数;fi为第i种在调查站点中出现频率。
Levins生态位宽度计算公式为

式中,Bi为第i种生态位宽度;j表示样方;r表示样方数量;Pij表示第i种生物占j样方总个体数的比例。
Pianka重叠指数计算公式为

式中,Qik为种i和种k的生态位重叠度;Pij和Pkj分别为种i和种k占j样方总个体数的比例。
利用方差比率(VR)法测定中华鲎幼鲎与其生境内主要优势种的总体关联性,其能够体现中华鲎幼鲎所处群落结构的稳定程度及演替趋势,并通过Ochiai指数(OI)、Dice指数(DI)、Pearson相关系数(rik1)和Spearman秩相关系数(rik2)体现两个物种间相关性的紧密程度[15]。
物种间总体关联性计算公式如下:
物种相对多度方差:
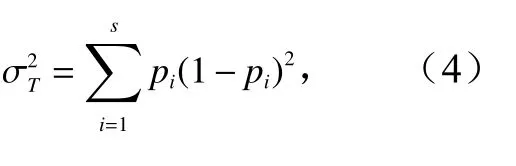
物种数方差:
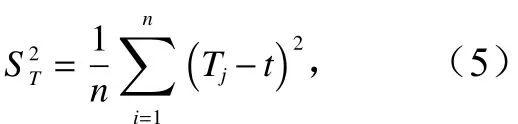
方差比率:

式中,pi为相对多度;n为样方总数;S为总物种数;t为平均物种数;Tj为j调查点物种数。当VR<1时,各物种整体呈负联结性;当VR>1时,各物种整体呈正联结性;当VR=1时,各物种间无关联性。
联结系数(AC)计算公式为
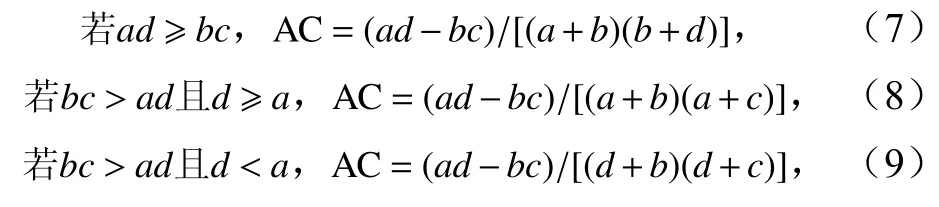
式中,a为物种A和物种B共同占据的样方数;b为仅物种A出现的样方数;c为仅物种B出现的样方数;d为均不曾有物种A和物种B出现的样方数。AC值域为[-1,1],越接近-1,即负联结程度越高,AC=0即为两物种间互相独立[16]。
Ochiai指数计算公式[7]为

Dice指数计算公式[7]为

OI与DI值域为[0,1],越接近0,即负联结程度越高。
Pearson相关系数计算公式[7]为

Spearman秩相关系数计算公式[7]为

假设中华鲎与竞争物种存在相互作用,且假设中华鲎独自在当前栖息地生存时,种群规模发展均遵循Logistic规律。得到种间竞争模型计算公式为
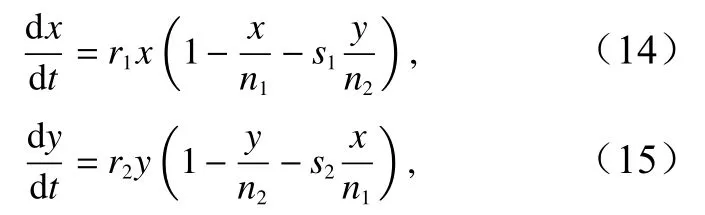
式中,x、y分别为中华鲎幼鲎与所竞争物种种群数量;r1、r2分别为固有增长率;n1、n2为最大容量;s1、s2为竞争系数。数据源自本研究团队2014-2018年间中华鲎幼鲎与短指和尚蟹(Mictyris brevidactylus)野外调查,以各地相邻年份物种数量差值计算初始固有增长率,以实地调查期间的最大样本容量值模拟最大容量,以相互竞争的两物种重叠系数及其倒数分别作为二者竞争系数。
以优势度大于0.02的物种为优势种[17],由于各取样地优势种不同,本研究选择所调查中华鲎幼鲎全部样本中出现频率大于0.10的物种作为研究对象,所有数据分析和制图均通过R3.5.1软件的spaa包实现,采样站位图由Arcgis软件制取,种间竞争模型制图由Matlab软件实现。
3 结果
3.1 中华鲎幼鲎生态位宽度及生态位重叠指数
调查期间共采集底栖生物样本12门类135科280种。其中中华鲎幼鲎生态位宽度居最宽位(4.252 2),小翼拟蟹守螺(Pirenella microptera)居最窄位(1.000 0)。依据各物种个体数计算物种优势度、出现频率和中华鲎幼鲎与其生境内主要优势种生态位宽度(表1)和生态位重叠指数(表2)。主要优势种中短指和尚蟹分布于高、中、低3个潮区,小翼拟蟹守螺主要分布于高潮区和中潮区,中阿曼吉虫(Armandia intermedia)、南海毛满月蛤(Pillucina vietnamica)、丝异须虫(Heteromastus filiformis)及丽文蛤(Meretrix lusoria)主要分布于中潮区及低潮区,熊本牡蛎(Crassostrea sikamea)、黑荞麦蛤(Xenostrobus atratus)、纹藤壶(Amphibalanus amphitrite)及韦氏毛带蟹(Dotilla wichmanni)仅分布于高潮区,突畸心蛤(Cryptonema producta)仅分布于中潮区,欧努菲虫(Onuphis eremita)和䗉螺(Umbonium vestiarium)仅分布于低潮区。
3.2 中华鲎幼鲎与各样本种间关联
本研究利用方差比率法对中华鲎幼鲎与其生境内主要优势种开展总体联结性检验,计算得方差比率VR值为0.978 022,由VR<1可知,中华鲎幼鲎与幼鲎生境内主要优势种整体表现出净负联结性[7]。联结系数(AC)显示,中华鲎幼鲎与其他各优势物种均有一定程度正向关联性(0≤AC<0.33)(图2),全部种对间有26个表现负联结性,65个种对间存在正联结性或独立状态,说明在当前演替阶段多数样本对资源利用情况相似,同时存在对食源及栖息地等资源竞争情况。OI系数值趋近于1时联结性增强,结果显示中华鲎幼鲎与南海毛满月蛤、欧努菲虫、丝异须虫、丽文蛤、短指和尚蟹具有紧密关联性(OI≥0.67),其中与南海毛满月蛤、丽文蛤及短指和尚蟹具有更紧密关联(OI≥0.83),相伴出现几率大(图3)。DI指数显示,中华鲎幼鲎与小翼拟蟹守螺相关性较弱(0.17≤DI<0.33),与其余样本相关性则较强(DI≥0.50)(图4)。Pearson相关系数测定结果显示,中华鲎幼鲎仅与短指和尚蟹(0.09≤rik1<0.39)及小翼拟蟹守螺(0.39≤rik1<0.70)呈正相关(图5),与其他样本生物均呈负相关。Spearman秩相关系数具有更高检测灵敏度,其计算结果显示,中华鲎幼鲎与短指和尚蟹关联性有所增强(0.37≤rik2<0.69)(图6)。
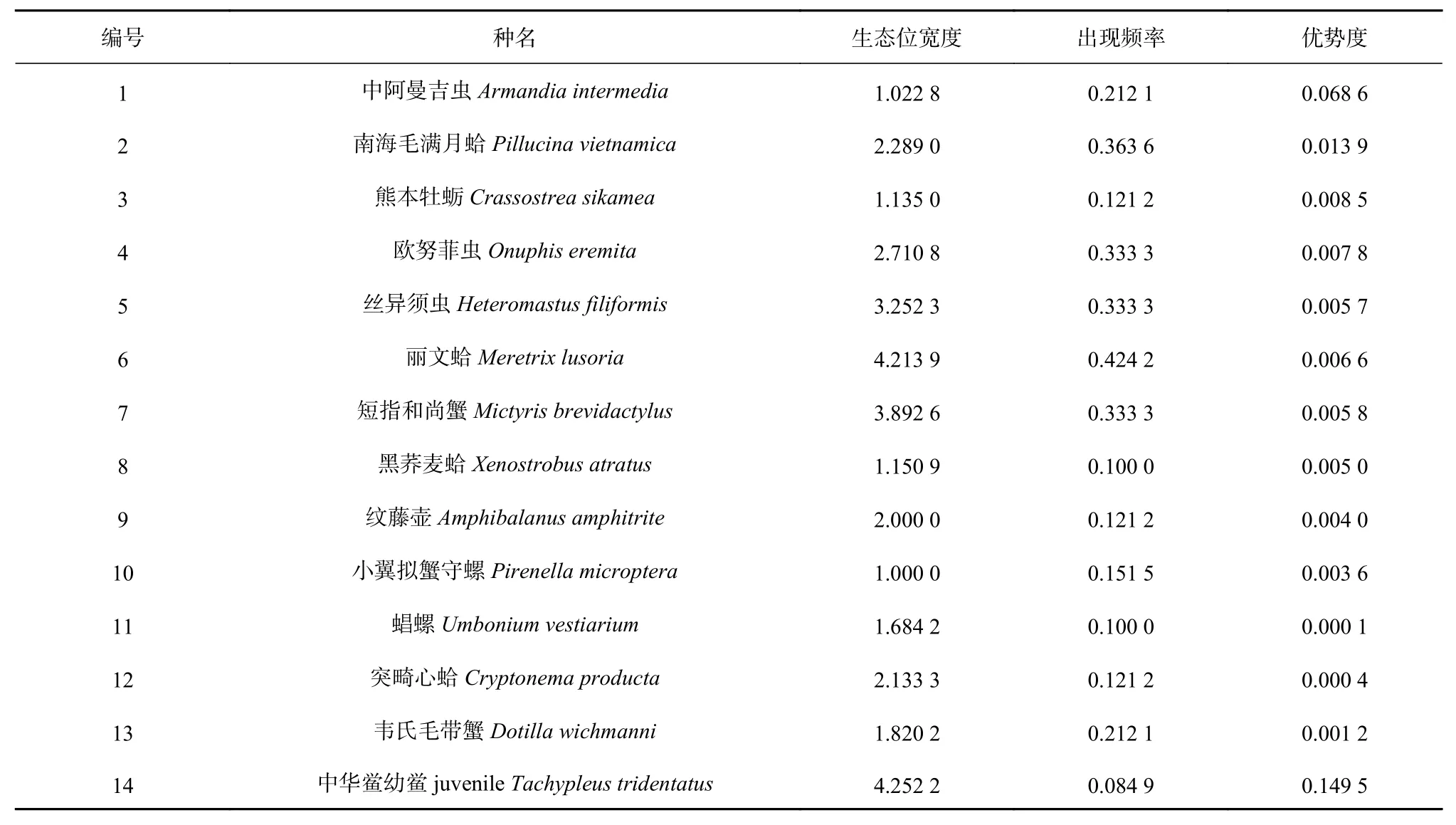
表1 中华鲎幼鲎生境主要优势种生态位宽度、出现频率及优势度Table 1 Niche width, occurrence frequency and dominance of dominant species in juvenile Tachypleus tridentatusnursery habitat

表2 中华鲎幼鲎生境主要优势种生态位重叠指数Table 2 Niche overlap index of dominant species in juvenile Tachypleus tridentatusnursery habitat

图2 中华鲎幼鲎生境主要优势种种间联结系数(AC)半矩阵图Fig. 2 Semi-matrix diagram of association coefficient (AC) of dominant species in juvenile Tachypleus tridentatusnursery habitat

图3 中华鲎幼鲎生境主要优势种Ochiai指数(OI)半矩阵图Fig. 3 Semi-matrix diagram of Ochiai index (OI) of dominant species in juvenile Tachypleus tridentatusnursery habitat

图4 中华鲎幼鲎生境主要优势种Dice指数(DI)半矩阵图Fig. 4 Semi-matrix diagram of Dice index (DI) of dominant species in juvenile Tachypleus tridentatusnursery habitat

图5 中华鲎幼鲎生境主要优势种Pearson相关系数(rik1)半矩阵图Fig. 5 Semi-matrix diagram of Pearson correlation coefficient(rik1) of dominant species in juvenile Tachypleus tridentatusnursery habitat
3.3 种间竞争动态模拟
在中华鲎幼鲎种群重要栖息地种间竞争关系分析基础上,选择短指和尚蟹作为中华鲎幼鲎关键竞争物种,利用二者固有增长率及竞争系数,以调查期间样本数最大值为种间竞争模型最大容量(表3),种间竞争模型计算结果显示,二者在西背岭和三娘湾地区呈现竞争状态,短指和尚蟹在中华鲎幼鲎种群规模减小时能够快速丰富其种群容量,竹林盐场地区则显示幼鲎与短指和尚蟹数量均呈现剧烈下滑状态(图7)。
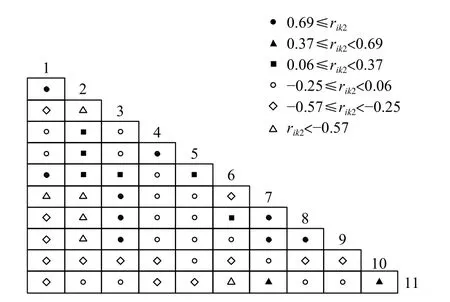
图6 中华鲎幼鲎生境主要优势种Spearman秩相关系数(rik2)半矩阵图Fig. 6 Semi-matrix diagram of Spearman rank correlation coefficient (rik2) of dominant species in juvenile Tachypleus tridentatusnursery habitat
4 讨论
4.1 中华鲎幼鲎生态位分析
中华鲎性成熟需9~12年,人类行为干预和生境破坏使得中华鲎数量锐减[4],生态位研究有助于建立其底栖生境保护和修复所需理论基础。生态位宽度可以反映物种利用不同资源的能力差异,其值越大,生物活动范围越广,对环境适应能力越强[18-19]。根据大型底栖动物生态位宽度值划分,Bi≤1.0即窄生态位种,1.0<Bi≤2.0即中生态位种,2.0<Bi即广生态位种[20]。探究底栖群落中物种生态位对保护濒危物种、修复生态结构及研究某一特定物种对群落的影响尤为重要[21]。
生境内主要优势种存在形式能够反映底栖生物对栖息地适应与否,优势种数目增加使群落结构愈发稳定[22-23]。本研究结果显示,中华鲎幼鲎为广生态位物种,其生态位宽度在所有样本中居最宽位,野外调查发现,包含砂质、泥质、礁石及红树林中一种或多种组合的滩涂均有中华鲎幼鲎出现,理论计算结果与中华鲎幼鲎自然分布相一致,均显示中华鲎幼鲎对各类生境有较强适应能力。较宽生态位物种种群数量愈少反映该区域生物群落受影响程度愈深,因此中华鲎幼鲎适合作为滨海湿地生态系统健康程度指示物种。

表3 各取样地中华鲎幼鲎与短指和尚蟹种间竞争模型参数Table 3 Parameters of interspecific competition model between juvenile Tachypleus tridentatusandMictyris brevidactylusin various sampling location
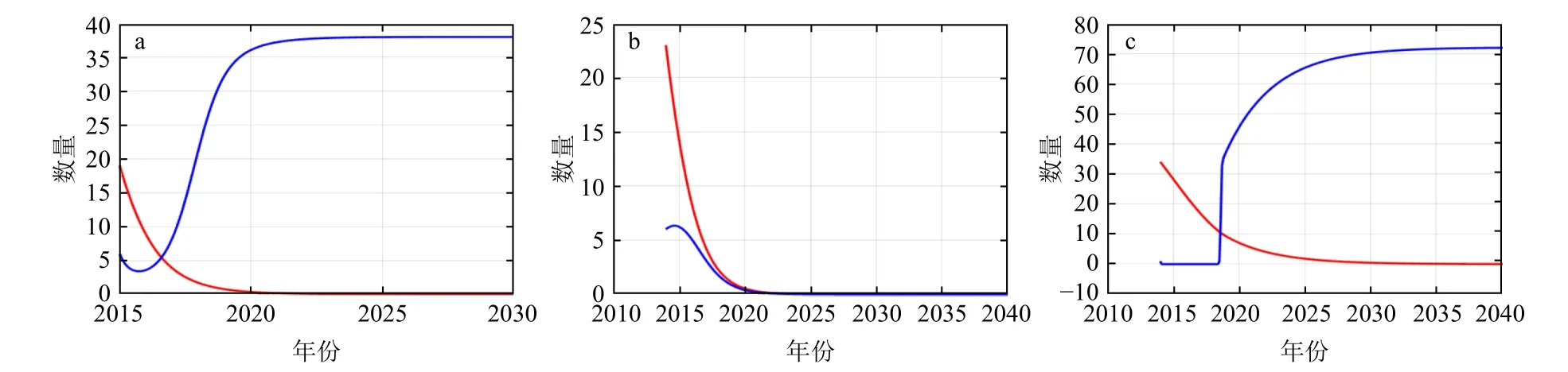
图7 不同采样地点中华鲎幼鲎与短指和尚蟹种间关系Fig. 7 Interspecific relationship between juvenile Tachypleus tridentatusandMictyris brevidactylusin various sampling location
生态位重叠指数(Qik)表达各物种利用资源的相似程度,体现物种间潜在竞争程度[24]。Qik值分3个层级,Qik≥0.6表示生态位重叠程度高,0.6>Qik≥0.3表示重叠程度适中,0.3>Qik表示重叠程度较低[25]。与生态位宽度较窄物种相比,较宽物种生态位重叠指数通常较高[26]。本研究显示,短指和尚蟹为与中华鲎幼鲎生态位重叠程度较高物种,其生态位宽度高达3.892 6。但生态位重叠程度并非完全和生态位宽度保持正相关关系[27],如南海毛满月蛤生态位宽度为2.289 0,其与中华鲎幼鲎生态位重叠指数仅为0.262 6,该结果与调查期间南海毛满月蛤集中分布于榕根山地区有关,二者生活空间重叠较低,但均在各自生境内具较强适应能力。作为中华鲎潜在饵料,南海毛满月蛤宽生态位和低重叠指数可使其自然种群规模较少受到中华鲎影响。
同区域栖息物种具有相似生态特性或对生境因子有互补性要求,其在形态学、生理学及生活史特征方面存有差异,并以此降低种间竞争[28-29]。中华鲎幼鲎与短指和尚蟹生态位宽度较为接近,二者生态位重叠指数高达0.837 5,推测中华鲎幼鲎迁移运动特征可能与短指和尚蟹重叠处较多,二者在饵料选择及栖息地选择中亦具有相似性。中华鲎幼鲎常以软体生物为食[30],食源不易获取情况下幼鲎会主动摄食有机质和藻类[2],其分布受潮间带坡度和地形指数等环境因子影响[31]。夏季水温较高时短指和尚蟹栖息于北部湾沿岸成为夏季光滩优势种,滤食泥沙中藻类[32]。各物种间生态位重叠指数在一定程度上反映生态系统复杂性,而生态系统复杂性有利于维护系统稳定性。本研究结果显示,与中华鲎幼鲎生态位重叠指数低于0.6的实验样本高达92.31%,体现多数样本在生活型、摄食选择及底栖群落中生态角色存有差异,各取样地生态系统具有一定程度稳定性。
4.2 中华鲎幼鲎种间联结分析
研究结果显示,中华鲎幼鲎所处群落种群总体关联性呈负联结性,且各物种间存在相互排斥和生态位分离现象。种间联结计算时,当取样地面积较小而物种种类较多时,会增加数量较少物种在计算结果中的比重,从而使AC值偏高[7]。调查期间各取样地物种组成差异较大,故本研究计算得到的AC值可能偏低,据此引入匹配系数以判别中华鲎幼鲎与其所处生境内主要优势种关联强弱。本研究OI和DI均显示中华鲎幼鲎与丽文蛤、南海毛满月蛤及短指和尚蟹关联性较显著(OI≥0.83,DI≥0.83)(图3,图4),表明此三者与中华鲎幼鲎具有相似资源利用形式,对栖息地生境改变具有相似适应性。Pearson相关系数测定中华鲎幼鲎与短指和尚蟹及小翼拟蟹守螺呈正联结性,表明三者具有相似适生区。Spearman秩相关系数不拘泥研究样本的空间分布形式[33],有更高检测灵敏度,结果显示中华鲎幼鲎同短指和尚蟹关联性更强。原因是中华鲎和蟹类的潜在食源均包括虾类[2,34],提升了二者共同出现几率。自然生境内种间竞争可影响群落结构组成、物种丰富度、生物多样性及群落动态[7],下一步可通过扩展时空范围探索中华鲎幼鲎与滨海湿地生物群落演替规律,进一步分析中华鲎幼鲎种群与其他生物群落间相互作用机制,深入理解其濒危机制。
4.3 中华鲎幼鲎栖息地种间竞争动态模拟分析
中华鲎生活史各时期均对水温、水质、底质类型等栖息地特征有所要求[35]。北部湾海域是中华鲎最理想的栖息地[36],2009年西背岭地区调查共发现幼鲎1 357只[37],本研究同地区调查发现幼鲎19只且最大容量仅102只,显示中华鲎幼鲎种群随时间推移出现变化。为直观体现模型模拟结构变化动态,作图时调整各取样地模拟动态时长(图7),显示中华鲎幼鲎与短指和尚蟹在西背岭和三娘湾地区呈此消彼长状态,与短指和尚蟹常成群行动有关。中华鲎幼鲎种群规模减小时能够释放大量共享资源,短指和尚蟹则集群活动迅速占领栖息空间,次年短指和尚蟹在中华鲎成鲎上岸产卵前率先进入繁殖期,进一步提升其在二者间的种间竞争力,据此推测数年后二者种间竞争对中华鲎种群规模影响较显著。竹林盐场地区显示中华鲎幼鲎与短指和尚蟹数量均骤降至极低水平,与该地区受生境破坏和过度捕捞导致滩涂底栖生物资源逐年下降有关。
种间竞争模型模拟种群动态变化显示模拟结果受到最大容量、固有增长率及竞争系数影响。固有增长率能够影响种群规模发展方向,图7显示西背岭、竹林盐场、三娘湾地区中华鲎幼鲎数量均减少,故通过提升中华鲎幼鲎固有增长率至0以上将可能使其摆脱短指和尚蟹种间竞争影响,使种群规模走势快速趋于最大容量。模拟结果显示,中华鲎幼鲎与短指和尚蟹在三娘湾地区可能在较短时期内通过种群内部剧烈竞争达到新的平衡状态,后续可针对中华鲎幼鲎与短指和尚蟹种间竞争在时间相采用常微分方程探讨二者竞争的拓扑结构,并拓展应用于中华鲎与其他大型底栖生物种间关系研究。
物种灭绝是生物多样性损失最敏感的指标之一,由不同地理种群逐个消失造成。本研究理论分析表明,中华鲎幼鲎对栖息环境适应性较强,与现实中华鲎濒危现状相悖,主要原因在于栖息地生境恶化以及捕捞采血或食用等人类干扰[38],因此建议将中华鲎作为未来滨海湿地生物多样性保护和管理的旗舰物种。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
中国农村水利水电(2021年11期)2021-11-29
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
热带农业科学(2019年8期)2019-09-19
中国科技纵横(2016年15期)2016-12-29
