
稻蟹共生系统温室气体排放特征及其影响因素
2022-09-25 10:41张怡彬徐洋王洪媛王绍蓬翟丽梅刘宏斌
农业资源与环境学报 2022年5期
张怡彬,徐洋,王洪媛,王绍蓬,翟丽梅,刘宏斌
(中国农业科学院农业资源与农业区划研究所∕农业农村部面源污染控制重点实验室,北京 100081)
由温室气体排放引起的全球变暖已成为当今全球性的环境问题。氧化亚氮(N2O)和甲烷(CH4)是两类主要的温室气体,其增温潜势分别是CO2的265 倍和28 倍[1-2]。农业活动产生的N2O 和CH4分别约占人类活动排放的50%和60%[3]。稻田生态系统不仅在保证我国粮食安全上发挥着重要作用,同时也是N2O和CH4的重要排放源[4-5]。稻田N2O 排放量占氮肥总输入的2.3%[6],且全球11%的人为CH4产生量来自稻田[7]。因此,降低稻田温室气体排放对减缓全球温室效应具有重要贡献。
稻田综合种养是一种稻田生态系统良性循环的高效养殖模式,一般采用水稻+水产∕水禽互利共生[8],实现“一水两用、一田双收、稳粮增效”[9-10]。近年来在国家的大力支持和推广下稻田综合种类模式得到迅速发展,2019年底全国稻田种养面积达到233万hm2[11]。目前,稻田综合种养模式对温室气体排放的影响研究主要集中在稻鸭、稻鱼以及稻虾等共生系统[12-17]。与水稻单作相比,稻鸭共生表现出对N2O排放的促进作用和对CH4排放的抑制作用[13,18];稻鱼共生系统表现出对N2O的抑制作用和对CH4排放的促进作用[15,19];而稻虾轮作系统对温室气体排放的影响则存在较大争议,有研究认为,稻虾轮作系统会增加N2O排放,降低CH4排放[14],但也有研究表明,稻虾轮作对N2O 和CH4均具有减排效果[16]。稻蟹共生是我国东北地区的典型综合种养模式,占我国稻田综合种养面积的5.94%[11]。然而,目前关于稻蟹共生系统对稻田温室气体排放的影响研究还较少。
因此,本研究选择我国东北地区典型的稻蟹共生模式,以辽宁省盐碱地利用研究所实验基地为依托,设置持续淹水水稻单作、晒田水稻单作和稻蟹共生3种模式,开展不同稻田生态系统下N2O 和CH4排放特征研究,对比分析稻蟹共生与不同水分管理措施水稻单作系统之间的温室气体排放差异性特征及关键环境因子,以期为阐明稻蟹共生模式对稻田系统温室气体排放的影响特征提供依据。
1 材料与方法
1.1 试验地点
本研究田间试验在辽宁省盐碱地利用研究所实验基地开展。该基地位于辽宁省盘锦市大洼区唐家镇刘家村(122°10′ E,41°02′ N),属北温带亚湿润区大陆性季风气候,四季分明,雨量充沛,无霜期约175 d,年平均气温8.3 ℃,年平均降水量645 mm,降雨多集中在7—8 月。试验地土壤类型为盐碱土,土壤基础理化性质:有机质含量10.16 g·kg-1,全氮含量0.51 g·kg-1,全磷含量9.23 g·kg-1,全钾含量25.61 g·kg-1,土壤容重1.34 g·cm-3,pH为7.93。
1.2 试验设计
采用微区试验的方法,共设置持续淹水水稻单作(RM-F)、晒田水稻单作(RM-D)和稻蟹共生(RC)3个处理。其中,RM-F 和RM-D 处理均设置3 个重复(2个处理共6个小区),RC处理设置4个重复,试验共10 个小区,完全随机排列。所有小区试验开始前进行统一的常规田间操作,基础土壤一致,小区面积为24 m(26 m×4 m)。
所有处理化肥用量相等,均为N 210 kg·hm-2、P2O596.4 kg·hm-2、K2O 77.1 kg·hm-2。肥料类型为复合肥(28-15-12)和尿素(N 46%)。根据当地施肥习惯,水稻单作处理(RM-F和RM-D)复合肥(642.9 kg·hm-2)作为基肥一次性投入,尿素(65.2 kg·hm-2)作为蘖肥投入,基肥∶蘖肥为8∶2;稻蟹共生处理所有肥料作为基肥一次性投入;所有处理灌水深度一致,苗期田面水位为5~10 cm,分蘖期为10~15 cm。RM-D 处理晒田7~10 d,其余处理无晒田过程。水稻的插秧与收获时间分别为2019 年5 月31 日和2019 年10 月14日。水稻品种为盐丰47,河蟹品种为中华绒螯蟹,投入蟹苗为大眼幼体,每个小区的蟹苗投入量为1 800只,投入时间为2019 年6 月1 日,收获时间为2019 年10月8日。整个生育期,每个小区的螃蟹饲料投入量为2.33 kg,饲料含氮量为3.7%。田面水的取样频率与气体采集时间一致,土壤样品的采集时间分别为试验前和试验结束后。
1.3 温室气体的采集与测定
温室气体(N2O、CH4)采用密闭静态箱原位采集,用气相色谱法测定排放通量[20]。取气装置是由PVC管制成的方箱,方箱尺寸为长60 cm、宽40 cm、高120 cm。方箱外面用锡箔纸包裹一层海绵,以避免阳光照射造成箱体内温度的变化。采样前用方箱直接罩住4株水稻植株,底部插入土壤中固定,利用田面水密封作用形成密闭环境。在施肥后的1、2、3、4、5、7 d取样,以后每隔7 d 采集1 次气体。需要说明的是,由于基地管理问题,8 月24 日—9 月24 日期间未能按计划进行气体样品采集,可能会对CH4和N2O 排放总量有一定的影响,但不会影响各处理间的差异性。采样口位于箱体顶部,每次取样在上午9:00—11:00进行,盖上箱后分别在0、10、20、30 min 采集气体100 mL 于锡箔采气袋中,每次采气均记录温度和大气压,采集的气体用气相色谱仪(Agilent 7890A)测定N2O和CH4的浓度,气体浓度通过标准气体和待测气体峰面积进行计算。具体计算公式[21]如下:
(1)气体排放通量:

式中:F为气体排放通量,N2O 的单位为μg·m-2·h-1,CH4的单位为mg·m-2·h-1;ρ为标准状态下气体密度,kg·m-3;V为采样箱体积,m3;A为采样箱覆盖面积,m2;Δc∕Δt为单位时间内采样箱内气体浓度变化率;T为采样过程中箱内的平均温度,℃。
(2)气体累积排放量:

式中:M为气体累积排放量,kg·hm-2;i为采样次数;t为采样时间,d;100为单位转换系数。
(3)全球温室气体增温潜势(GWP):

式中:GWP为全球增温潜势,t·hm-(2以CO2当量计);为总N2O 排放量,kg·hm-2(以N2O 计)为总CH4排放量,kg·hm-(2以CH4计);265 指N2O 的增温潜势是CO2的265 倍;28 指CH4的增温潜势是CO2的28倍。
1.4 田面水指标的测定
每次测定温室气体排放通量的同时进行田面水样品采集,并进行田面水水深(SD)、水温(ST)、溶解氧(DO)、氧化还原电位(Eh)、pH 等指标的测定。含量:田面水首先用滤纸过滤,然后用流动分析仪(AA3)测定;ST、DO、Eh 及pH:采用水质多功能参数分析仪(Professional plus YSI)测定。
1.5 土壤样品的测定
分别在水稻插秧前和收获后,取样测定土壤理化指标[22]。其中,有机质(SOM)采用重铬酸钾容量法-外加热法测定;全氮(TN)采用凯氏定氮法测定;全磷(TP)采用NaOH 熔融-钼锑抗比色法测定;全钾(TP)采用NaOH熔融-火焰光度法测定采用0.01mol·L-1CaCl2浸提-流动分析仪测定;微生物量碳(MBC)、微生物量氮(MBN)采用氯仿熏蒸-K2SO4浸提,Vario-TOC自动分析仪测定[23]。
1.6 数据处理
所有数据均采用Microsoft Excel 2016、Canoco 5和AI 软件处理并制图,并采用SPSS 19.0 软件中单因素方差分析(One-way ANOVA)的Duncan 检验统计处理之间的差异,显著水平为P<0.05。
2 结果与分析
2.1 监测期自然条件
监测期间,采样时间段的最高温度为30.3 ℃,最低温度为10.8 ℃(图1)。降雨主要集中在7 月底和8月初,最大降雨量达到79.5 mm;生育期累积降雨量为539.7 mm,占全年降水量的87.6%。其间,由于长时间未下雨,田面水深下降,分别在6月1日(插秧期,灌水量0.5 m3)、6 月18 日(追肥期,灌水量0.8 m3)和7月24日(抽穗期,灌水量1.0 m3)进行灌溉以补充水位。

图1 监测期的降雨与日平均气温Figure 1 Rainfall and daily mean temperature during monitoring period
2.2 N2O排放特征
水稻全生育期3 个不同处理稻田N2O 排放表现出相同的变化趋势,RM-F 处理的排放通量在追肥期相对较高(图2)。基肥期,三个处理都没有出现明显峰值,排放量均较低,N2O 排放通量基本处于15.2~35.0 μg·m-2·h-1。追肥后,水稻单作和稻蟹共生处理均出现了一个波峰,RM-F 处理和RC 处理的最大峰值均出现在追肥后的第2 天,分别为99.5 μg·m-2·h-1和55.1 μg·m-2·h-1,RM-D 处理的最大峰值则出现在追肥后的第3天,为50.8 μg·m-2·h-1。值得注意的是,从追肥到7月24日(灌水),稻田N2O 排放一直处于较高的波动状态,N2O 排放量占总排放量的57.6%~59.9%。较高排放水平的维持与田面水深的持续下降密切相关。之后N2O 排放基本稳定在较低水平,这一方面可能是由于前期投入的氮肥和饲料已被大量消耗;另一方面主要是因为后期为保证螃蟹的快速生长,田面水维持了较深的状态,抑制了N2O 排放。综上,研究区域水稻生长前期是稻田N2O 排放的主要排放时期。

图2 稻田N2O排放通量特征Figure 2 Characteristics of N2O emission from paddy field
2.3 CH4排放特征
水稻全生育期3个不同处理稻田CH4排放亦表现出相同的变化趋势,并在排放高峰期表现出显著差异(图3)。在水稻生长前期(抽穗期之前),3 个处理的CH4排放量均较低,CH4排放通量基本处于0.26~24.5 mg·m-2·h-1。在水稻生长进入抽穗期后,CH4排放量占水稻整个生育期排放量的88.4%~94.3%,CH4排放出现了2 个高峰。RM-D 处理在第一个排放高峰期CH4排放通量最高,为87.1 mg·m-2·h-1;RM-F 处理的略低,为73.5 mg·m-2·h-1;RC 处理的最低,仅为15.7 mg·m-2·h-1。CH4排放通量第二个高峰期出现在8 月21 日(进入雨季),此时RC 处理最高,为37.9 mg·m-2·h-1,而RM-D处理最低,仅为10.8 mg·m-2·h-1。
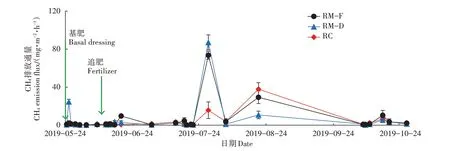
图3 稻田CH4排放通量特征Figure 3 Characteristics of CH4 emission flux from paddy field
2.4 温室气体累积排放量及增温潜势
不同稻田系统水稻全生育期的温室气体排放结果(表1)表明,RC 处理的N2O 累积排放量显著低于RM-F 处理23.9%(P<0.05),低于RM-D 处理16.7%(P>0.05)。RC 处理的CH4累积排放量低于RM-F 处理13.5%(P>0.05),却显著高于RM-D 处理34.0%(P<0.05)。这表明稻蟹共生模式能够降低稻田生态系统N2O 排放,与持续淹水水稻单作模式相比,其CH4累积排放量降低,但相比于晒田模式,CH4累积排放量增加。不同处理的增温潜势表现为RM-F>RC>RM-D,与持续淹水单作模式相比,稻蟹共生模式增温潜势降低13.6%;而与晒田单作模式相比,稻蟹共生模式增温潜势增加32.6%,但差异不显著。可见,相比于种养模式的改变,水分管理对增温潜势的调控作用更强。

表1 稻田系统温室气体累积排放量及增温潜势Table 1 Cumulative greenhouse gas emissions and warming potential of rice field system
2.5 环境因子与温室气体排放的相关性
Pearson 相关性分析(表2)表明,N2O 排放与田面水水深和水温呈显著负相关,与pH呈显著正相关;而CH4排放与田面水水深和水温呈极显著正相关(P<0.01),与NO-3-N 含量和pH 呈显著负相关。可见,田面水水深、水温以及pH 等环境因子对N2O 和CH4排放的影响作用相反,这可能是水稻生长期N2O 和CH4排放表现出此消彼长关系的主要原因。
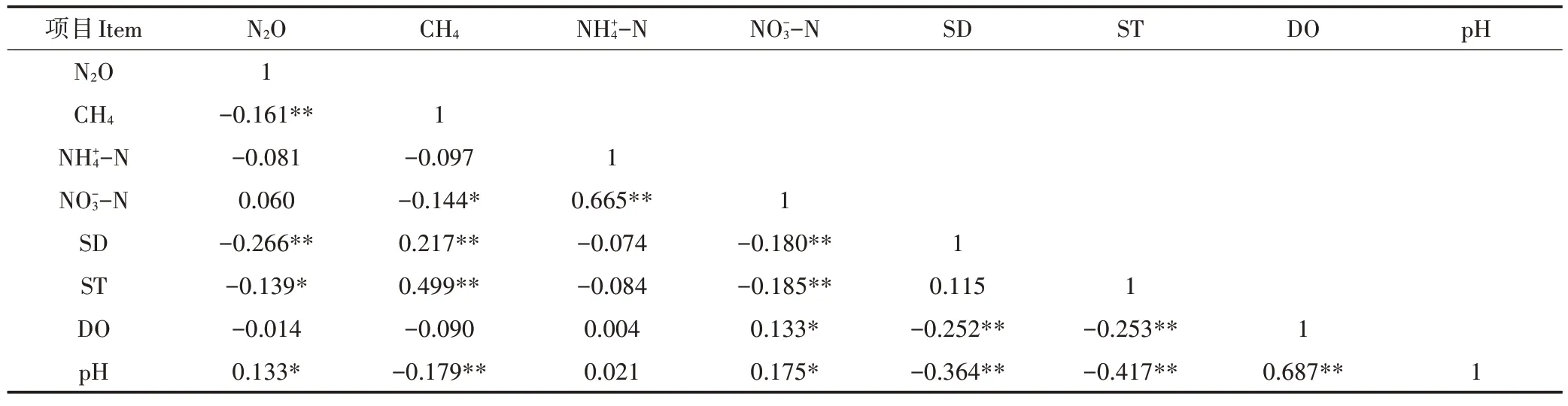
表2 温室气体排放与环境因子的相关性Table 2 Correlation between greenhouse gases emission and environmental factors
稻田土壤理化指标与温室气体排放之间的冗余分析(图4)表明,土壤NH+4-N、NO-3-N、SOM、MBC、MBN、TN、TP、TK、pH 等土壤生物化学指标对水稻生长期温室气体排放影响的解释率可达83.10%。第一主成分和第二主成分的贡献率分别达52.38%和30.72%。其中,pH(33.3%,F=4.0,P=0.01)和NO-3-N(20.0%,F=3.0,P=0.042)对温室气体排放的影响最大。

图4 土壤生物化学特征与温室气体累积排放量的冗余分析Figure 4 Redundancy analysis of soil biochemical characteristics and cumulative greenhouse gas emissions
稻田田面水理化指标与温室气体排放之间的冗余分析(图5)表明,田面水NH4+-N、NO3--N、DO、SD、ST、Eh 和pH 等环境因子对水稻生长期温室气体排放影响的解释率可达87.25%,第一主成分和第二主成分的贡献率分别达54.88% 和32.37%。其中,DO(35.6%,F=4.4,P=0.012)和pH(26.7%,F=5.0,P=0.014)对温室气体排放的影响最大。
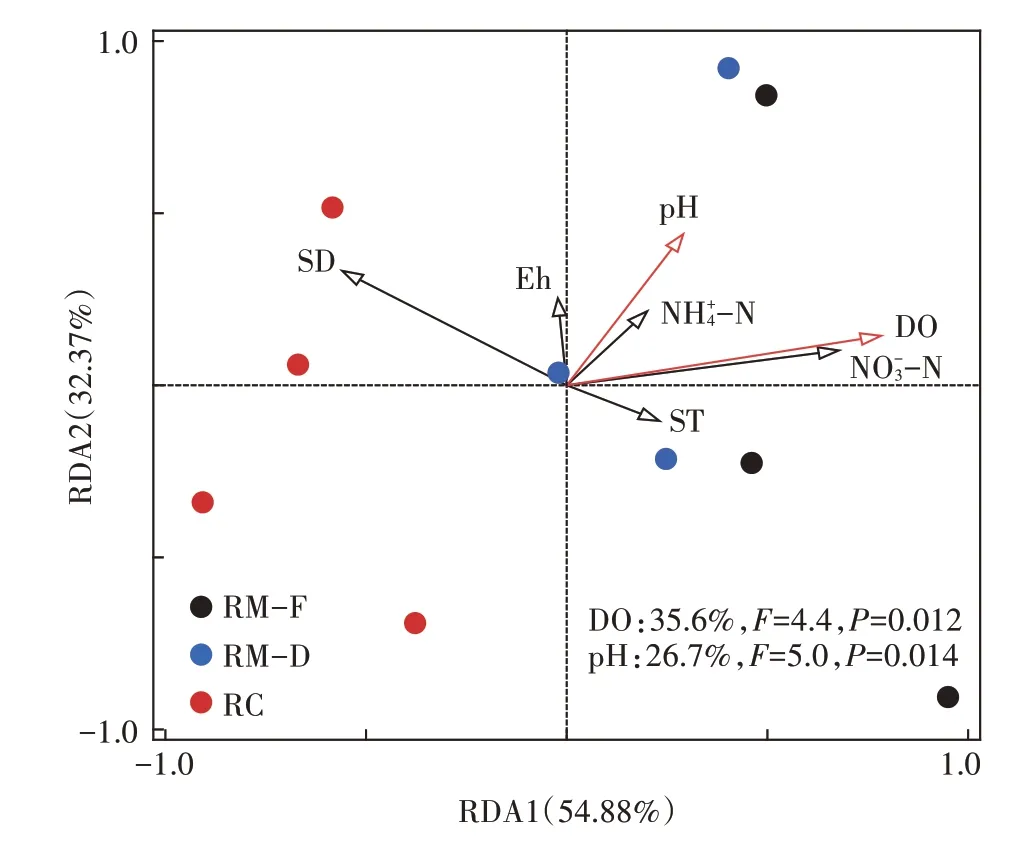
图5 田面水理化性质与温室气体排放通量的冗余分析Figure 5 Redundancy analysis of physical and chemical properties of field surface water and greenhouse gas emission flux
3 讨论
3.1 共生系统对N2O排放的影响
稻田N2O 排放受外源碳氮供应水平的影响[24],N2O 排放峰值主要发生在施肥后[25]。本研究结果表明稻田系统N2O 排放主要发生在追肥前期,基肥期和追肥后期N2O 排放一直处于较低水平。基肥期N2O排放处于较低水平的主要原因是稻田系统淹水状态下87%的N2O 通过水稻植株排放[26]。基肥期秧苗移栽前,N2O 排放只能通过土壤向水体扩散,再向空气排放;而移栽初期,秧苗还未深入扎根,根系和通气组织尚未生长成熟,对N2O 的输出作用较小[3]。追肥期前期,水稻植株已生长良好,开始分蘖,通气组织发达,为N2O 提供了良好的排放通道,导致N2O 排放量急剧增加,形成N2O 排放峰。追肥后期,特别是7 月底之后,N2O 排放量一直呈现较低水平,主要原因可能是此时试验基地进入高温多雨季节,大暴雨造成田面水位的急剧上升,形成严格厌氧环境,抑制了稻田N2O 的产生。并且,追肥前期稻田田面水及土壤中的含量较高,能够为N2O的产生提供充足的物质基础,而追肥后期田面水及土壤中养分含量较低,减少了N2O 的产生。侯玉兰[3]和黄树辉等[27]的研究也得到了相似的结果。值得注意的是,稻蟹共生处理在整个水稻生长周期出现了多个峰值,这些峰值主要发生在饲料投喂后。据此推测,N2O 排放峰值出现的原因可能是螃蟹饲料的投入增加了土壤中易利用碳氮养分的含量,增强了土壤微生物硝化反硝化过程N2O的产生,从而促进了N2O的排放[14]。
与水稻单作处理相比,稻蟹共生处理能够有效降低稻田系统N2O 排放,减排量可达16.7%~23.9%。稻蟹共生模式下稻田N2O 排放降低的原因主要有两方面:一是螃蟹捕食水中杂草和浮萍绿藻等生物,杂草减少限制了淹水稻田N2O 通过植株通气组织向大气排放,而浮萍绿藻的减少会抑制光合作用,降低水中DO 含量,强还原环境有利于N2的产生,从而降低N2O排放[13];另一方面,随着螃蟹个体的生长,为保证螃蟹的生存空间,维持了较高的水位,同时,螃蟹通过呼吸作用大量消耗田面水中的氧气,使土壤处于严格厌氧环境,促进了N2O 向N2的转化,进而抑制了N2O 的产生与排放[3,27]。
不同处理的水分管理方式和种养模式不同,其对稻田系统田面水∕土壤养分含量、pH、DO 等关键环境因子的作用强度也不同,导致各处理N2O 排放量具有差异。水分管理措施是影响农田N2O 和CH4排放的重要因素之一[28]。本研究表明,水稻单作系统晒田过程能够降低8.7%的N2O 排放量,而且水稻生长过程中水位高度与N2O 排放通量之间呈极显著负相关。这主要是由于土壤水分变化会直接影响农田系统中硝化、反硝化和N2O 还原之间的平衡,从而影响N2O排放[29]。土壤pH 可通过影响反硝化和硝化微生物的活性及相应的氮素转化过程间接影响N2O 的释放[30-31]。本研究表明稻田N2O排放与pH呈正相关,黄耀等[32]和LI 等[33]的研究也得出类似结论。LI 等[33]认为,淹水期N2O 排放量与pH 呈显著的相关性,排水后则无相关性;黄耀等[32]研究发现当pH 在5.6~8.6 之间时,pH和N2O排放呈显著正相关。
已有研究表明,N2O的排放会受到田面水中和含量的显著影响[21],然而在本研究中却未表现出显著相关性。这可能是由于本研究所处的东北稻区在施入基肥后会泡田7~10 d,然后再开始插秧,此时田面水中含量在经历过施肥后的高峰后,回归到相对较低的平稳期;而水田87%的N2O 通过水稻植株排放[26]。因此,虽然田面水和含量较高,但是由于缺少水稻植株通气组织的传输,基肥期的N2O 排放量较低,最终导致田面水中与N2O 排放之间无显著的线性相关。
3.2 共生系统对CH4排放的影响
稻田CH4排放是产甲烷菌产生CH4和甲烷氧化菌最终氧化为CO2的净效应[34]。本研究结果表明稻田系统CH4排放主要发生在水稻追肥后期,基肥期和追肥前期CH4排放一直处于较低水平。已有研究表明稻田系统CH4从产生到排放,主要通过水稻植株的运输、扩散和蒸腾作用这3 个途径[35-36]。因此,稻田CH4排放的整个过程均受到水稻生长状况、温度、土壤特性等生物和非生物因素的影响。在基肥期和追肥前期,稻田未进行水稻移栽或处于移栽初期,秧苗尚未深入扎根,根系和通气组织尚不发达,且这段时期温度较低,植株生长缓慢,土壤微生物活性较低,这些都可能抑制稻田CH4的排放。另外,在追肥前期,田面水位较低,未形成严格的厌氧环境,也不利于CH4的产生。水分对稻田CH4排放起到决定性的作用[37]。水分通过影响土壤氧化还原电位、氧气供给、土壤微生物(最主要的是产甲烷菌)活性及CH4扩散速率来实现对稻田CH4排放的调控[38]。追肥后期,试验地进入雨季,稻田系统CH4排放的两个峰值分别出现在7 月和8 月,上述现象产生的原因可能是试验区CH4排放受降雨影响较大,两次峰值均出现在强降雨之后,田间水深随着降雨量的增大而升高,从而促进了CH4的排放。这也与Pearson 相关性分析得到的田面水深与CH4排放极显著相关的结果一致。同时,试验地的雨季伴随着高温天气,温度的升高有利于CH4的排放。已有研究表明,CH4排放与大气温度呈正相关[39]。水稻进入抽穗期后,植株生命力最旺盛,CH4主要通过植株通气组织向大气排放,故此时CH4排放量较高[40]。综上,在本研究区域,水稻生长中期是稻田CH4排放的主要排放时期,同时降雨引起的水位上升和DO 降低可能是造成CH4排放通量剧增的重要原因。
水分管理是影响种养结合稻田CH4排放的重要因素之一。与持续淹水水稻单作处理相比,稻蟹共生处理能够有效降低稻田系统CH4排放,减排量可达13.5%。以往关于稻鸭共生和稻虾轮作的研究也得出相似的结论[21],稻鸭共生可以降低稻田系统18%~25%的CH4排放[21];稻虾轮作可降低18%~41%的CH4排放[14,16]。共生系统CH4排放量降低的原因,可能是动物活动促进了田面水和稻田土壤的交换,提高了甲烷氧化菌的活性,增强了CH4氧化速率。而与晒田水稻单作处理相比,本研究的稻蟹共生系统增加了34.0%的CH4排放。主要原因是,相比晒田处理,稻蟹共生稻田一直处于持续淹水的严格厌氧环境,产甲烷菌的活性更强,从而增加了CH4排放。BHATTACHARYYA 等[15]和DATTA 等[19]在稻鱼共生模式的研究中也得到了相同的结论。本研究表明,田面水水深、水温、pH 以及NO-3-N 含量是影响CH4排放的主要因素。这些因素主要受水分管理的影响,不同水分管理措施会改变土壤含氧量,影响产甲烷菌活性,调控稻田CH4排放。相比于持续淹水,干湿交替可显著降低季节性CH4排放[7,41-42]。
目前,关于稻蟹共生模式对稻田系统温室气体排放的影响研究较少。WANG 等[43]的研究表明,与常规水稻单作相比,稻蟹共生可以降低19.7%~28.2%的N2O排放,增加29.2%~36.8%的CH4排放量,这与本研究结果一致。本研究结果表明,与晒田水稻单作处理相比,稻蟹共生模式能够降低稻田系统16.7%的N2O排放,增加34.0%的CH4排放。而与持续淹水水稻单作处理相比,稻蟹共生模式能够降低稻田系统23.9%的N2O 排放和13.5%的CH4排放。可见,在同样的水分管理方式下,稻蟹共生能够有效降低稻田系统N2O和CH4排放。同时,整个水稻生长期N2O 和CH4排放表现出互为消长的关系。有研究认为导致稻田N2O和CH4排放互为消长的关键因素是土壤水分状况[44-45]。追肥前期,田面水位较低,N2O 排放通量高,而CH4排放通量低;进入高温雨季,稻田水位明显升高,此时N2O 排放通量降低,而CH4排放通量明显增加。田面水水深、水温和pH 以及土壤NO-3-N 含量和pH 是导致N2O 与CH4排放互为消长的主要因素。Pearson 相关性分析表明田面水深和水温的升高会抑制N2O排放,但会促进CH4排放;土壤NO-3-N含量、pH是田间温室气体排放的主要影响因子。
4 结论
(1)在相同的水分管理方式下,稻蟹共生能够降低稻田系统N2O 和CH4排放;而相比于晒田水稻单作处理,持续淹水的稻蟹共生模式会降低N2O 排放、促进CH4排放。整体而言,与持续淹水处理相比,稻蟹共生模式可降低13.6%的增温潜势;与晒田水稻单作处理相比,稻蟹共生模式则会增加32.6%的增温潜势。这表明相比于种养模式的改变,水分管理方式对增温潜势的影响作用更大。
(2)稻田系统N2O 和CH4的排放呈互为消长关系,主要受田面水水深、水温和pH 以及土壤NO-3-N含量和pH的影响。田面水水深和水温的增加会抑制N2O 排放,但会促进CH4排放;土壤NO-3-N 含量和pH是田间温室气体排放的主要影响因素。
猜你喜欢
河南水利与南水北调(2022年7期)2022-08-18
磷肥与复肥(2022年6期)2022-08-02
——以长江中游单季稻田为例
湖北农业科学(2022年10期)2022-06-14
农业灾害研究(2022年1期)2022-05-07
核农学报(2021年11期)2022-01-04
腐植酸(2021年2期)2021-12-04
农业与技术(2021年12期)2021-07-02
暴雨灾害(2021年2期)2021-04-02
干旱地区农业研究(2020年2期)2020-06-10
山东农业科学(2019年11期)2019-12-24
