
魔芋不同生理状态的愈伤组织和丛生芽内源激素及抗氧化酶研究
2022-09-19 12:29蔡阳光段龙飞覃剑锋郭邦利陈国爱张百忍
西南大学学报(自然科学版) 2022年9期
蔡阳光,段龙飞,覃剑锋,郭邦利,陈国爱,张百忍
安康市农业科学研究院, 陕西 安康 725021
植物的生长过程是植物内源各激素水平间的动态平衡过程, 内源激素对植物愈伤组织生长和分化有调控作用[1]. 但在诱导植物组织分化过程中, 内外源激素之间存在相互作用, 外源激素与内源激素的种类和水平有着密切的关系, 并通过多种机理实现相互作用, 一起构成了一个非常精细而复杂的调控网络[2-4]. 通过外源激素的适时适量运用, 可使植物体内的生理生化反应以及激素水平和种类发生显著的变化, 进而调控作物的源库关系[5-7]. 王京伟等[8]研究发现, 内外源激素协同调控山丹小鳞茎的分化方向, 且在培养过程中发现, 添加的外源激素不同, 小鳞茎的形态变化也不相同, 对应的内源激素水平也发生了一定的变化. 李本波[9]在杜梨愈伤诱导发生的研究中指出, 外源激素和内源激素在杜梨愈伤组织培养中有着较强的相互作用, 添加不同的单一外源激素, 可促进杜梨内源激素的合成, 同时还可以引起激素间的比值发生变化; 且不同外源激素对杜梨愈伤组织内源激素影响存在一定的差异, 内源激素水平或不同内源激素间比值与不添加外源激素的愈伤组织间差异有统计学意义.
魔芋作为地下球茎类作物, 其离体器官发生形式与内源激素变化及平衡水平相关[10]. 吝祥根[11]研究指出, 在细胞分化过程中, 高水平的内源ZR,ZR/IAA比值以及适量的IAA和GA3有利于花魔芋愈伤组织细胞的分化, 而ABA不利于愈伤组织细胞的分化; POD α-淀粉酶和PAL活性增加有利于愈伤组织细胞的再分化, 这些酶可能是以提高细胞的代谢水平以及提供细胞分化所需要的物质和能量的方式, 发挥其对细胞分化的促进作用. 黄丹枫等[12-14]研究指出, 异戊烯基嘌呤类(iPAs)CTK是决定魔芋细胞分化和器官发生的内源激素; 细胞内异戊烯基嘌呤类细胞分裂素质量分数的上升与不定芽发生呈正相关, 培养基中细胞分裂素类物质的种类和浓度, 影响魔芋愈伤组织细胞内iPAs的生物合成, 从而影响魔芋愈伤组织的细胞分化、 器官发生和植株再生; 另外, 内源细胞分裂素iPAs与魔芋不定芽分化正相关, 通过促进遗传物质的合成, 提高CAT,PAL,POX酶活性而起作用, 同时分化培养基中适宜的6-BA,IAA浓度与iPAs,CAT活性正相关, 也促进了魔芋不定芽的分化. 胡建斌等[15]对不同类型的魔芋愈伤组织研究后发现, 只有结构致密且表面呈瘤状的愈伤组织(Ⅲ型)才能诱导形成微型薯, 其他类型的愈伤组织(Ⅰ型, Ⅱ型)则倾向于形成不定芽; 进一步研究两种途径的愈伤组织中的内源激素发现, 赤霉素与茉莉酸的平衡是调控魔芋离体形态建成的主要因素. 魔芋是我国西部山区重要的特色经济作物, 繁殖系数低, 组织培养是快繁和优质种质保存的重要途径. 目前魔芋组培主要经过外植体脱分化和再分化进行, 在脱分化和再分化过程中, 受遗传因素、 培养条件、 生理状态、 激素以及次生代谢物质的影响, 愈伤组织和丛生芽形态多样化且频率不一致, 如褐化、 僵化、 老化和玻璃化等, 会对后续的继代培养和形态建成产生不利影响. 另外对魔芋离体细胞脱分化、 再分化和形态建成的内在机制及其调控机制, 内源激素对魔芋离体再生的影响及其作用机制, 以及各激素间相互影响均需进一步验证[16-17]. 本研究以遗传同质、 生理状态不同的魔芋愈伤组织和丛生芽作为研究对象, 通过测定其内源激素的质量分数以及SOD,POD,CAT酶的活性, 分析其相关性, 探明内源激素对形态建成的影响, 为定向调控奠定基础.
1 材料与方法
1.1 试验材料
杂交魔芋品种“安魔128”, 安康市农业科学研究院自主选育的魔芋品种, 2019年种植于该院示范园魔芋繁育基地, 常规管理, 选取3龄球茎作为外植体组培, 挑选同期不同生长及分化状态的愈伤组织和丛生芽作为检测材料.
1.2 组织培养
试验于2020年4月23日在安康市农业科学研究院魔芋组培中心进行. ① 选取1.21 kg健康无外伤的“安魔128”球茎, 清洗消毒后, 将其均匀切成1 cm3大小, 接种于愈伤组织诱导培养基中: MS+2.0 mg/L 6-BA+0.5 mg/L 2,4-D+0.2 mg/L NAA+4.2 mg/L琼脂+30 g/L蔗糖, pH值为5.8~6.0, 每瓶接种1块, 共227瓶; ② 培养至32 d, 挑选3种不同生长状态的愈伤组织进行内源激素及酶活检测, 每个处理挑选10瓶; ③ 同时挑选质地坚硬、 表面呈颗粒状的愈伤组织, 平均切分成2块, 接种于芽分化诱导培养基中: MS+3.0 mg/L 6-BA+1.0 mg/L KT+0.2 mg/L 2,4-D+0.5 mg/L NAA+4.2mg/L琼脂+20 g/L蔗糖, pH值为5.8~6.0, 每瓶接种1块, 共169瓶; ④ 培养至40 d, 挑选不同分化状态的丛生芽进行内源激素及酶活检测, 每个处理挑选10瓶.
1.3 测定指标及方法
2020年5月24日挑选3种类型愈伤组织进行检测, 每种类型挑选10瓶, 愈伤组织平均9.49 g/个. 2020年7月3号挑选5种类型丛生芽进行检测, 每种类型挑选10瓶, 丛生芽平均7.53 g/个. 采用高效液相色谱法[18-19]测定GA3,ABA质量分数; 采用酸性茚三酮法[20]测定脯氨酸(Pro)质量分数, 硫代巴比妥酸显色法[21]测定MDA质量分数; 采用对邻苯三酚自氧化法[22]测定超氧化物歧化酶(SOD)的活性, 参照陈建勋等[23]的方法测定过氧化物酶(POD)的活性, 紫外分光光度法[24]测定过氧化氢酶(CAT)的活性.
采用酶联免疫分析(ELISA)试剂盒测定6-BA,KT,2,4-D及NAA的质量分数: 取0.5 g经液氮处理后的样品, 充分研磨, 加入PBS, 2 000~3 000 r/min离心20 min, 收集上清液后分装, 留1份待测, 其余保存于4 ℃冰箱备用. 往预先包被6-BA,KT,2,4-D及NAA捕获抗体的微孔中, 依次加入标本、 标准品、 HRP标记的检测抗体, 经过温育并彻底洗涤. 用底物TMB显色, TMB在过氧化物酶的催化下转化成蓝色, 并在酸的作用下转化成最终的黄色. 颜色的深浅和样品中各激素的多少呈正相关. 用酶标仪在450 nm 波长下测定吸光度(OD值), 计算样品浓度, 用标准品做对照计算各指标的质量分数, 每个处理重复测定3次.
1.4 数据统计与分析
用Excel 2003与SPSS 19.0软件进行数据整理与统计分析.
2 结果与分析
2.1 不同类型魔芋愈伤组织生理指标分析
由表1可知, 魔芋愈伤组织类型3的GA3,Pro质量分数及抗氧化酶SOD,CAT活性均显著高于其他2种类型, 同时ABA,MDA质量分数均较低, 表明愈伤组织类型3较类型1和类型2生长更旺盛, 这与图1中的外在表现较一致. 类型3愈伤组织分化效果最好, 质地坚硬松散, 表面有颗粒状, 呈粉绿色; 类型2分化效果良好, 但质地较软呈水渍状, 表面暗红色; 类型1分化不完全, 质地坚硬不松散, 且伴有褐化现象.
从表1中还可以看出, 愈伤组织类型3中4种内源激素质量分数均显著高于类型1和类型2, 且各激素6-BA, KT, 2,4-D, NAA的比例为114∶80∶68∶100, (6-BA+KT+2,4-D)与NAA的比值R=2.62∶1(类型3)<2.71∶1(类型2)<2.89∶1(类型1). 4种内源激素水平间差异有统计学意义, 且当(6-BA+KT+2,4-D)与NAA的比值R≤2.62时, 有利于魔芋球茎愈伤组织的分化. 同时, 在诱导脱分化形成愈伤组织的过程中, 单独使用2,4-D也可诱导愈伤组织的形成, 但形成的愈伤组织较松软, 难分化; 而使用中低浓度的2,4-D配合6-BA(KT)对加速细胞分裂快速形成胚性愈伤具有明显的作用.
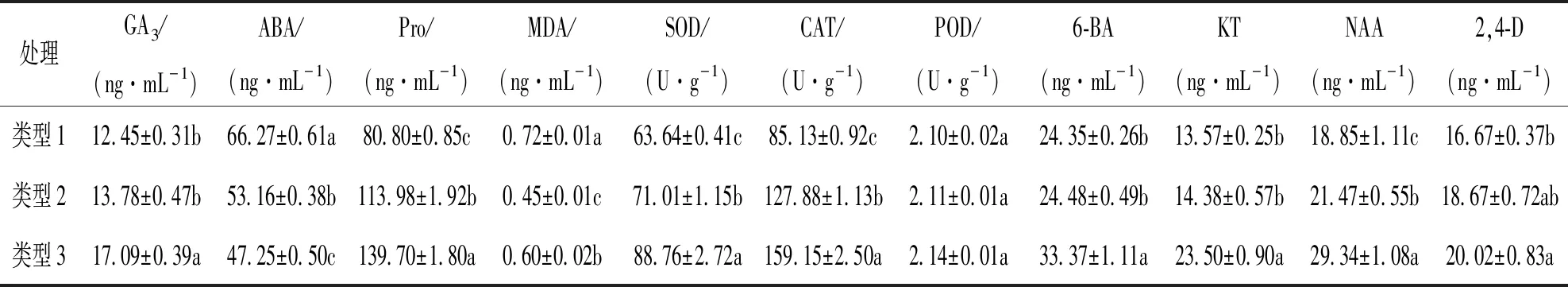
表1 3种类型魔芋愈伤组织生理指标比较

图1 3种不同生长状态的魔芋愈伤组织
2.2 不同类型魔芋丛生芽生理指标分析
由表2可知, 类型3魔芋丛生芽GA3,Pro的质量分数及抗氧化酶活性均较高, 且ABA,MDA质量分数均较低, 较其他类型丛生芽差异有统计学意义. 结合图2丛生芽分化的外在表现, 表明该类型芽处于生理旺盛阶段. 类型1丛生芽由于褐化影响了正常的分化和生长; 类型2丛生芽出现了老化现象, 再生频率低; 类型4丛生芽与类型3相比, GA3,CAT质量分数较低, 导致有效芽分化数相对较少; 类型5丛生芽SOD活性最高, 同时ABA,MDA质量分数也较高, 而GA3,Pro质量分数及CAT活性相对较低, 可能细胞膜受到氧自由基的伤害, 从而形成外在的玻璃化状态以及显示边缘水渍状.
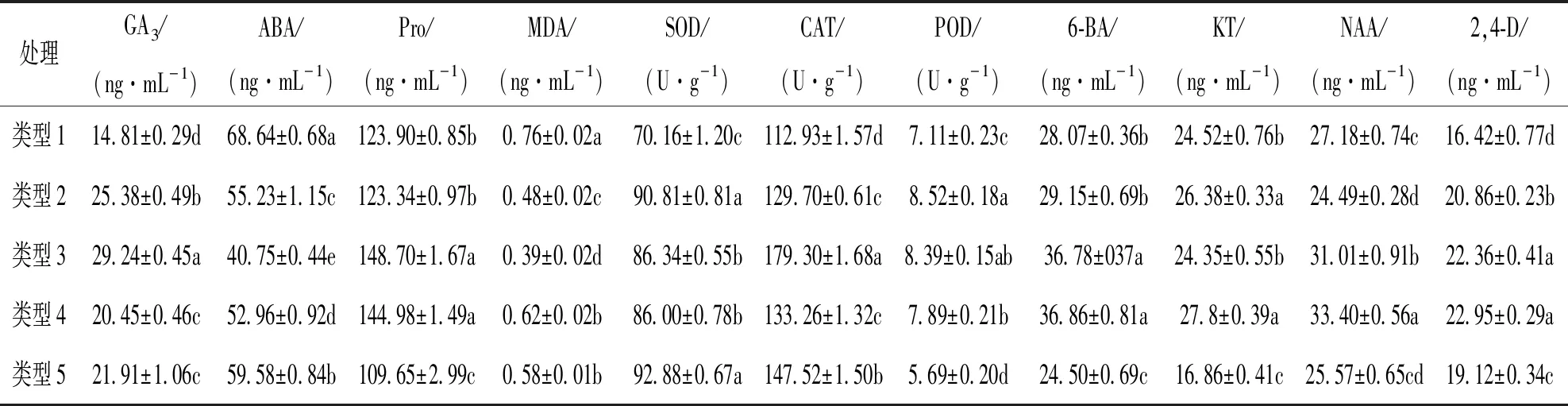
表2 5种类型魔芋丛生芽生理指标比较
由表2还可以了解到, 不同类型魔芋丛生芽内源激素水平有差异, 类型3各激素6-BA, KT, 2,4-D, NAA的比例为119∶79∶72∶100, (6-BA+KT+2,4-D)与NAA的比值R=2.70∶1, 其中2.37∶1(类型5)<2.53∶1(类型1)<2.62∶1(类型4)<2.70∶1(类型3)<3.12∶1(类型2). 说明对于魔芋丛生芽分化而言, 细胞分裂素和生长素总的比值应维持在适宜水平, 比值过高(类型2), 易出现老化芽现象; 比值过低又会导致分化慢, 分化不完整. 另外, 仅KT和NAA差异显著的类型3和类型4, 芽的分化数量和形态也有差异.
在图2中, 类型3芽密, 质地坚硬松散, 芽头呈嫩绿色; 类型4芽密, 呈松散状, 芽头多呈粉红色; 类型1芽少, 质地较硬不松散, 伴有部分褐化; 类型2芽密, 质地较硬不松散, 芽头大多呈浅粉色, 并且芽外形呈花瓣状, 难以形成有效苗; 类型5芽密, 质地偏软不松散, 芽较长且呈白色或透明状, 幼苗细弱且叶片呈水渍状, 即玻璃化.

图2 5种不同生理状态魔芋丛生芽
2.3 魔芋愈伤组织内源激素质量分数及抗氧化酶相关性分析
由表3可知, 内源激素KT和NAA质量分数之间呈极显著负相关, 6-BA和2,4-D质量分数之间呈极显著正相关. 6-BA,2,4-D与POD活性之间呈极显著正相关, 与SOD活性之间呈显著负相关. NAA与SOD,CAT活性之间呈显著正相关, 2,4-D与CAT活性之间呈显著负相关. SOD与CAT之间呈极显著正相关, 而与POD之间呈极显著负相关. ABA,Pro和MDA与衰老和逆境胁迫有关, 与NAA呈极显著正相关, 与KT呈负相关, 较低的NAA和较高的KT水平有利于愈伤组织的增殖.
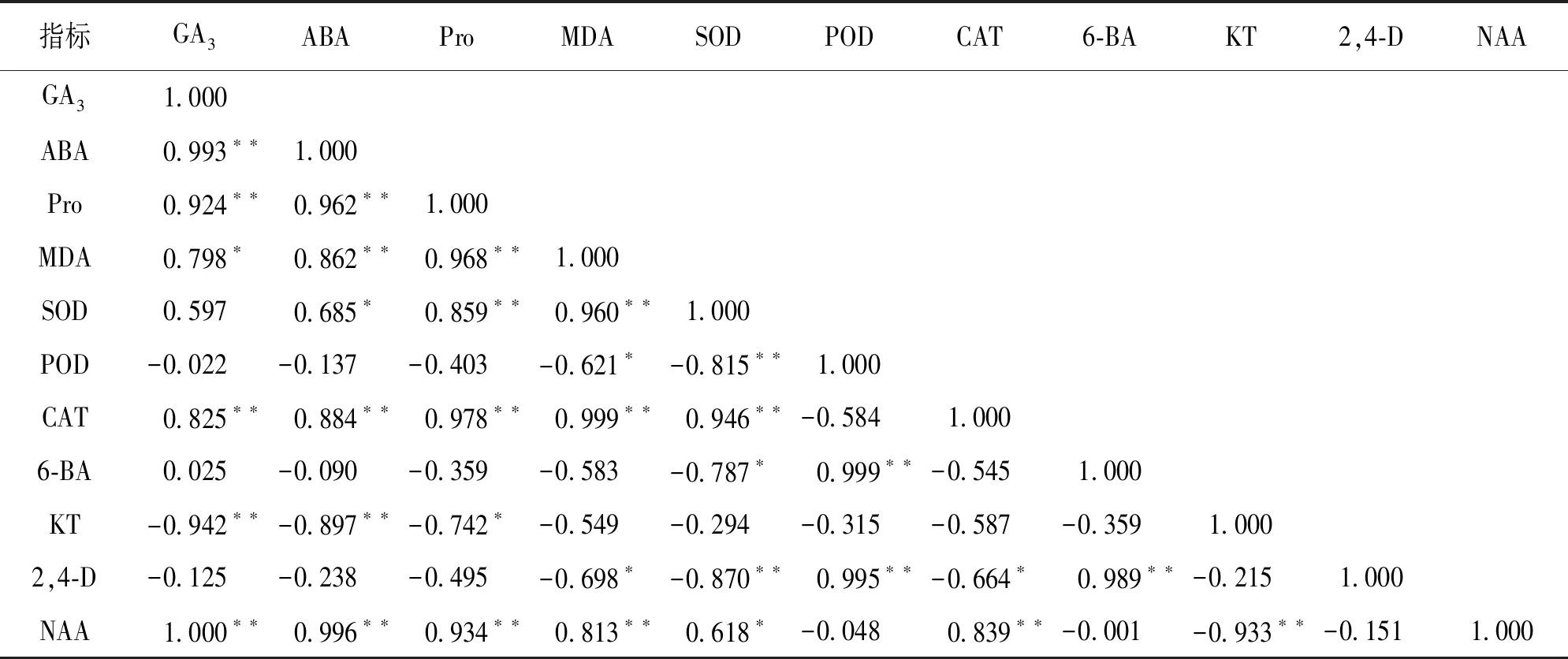
表3 魔芋愈伤组织生理指标的相关性分析
2.4 魔芋丛生芽内源激素质量分数及抗氧化酶相关性分析
由表4可知, 内源激素KT与2,4-D,6-BA之间均呈极显著正相关, 6-BA,KT及2,4-D与SOD,POD活性之间均呈极显著或显著正相关. KT,2,4-D及NAA质量分数与CAT活性之间均呈显著或极显著正相关, 保护酶SOD与CAT,POD活性之间呈显著或极显著正相关. 另外, SOD,CAT,GA3及Pro与MDA之间呈显著或极显著负相关, Pro与CAT之间呈极显著正相关. 表明KT与2,4-D,6-BA之间相互促进, 且内源各激素质量分数的增加会促进相关抗氧化酶活性的增高; 同时抗氧化酶与相关生理物质间存在相互促进与抑制作用. 6-BA与ABA呈极显著正相关, NAA与ABA呈极显著负相关, 较低的6-BA和较高的NAA有利于芽的分化.
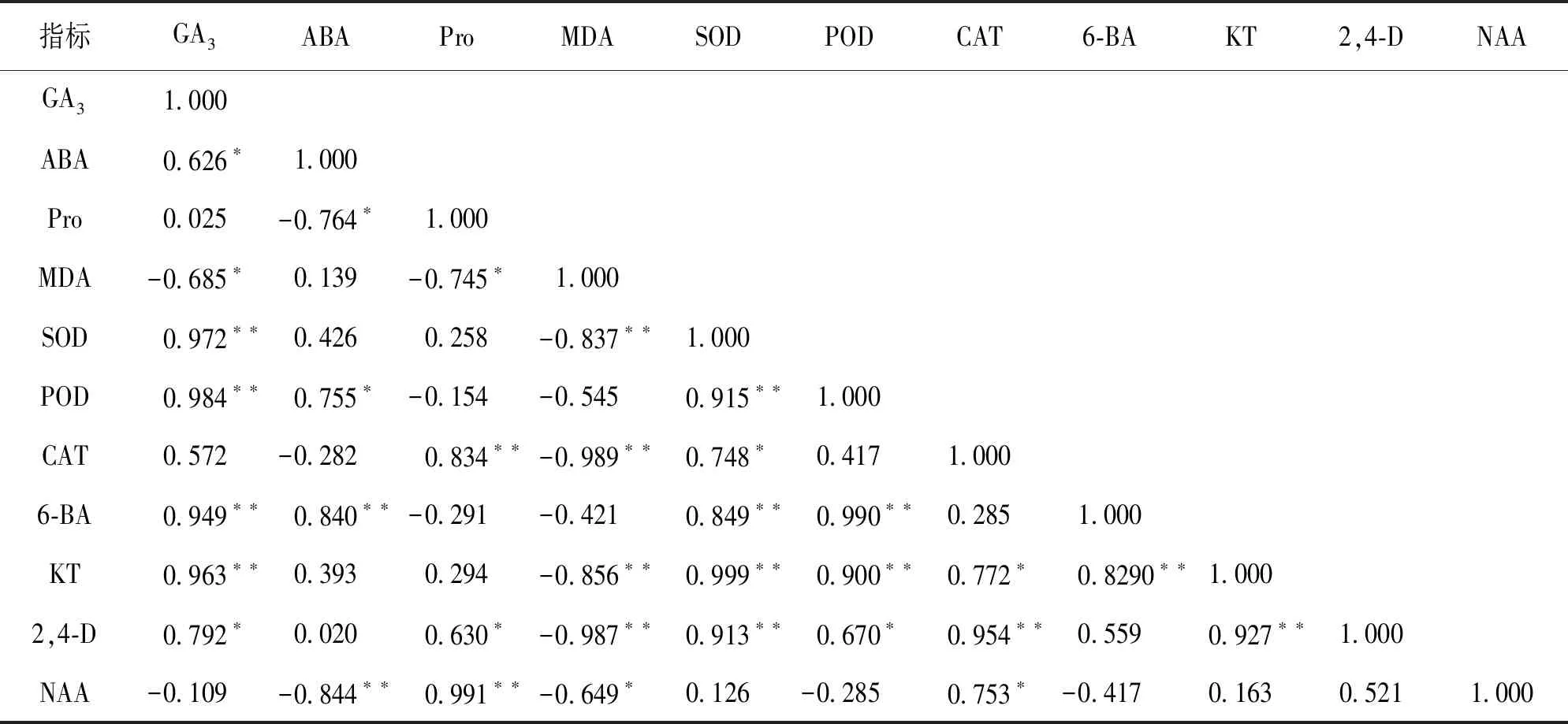
表4 魔芋丛生芽生理指标的相关性分析
3 讨论
刘清等[25]研究指出, 内源激素对水稻愈伤组织的生理表现有着极其重要的影响, 其种类、 质量分数及协调平衡是水稻愈伤组织形成和分化的关键. 合适的浓度和比例能有效促进外植体的脱分化与再分化, 并协同促进细胞的分裂、 分化和生长[26]. 郭敏敏等[27]通过测定棉花愈伤组织中的内源激素后发现, 不同内源激素适宜的浓度和比例调节外植体的脱分化和再分化, 各种激素协同作用促进细胞的生长和分化. 本研究也发现, 对于魔芋愈伤组织和丛生芽的分化, 内源细胞分裂素和生长素的总比值应维持在相对适宜的水平, 且不同组织各细胞分裂素的作用也有主次之分; 如6-BA, KT, 2,4-D质量分数太高会影响愈伤组织和丛生芽的分化, 导致组织的老化; NAA在一定浓度范围内, 可促进组织的生长分化和提高抗氧化的作用. 肖关丽等[28]发现, 植物组织培养过程中的各种变化与内源激素种类、 水平及配比具有重要关系, 包括对胚性细胞及体胚的形成, 以及愈伤组织分化芽和根等均具有显著影响. 适当提高内源激素NAA质量分数或提高6-BA/NAA, KT/NAA以及(6-BA+KT+2,4-D)/NAA的比值能显著促进愈伤组织及丛生芽的生长分化, 2,4-D与NAA的比值较大对促进丛生芽分化有抑制作用. 王秀红[29]研究指出, 内源激素的质量分数和配比是影响愈伤组织诱导率和绿苗分化率的关键因素之一, 表现为IAA和GA呈负效应, ABA,Z+ZR呈正效应; 同时, 内源激素的配比对愈伤组织诱导率和绿苗分化率也有一定作用, 表现为IAA/Z+ZR对愈伤诱导率有一定的正效应, Z+ZR/IAA,ABA/IAA对绿苗分化率呈显著正相关. 有研究证实, 魔芋细胞内CTK,IAA的代谢变化是细胞分化、 器官发生的调节因子和物质基础, 培养基中激素种类和浓度通过影响内源激素的代谢而起作用, 且影响魔芋细胞分化的内源CTK主要是iPAs类[30], 这与本研究结果基本一致. 另外, 魔芋细胞分化、 器官发生的生化反应顺序是: 激素代谢, RNA及DNA合成代谢, 蛋白质合成酶系统反应. 本研究也发现, 不同内源激素与抗氧化酶间均存在直接或间接相关作用, 同时与其他生理物质间也存在着相互作用, 从而共同影响愈伤组织和丛生芽的分化和外在形态.
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年9期)2022-07-18
中老年保健(2021年1期)2021-12-04
中国信息化周报(2020年10期)2020-04-07
江河文学(2020年6期)2020-01-04
东坡赤壁诗词(2018年3期)2018-07-16
家庭百事通·健康一点通(2018年3期)2018-03-30
农民致富之友(2017年23期)2018-01-02
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
银行家(2014年1期)2014-02-14
