
喀斯特不同土壤厚度小生境下土壤种子库特征
2022-09-19 12:19:54罗杰陈金艺郭旭曼雷颖刘锦春
西南大学学报(自然科学版) 2022年9期
罗杰,陈金艺,郭旭曼,雷颖,刘锦春
西南大学 生命科学学院/三峡库区生态环境教育部重点实验室/重庆市三峡库区植物生态与资源重点实验室, 重庆 400715
各种植物种子所采取的散落策略各异, 但无论是重力传播、 风播、 水传播, 还是依赖于各种动物传播, 成熟植物的种子最终都将回到土壤表面或进入土壤内部, 为植物群落的演替、 群落遭受干扰和破坏后的恢复补充新个体[1-2]. 可见, 土壤是陆生植物种子最主要的贮藏库. 而土壤种子库是指存在于土壤表层凋落物和土壤中全部活性种子的总和, 是植物繁殖体的储备库, 在种群天然更新、 群落演替、 植被恢复和生物多样性的保持等方面均发挥着关键作用[3-6]. 因此, 土壤种子库的研究对揭示地表植被的演替规律[7]、 探索过去植被的痕迹[8]、 预测未来植被的发展和探讨植被恢复的潜力[9]都具有十分重要的意义, 已成为现代生态学研究的热点之一[10-11].
我国西南喀斯特地区成土缓慢, 水土流失极快, 加上地形陡峭破碎、 剥蚀强烈、 洼地堆积等特点[12], 形成具有空间分布差异的不同土壤厚度小生境. 研究表明, 喀斯特地区土壤厚度不仅决定了喀斯特土壤生产力水平及其可持续能力[13], 更是喀斯特石漠化治理的重要影响因素[14]. 同时, 因植被的立地条件各异及生物间的竞争、 捕食关系等, 喀斯特不同土壤厚度小生境对种子的收集和储存及种子后续的萌发都有着不同的影响[15], 从而也将影响植被的更新和恢复进程. 目前, 大量研究关注了喀斯特生态系统中种子库与群落演替关系[16]、 土壤种子库与地上植被的关系[17]、 不同土地利用类型[18]和人为干扰方式下[19-20]土壤种子库特征. 然而, 对于异质性土壤生境下的种子库研究鲜有关注. 另外, 喀斯特地区生态系统退化, 植被多以草本、 灌木为主, 生存生长均受到一定限制[21]. 因此, 本研究通过对喀斯特地区灌草植物占优势的群落进行土壤种子库调查, 探究不同土层厚度小生境下土壤种子库储量、 分布特征和多样性, 以期为喀斯特地区植被恢复重建和可持续发展提供理论参考.
1 研究方法
1.1 研究区概况
研究区域位于重庆市沙坪坝区中梁山北段(29°39′-30°03′N, 106°18′-106°56′E), 为典型的喀斯特山地. 该区域属亚热带季风气候, 平均气温16.8~18 ℃, 气温较市区低4~5 ℃; 降雨充沛, 年降水量为1 000~1 300 mm; 海拔稍高于重庆的平均海拔, 为500~700 m. 本实验样地的主要植被是次生演替形成的灌木和草本植物, 土壤为黄色石灰土, 土层厚薄不均匀, 石缝众多.
1.2 土壤小生境调查
2019年11月18-25日, 在研究区域内随机选择3个植被类型一致的样地, 每个样地设置3个10 m×10 m的大样方, 在每个大样方的对角线(含四角)每隔1 m设置一个面积为50 cm×50 cm的小样方, 用竹签在4个角进行固定. 因此每个大样方共设置15个小样方(如果遇到石质基质不能取样, 则根据实际情况可向左右偏移), 共计135个小样方. 用钢钎在每个小样方内随机插入10次, 求平均值作为该样方的土层厚度, 通过对数据进行分析, 将调查的135个样方根据具体土层厚度划分为浅土层(0~10 cm), 浅中土层(0~20 cm)、 中深土层(0~30 cm)和深土层(0~40 cm)4个厚度等级.
1.3 土壤种子库取样
完成土层厚度的调查定位后进行土壤种子库取土, 采集时间选择在11月底, 也即多数植物种子成熟后的季节, 以小样方的实际土层厚度为标准, 在每个小样方的中心以12 cm×12 cm×10 cm(长×宽×高)的标准进行取土. 由于土壤环刀取土量不足, 正方形器具不好取样, 因此采用小土铲切出12 cm×12 cm的面积进行深度10 cm分层取样, 即浅土层(0~10 cm)的小样方共取土一层, 浅中土层(0~20 cm)的小样方共取土2层, 中深土层(0~30 cm)的小样方共取土3层, 深土层(0~40 cm)的小样方共取土4层. 每取一层土用尺子进行深度测量, 避免不同深度的土层混合. 将所取的小样方的土样分层装袋、 编号并带回实验室做萌发实验备用.
1.4 种子萌发实验
萌发实验在西南大学(29°49′N, 106°25′E)生态园实验基地内进行, 该基地海拔225 m, 属于亚热带季风气候. 将带回的供试土样自然风至微干, 去除残留在土壤中的植物根系和其他杂质等后, 将每份土样混合均匀. 混匀后的土样均摊于29 cm×29 cm×5 cm发芽盘中, 再将发芽盘置于温室大棚内(利于冬季保温保水, 保证不受外界种子的干扰)自然萌发. 萌发过程中适当补水, 保持土壤湿润. 种子开始萌发后, 每日观察和记录出苗的种类与数量, 标记已萌发的幼苗, 待其长大到能判别种属时, 记数(视其为有生命的种子)并将其从发芽盘上去除. 土壤搅拌后连续两周内无种子萌发时, 停止萌发实验. 对于只能等到成熟或开花时才能鉴定的植物, 将其移栽到其他容器中进行培养直至鉴定. 种子萌发实验于2019年12月21日-2020年7月28日进行.
1.5 数据分析
1.5.1 数据处理
土壤种子库密度: 土壤种子库密度用单位面积土壤内所含有的活力种子数量表示[22].
种子库中种子生活型: 参照《中国植被》(1980)中所采用的生活型系统, 将记录到的植物分为乔木(含小乔木)、 灌木(含小灌木)、 藤本(含草质和木质)、 多年生草本(含半灌木)植物和一年生草本植物5种类型, 分别计算每一类型植物占植物种总数的百分比[22].
种子库物种百分比: 计算种子库中每一种植物种子数所占所有物种种子总数的百分比[23].
1.5.2 衡量指标
种子库特征采用Margalef丰富度指数、 生态优势度指数(Simpson优势度指数)、 Shannon-Wiener多样性指数和Pielou均匀度指数[24]度量; 各生境不同土层之间种类组成上的相似性采用Sorensen相似性系数(Similarity Coefficient, SC)公式[25]计算. 各指数计算方法如下:
(1) Margalef丰富度指数(DMG):
式中:DMG为Margalef丰富度指数;S为群落物种数;N为所有类群的总个体数.
(2) Simpson优势度指数(D):
式中:D为 Simpson优势度指数;S为群落物种数;Pi为种i的个体数占群落中总个体数的比例, 即Pi=ni/N.
(3) Shannon-Wiener多样性指数(H):
式中:H′为 Shannon-Wiener指数;S为群落物种数;Pi为物种i个体数所占物种总个体数比例.
(4) Pielou均匀度指数(E):
E=H′/lnS
式中:E为Pielou均匀度值;H′为 Shannon-Wiener 多样性指数;S为群落物种数.
(5) Sorensen相似性系数(Similarity Coefficient, SC):
SC=2w/(a+b)
式中:w为土壤种子库和地上植被共有的植物种数;a和b分别为土壤种子库和地上植被的植物种数.
1.5.3 统计分析
采用SPSS软件中单因素方差分析(One-way ANOVA)比较不同土层厚度下土壤种子库植物萌发总密度, 用最小显著差异法(LSD)进行多重比较检验, 数据分析在SPSS 13.0和Excel中完成, 并用Origin 2018作图.
2 结果与分析
2.1 不同土壤厚度小生境下土壤种子库的储量与空间分布
从浅土到深土生境, 即随着小生境土壤厚度的增加, 土壤种子库总密度处于逐渐递增的趋势并在深土生境中达到最大值. 同一土壤生境下, 随着土壤层级的增加, 土壤种子密度呈显著递减趋势(图1).
对不同小生境的相同土层进行水平对比发现, 0~10 cm土层下浅土生境土壤种子密度最高, 浅中土生境的土壤种子密度最低, 二者差异有统计学意义, 其他小生境在这层密度差异不明显; 10~20 cm土层下深土生境中土壤种子密度达到最高, 中深土生境密度最低; 20~30 cm土层下同样为深土生境中的土壤种子密度大于中深土生境(图1).
2.2 不同土壤厚度小生境下土壤种子库物种及生活型组成
在本研究4种喀斯特小生境土壤种子库中, 共统计到69种植物种子, 隶属于3纲22目30科62属, 其中以菊科植物(17种)最多, 其次为禾本科植物(7种), 再次为蔷薇科植物(4种), 唇形科、 大戟科、 蓼科、 桑科、 石竹科均为3种, 其余科为1~2种. 其中, 在浅土生境土壤种子库中共统计到44种植物种子, 隶属于3纲19目24科41属, 优势种以风轮菜(Clinopodiumchinense)、 酢浆草(Oxaliscorniculata)、 八宝(Hylotelephiumerythrostictum)、 繁缕(Stellariamedia)、 鼠曲草(Pseudognaphaliumaffine)为主; 浅中土生境土壤种子库中统计到49种植物种子, 隶属于3纲18目25科45属, 优势种以酢浆草、 风轮菜、 黄鹌菜(Youngiajaponica)、 鼠曲草、 马唐(Digitariasanguinalis)为主; 中深土生境土壤种子库中统计到54种植物种子, 隶属于3纲18目22科52属, 优势种以酢浆草、 风轮菜、 鼠曲草、 黄鹌菜、 马唐为主; 深土生境土壤种子库中统计到48种植物种子, 隶属于2纲18目21科45属, 优势种以风轮菜、 鼠曲草、 酢浆草、 马唐、 黄鹌菜为主.
进一步对不同土壤厚度小生境的同一土层厚度进行种子库种类探究, 发现在各土壤厚度下种子库均以菊科植物和禾本科植物占优势. 在0~10 cm土壤厚度下, 中深土生境中土壤种子库物种种类为50种, 在4种小生境下最高, 而深土生境中土壤种子库仅有44种植物种类, 为最低; 10~20 cm土壤厚度下的趋势与0~10 cm土层一样, 在中深土生境土壤种子库中植物种类数最高而在深土生境土壤种子库中最低; 在20~30 cm土壤厚度下, 中深土生境土壤种子库中植物种类为32种, 而深土生境土壤种子库中植物种类有31种. 从总的土层来看, 各小生境土壤种子库物种数随着土层的加深而减少(图2).

S10表示浅土生境, S20表示浅中土生境, S30表示中深土生境, S40表示深土生境; D0~10表示0~10 cm土层, D10~20表示10~20 cm土层, D20~30表示20~30 cm土层, D30~40表示30~40 cm土层, 下同. 不同字母代表不同土壤厚度小生境下同一土层间差异有统计学意义, p<0.05.图1 不同土壤厚度小生境下不同土层中土壤种子库密度(平均值±标准误)
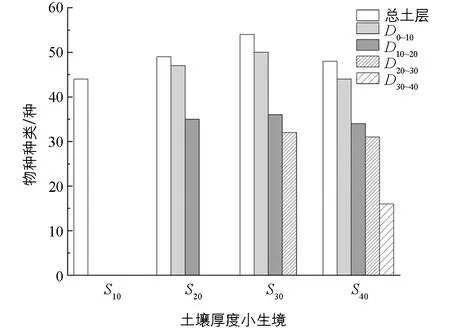
图2 不同土壤厚度小生境下土壤种子库的植物种类
4种小生境土壤种子库中均以草本植物为主, 乔木、 灌木和藤本植物3类生活型所占的比例较小, 在每一生境中多年生草本分别占比47.7%(浅土生境), 46.9%(浅中土生境), 44.4%(中深土生境), 41.7%(深土生境), 一年生草本分别占比40.9%(浅土生境), 30.6%(浅中土生境), 38.9%(中深土生境), 39.6%(深土生境), 深土生境土壤种子库较其他小生境的种子库更丰富, 除了多年生草本、 一年生草本、 一年生或多年生草本、 灌木、 藤本植物外, 还发现了乔木植物的种子(图3).
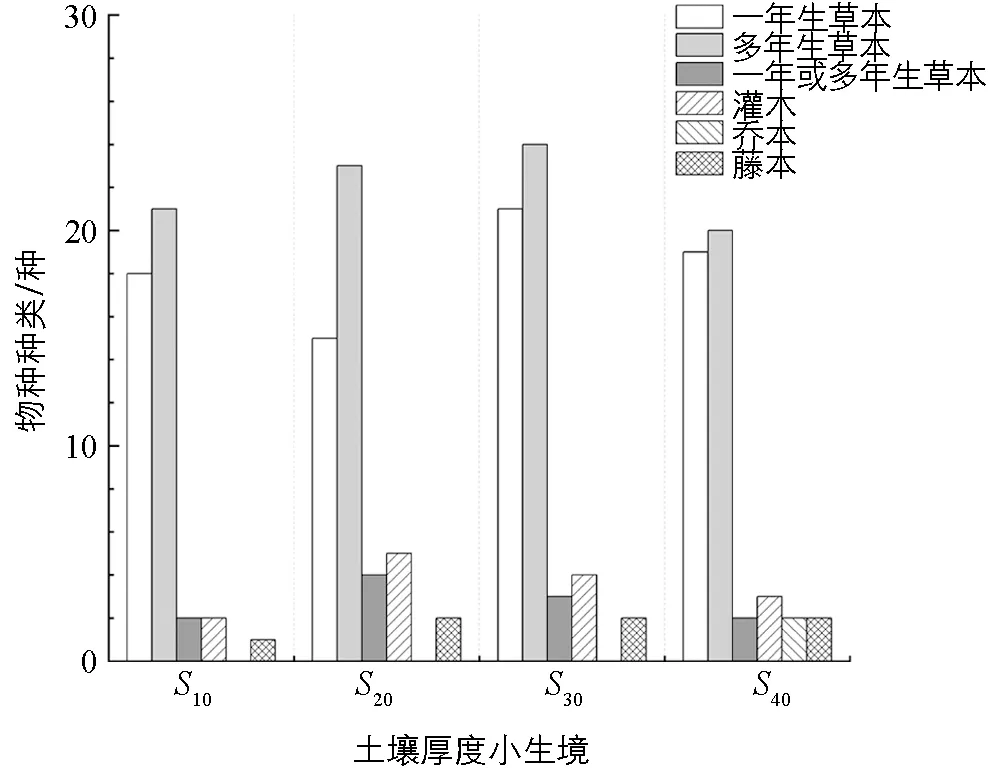
图3 不同生活型植物种子物种数在四种土壤厚度小生境中的分布
对各小生境土壤种子库中植物生活型在不同土层下的种类数进行探究, 我们发现除乔木外, 在4种小生境下各生活型的种类均随着生境土壤厚度的加深而递减. 各小生境均在0~10 cm土层下具有最大的一年生草本、 多年生草本、 一年或多年生草本、 灌木、 藤本物种数量. 在0~10 cm土层下, 4种小生境中土壤种子库均以草本植物为主, 并在中深土生境时各植物生活型种类数量取得最大值; 在10~20 cm土层下, 浅中土、 中深土、 深土生境中土壤种子库均以多年生草本和一年生草本植物为主, 但在中深土生境下占比最高, 达88.9%(浅中土生境为82.9%, 深土生境为85.3%); 在20~30 cm土层下, 深土生境(87.1%)下草本植物中的占比较中深土生境(90.6%)更低(图4).

图4 土壤中不同生活型植物种子在不同土壤厚度小生境中分层分布
2.3 不同土壤厚度小生境下土壤种子库多样性分析
各小生境下土壤种子库在中深土生境的Margalef物种丰富度指数最高, 其次为浅中土、 深土、 浅土生境; 而Shannon-Wiener多样性指数为浅中土生境最高, 浅土生境最低; 物种均匀度指数在浅中土生境中最高, 在浅土生境与中深土生境相同, 但总体上4个小生境均匀度指数相差不大(均匀度指数越高, 植物的数量在此区域内的一致程度较其他层次越高); 生态优势度则呈现出相反的趋势, 在浅土生境中达到最高, 物种数量分布呈现不均匀的现象更明显(表1).

表1 不同土壤厚度小生境总物种多样性
对不同土壤小生境同一土层的土壤种子库物种多样性指数进行分析, 0~10 cm土层下中深土生境中Margalef物种丰富度指数最高, 其次为浅中土、 浅土, 深土生境最低; Shannon-Wiener多样性指数为浅中土生境最高, 其次为深土、 中深土、 浅土生境; 均匀度指数在浅中土和深土生境下最高, 浅土生境中最低; 生态优势度在浅土生境最高; 在10~20 cm土层下, 物种丰富度指数从高到低依次为中深土、 浅中土、 深土生境; 多样性指数为浅中土生境最高, 深土生境最低; 均匀度指数从高到低依次为浅中土、 中深土、 深土生境; 生态优势度则在深土生境中最高; 在20~30 cm土层下, 中深土生境物种丰富度和生态优势度指数均高于深土生境, 但多样性指数和均匀度指数相比较低(表2).
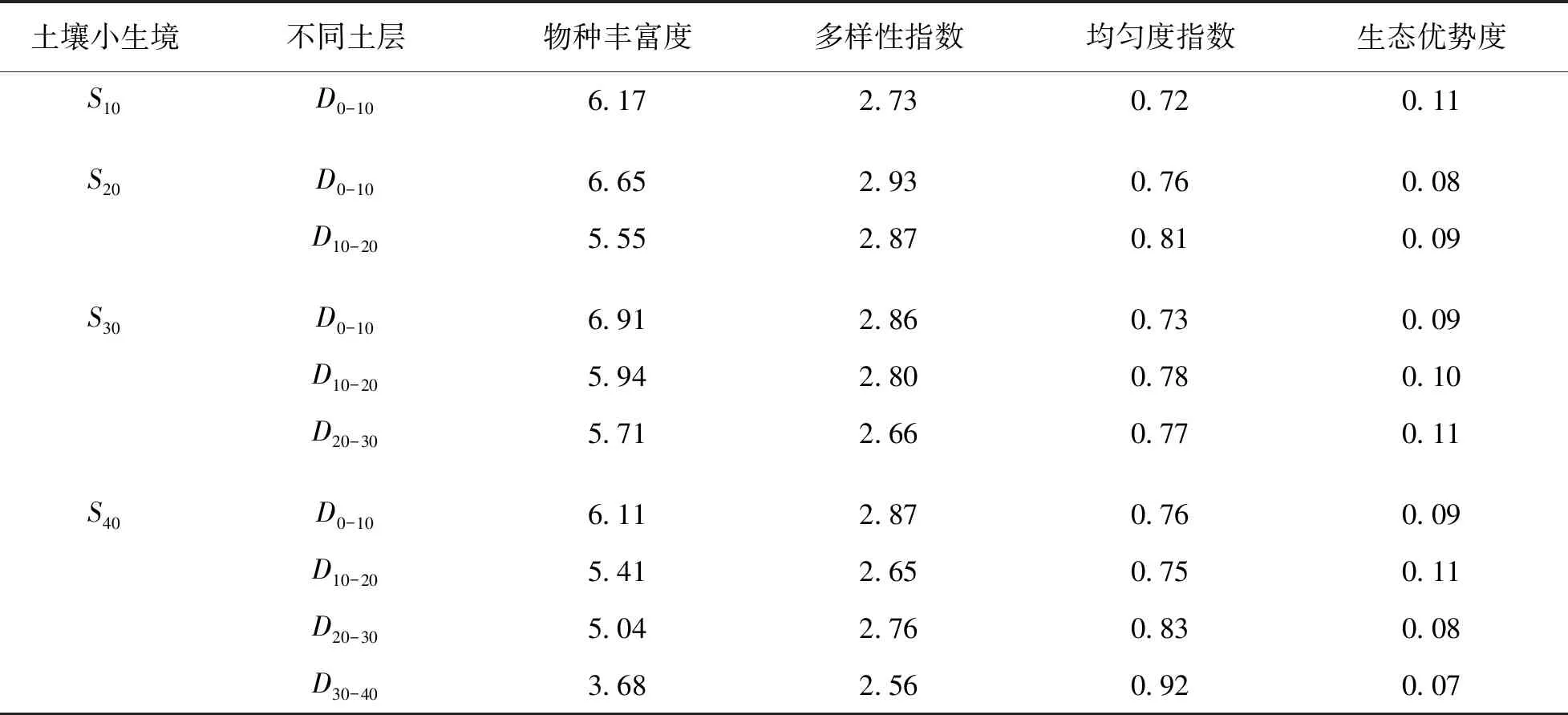
表2 不同土壤厚度小生境下不同层土壤种子库的物种多样性
2.4 土壤种子库的相似性特征
从表3中可以看出, 各小生境不同土层之间种子库组成成分相似性系数变化范围为0.320~0.852, 大部分为中等相似性(相似性值位于0.50至0.75之间), 体现出较大的差异性, 在同一土壤生境下各土层间与不同土壤生境下相同土层间土壤种子库的相似性均较高, 但与不同土壤生境下的不同土层土壤种子库之间大部分呈现出较低的相似性(表3).
3 讨论
3.1 不同土壤厚度小生境下土壤种子库总密度、 物种组成与多样性分析
从不同土壤厚度小生境来看, 土壤种子库中植物总的物种组成和密度均在深土生境中最高, 与朱波等[26]的研究相同. 可能原因是浅土生境中植物本身比较少, 因此产生的种子数量也较少, 深土生境中植物种类增多, 产生的种子数量也较多. 另外也说明更深的土壤能储存更多的种子, 因为深土小生境相较于浅土小生境有更大的种子储存空间. 对于物种组成, 我们发现在不同小生境下土壤种子库的物种组成较为一致, 按科来分, 物种数量从多到少依次为: 菊科、 禾本科、 蔷薇科、 唇形科、 大戟科, 按生活型来分, 物种数量从多到少依次为: 多年生草本、 一年生草本、 灌乔木, 说明研究区域目前处于演替的早期阶段, 物种主要为菊科和禾本科等先锋草本植物.
土壤种子库的物种丰富度、 多样性和均匀度指数在中土生境中取得最大值. 可能原因是: 浅土生境中由于植物本身数量较少, 因此产生的种子数量也较少; 随着土壤深度的增加, 土壤的种群承载力越来越强, 开始能够承载较大个体的植物, 在中等深度的土壤中较大个体与小个体植物之间能够共存, 使中等深度的土壤小生境上的植物种数更加丰富, 种子数量较浅土生境有所增加; 在深土生境中, 个体较大的植物在竞争中胜出, 在群落中占优势地位并排挤掉其他小型植物种, 相应地留在土壤中的草本、 灌木植物种类较中等土厚生境种类减少, 较大个体的种子种类少量增加(比如乔木种子出现), 表现出物种丰富度、 多样性略低于中等土深生境. 除此之外, 生态优势度值在浅土生境中略高于其他小生境. 生态优势度值越高表明群落中优势种越单一或稀少, 一旦遭受破坏则恢复能力较差, 一般在演替早期, 水热条件较差或干扰强度大的群落生态优势度值较高[27]. 说明本研究中的浅土生境多是先锋种定居于此, 其种子库表现出的恢复潜力不大. 但是, 生态优势度的变化趋势从另一方面也反映出随着土壤厚度的加深, 土壤种子库中种类和生活型组分趋向多样化, 生境更加优越, 被破坏后恢复能力也更强.
3.2 不同土壤厚度小生境各土层中土壤种子库密度、 物种组成与多样性分析
土壤在不同的深度具有不同的理化性质和营养条件, 表层土壤的结构疏松, 通气性强, 微生物活动强烈, 土壤养分含量较高, 而深层土壤更接近于母质, 土壤的养分含量急剧降低[28]. 有研究发现, 喀斯特地区土壤种子库中的种子多集中分布于土壤表层, 随着土厚的加深, 土壤中植物种子的种类和数量逐渐减少[18,20,29]. 与上述研究结果相似, 本研究在4种不同土壤厚度小生境下, 植物的种子数量和种类随着土层深度的增加均呈现出垂直递减的趋势. 虽然落到地表的种子通常通过雨水冲刷、 动物活动或土壤裂隙的存在等不同的途径向下移动, 但是大多数植物的大部分种子难以主动埋入较深的土层中, 所以种子数量分布多集中在土壤表层(0~10 cm), 土壤种子库的物种多样性和丰富度指数也随着土层加深而降低.
种子库除了在同一生境的不同土层变化, 我们也发现在不同生境的相同土层上, 其物种组成和生活史具有相似性, 即各小生境土壤种子库的物种组成在相同土层下均以菊科植物和禾本科植物占优势; 除深土生境20~30 cm土层外, 各土壤小生境种子库在相同土层下, 物种组成按生活型划分, 物种数量从多到少依次为: 多年生草本、 一年生草本、 灌乔木, 这与本研究中的不同小生境的土壤种子库的物种组成和生活型相同. 段启辉等[30]在相同研究区对于种子大小分布的研究结果表明, 该地区大部分种子为小种子. 这些先锋植物往往具有较强的生态适应能力, 例如风传播是菊科常见的传播方式[31]. 另外, 菊科和禾本科的种子多而小[32], 易在雨水的冲刷和动物的活动下, 通过土壤裂隙和掩埋进入土中, 使同一生境下不同土层的土壤种子库出现与该生境基本相同的物种组成和生活型组成.
不同土壤小生境不同土层之间种子库体现出较大的差异性, 即在同一土壤生境下的各土层间与不同生境下的相同土层之间土壤种子库的相似性均较高, 但与不同生境下的其他层次大部分呈现出较低的相似性. 一般来说, 同一生境下的植物类型在同一垂直剖面上的组成和结构较为相似, 因此土壤种子库间具有较高的相似性. 虽然不同土壤小生境相同土层的土壤深度基本一致, 但是小生境总体土壤深度不同, 承载的植物群落不同, 更深的土壤能保存更多的水分[33], 能够支撑植物群落进一步发育, 使不同小生境物种组成间的差异性增强, 群落的生态距离逐渐增大. 所以不同土壤小生境的相同土层的土壤种子库的相似性低于同一土壤生境下不同土层土壤种子库的相似性, 且各土壤小生境同一土层相似性系数各异.
本研究表明, 喀斯特不同土壤厚度小生境会影响土壤种子库的密度及其分布, 深土小生境中土壤种子库的总密度最大, 同一土壤生境不同土层的种子库密度呈现出降低的趋势. 土壤种子库的物种组成和多样性也会随着土壤深度变化而呈现出一定的规律性, 浅土和深土生境可能由于资源限制和种间竞争等原因限制, 使中深土小生境中土壤种子库物种组成最丰富, 多样性最高. 不过中深土小生境维持物种多样性的原因还需要进一步探明. 另外, 本研究中所有土壤异质性小生境中的土壤种子库组成均具有较高的相似性, 并且大部分以草本为主, 表现出的恢复潜力较小, 植被的恢复还需要适度进行人为干预.
猜你喜欢
科普童话·学霸日记(2022年1期)2022-05-30 10:48:04
科普童话·学霸日记(2022年1期)2022-05-30 10:48:04
——以贵阳花溪公园为例
山地农业生物学报(2022年3期)2022-05-13 09:58:54
敦煌学辑刊(2017年4期)2017-06-27 08:07:18
物流技术与应用(2017年3期)2017-05-17 05:29:04
现代防御技术(2016年1期)2016-06-01 12:13:27
启蒙(3-7岁)(2016年10期)2016-02-28 12:27:10
启蒙(3-7岁)(2016年9期)2016-02-28 12:26:59
计算机工程(2015年8期)2015-07-03 12:19:20
计算机工程(2014年6期)2014-02-28 01:26:31
