
谷物发酵产品的营养功能提升与益生功能研究进展
2022-09-17 12:58:00杨庆华张亚飞田晓静王稳航
食品与发酵工业 2022年17期
杨庆华,张亚飞,田晓静,王稳航
(天津科技大学 食品科学与工程学院,天津,300457)
谷物属于禾本科植物,其果实称为颖果,所有的谷物颖果均具有相同的基本结构(麸皮、胚乳和胚芽)及相似的营养特征[1](以小麦为例,其结构及营养分布见图1)。谷物中含有丰富的碳水化合物、蛋白质、矿物质、膳食纤维和维生素等营养成分,是维持人体活动的重要能量来源[2],也是最具潜力的食品加工原料之一。
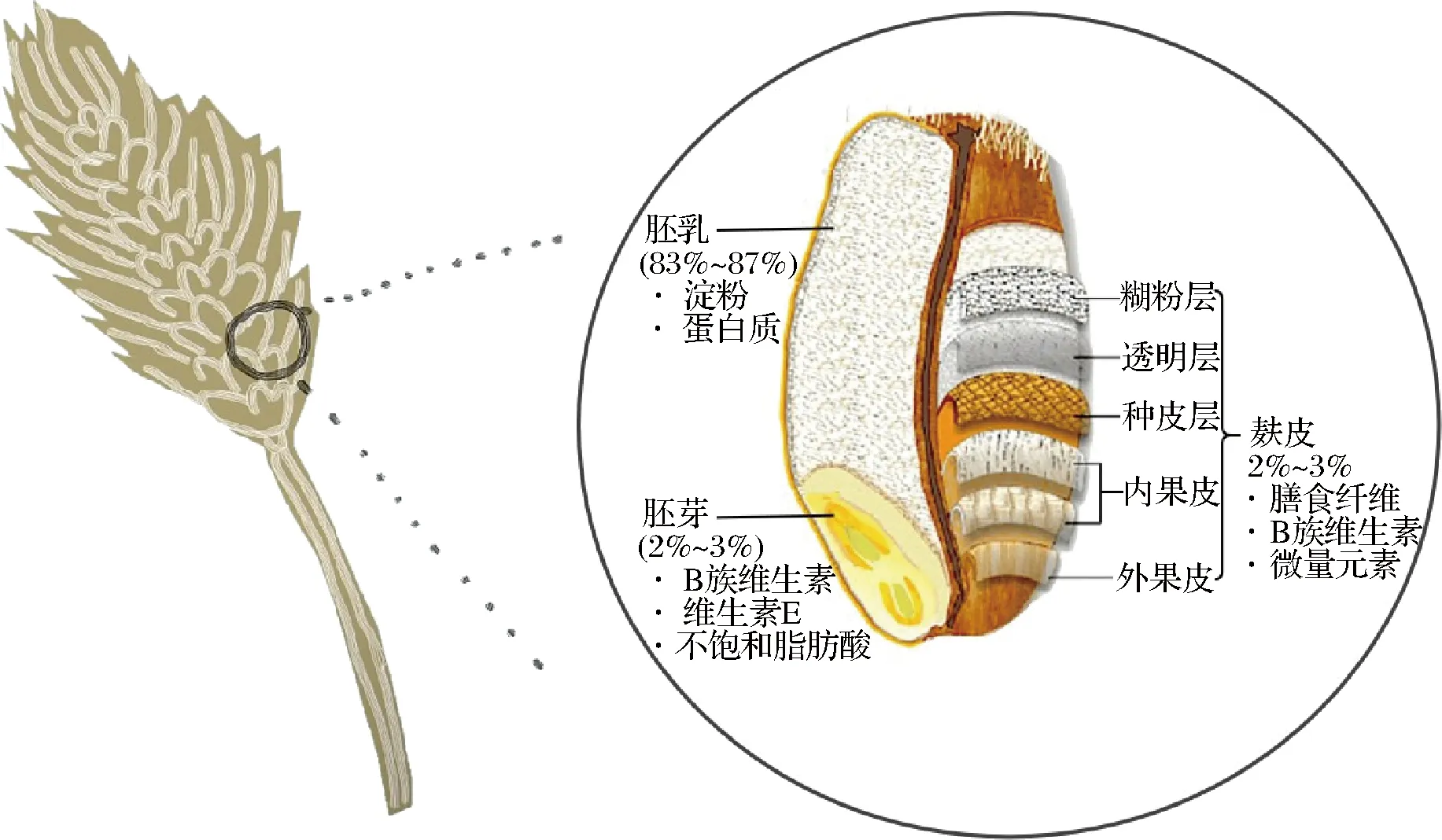
图1 小麦的结构特征及营养分布Fig.1 Structural characteristics and nutrient distribution of wheat
但由于抗营养素(如植酸、单宁和多酚)的存在、蛋白质人体消化率偏低、赖氨酸缺乏及粗糙的质地结构,谷物食品可能会存在营养利用受限、不易消化、味道寡淡、可食性差等问题。近年来,为改善谷物的营养及风味缺陷,发酵成为谷物加工的首选方式。发酵主要通过酸的激活、酶的催化及微生物的代谢产物对谷物基质产生影响[3],提高其蛋白质消化水平,生成酚类、维生素等功能活性物质,增强谷物营养,同时提升其风味和适口性。相较于营养损失严重的研磨、浸泡、蒸煮等加工方法,谷物发酵能最大限度地满足人类对健康、营养、低脂或纯素等饮食模式的需求[4],成为功能性食品时代的新选择。
近年来,发酵对谷物的积极作用受到国内外学者的广泛关注,本文综述了国内外传统和新型谷物发酵产品,比较分析了发酵对谷物消化率和生物活性的影响,以及作为益生菌载体的作用和益生元潜力。旨在为谷物发酵类产品提供更为全面的营养评价,并为新型谷物产品的开发提供参考和启示。
1 谷物发酵
1.1 谷物发酵剂
发酵主要通过微生物的活动对底物进行有益的转化或修饰以得到所需的产品,谷物食品中涉及的微生物主要为乳酸菌、酵母菌及真菌(霉菌)。乳酸菌具有降解谷物中淀粉、葡萄糖等糖类物质的能力,从而产生大量乳酸、乙酸等物质。大多数谷物都依赖乳酸菌来介导发酵过程[4],尤其是异型发酵(除乳酸外,还包括乙酸、乙醇、CO2等产物)的乳酸菌菌株(除片球菌属、乳球菌属、链球菌属外的大多数乳酸菌[5])。该类型的发酵通常会产生更多的理想风味化合物,如乙醛和双乙酰,赋予产品独特的甜酸味和香气,因此可以改善谷物感官活性化合物含量低,味道寡淡等不良特性[6]。酵母菌是负责生产谷物酒精饮料和面包的主要微生物,其代谢产生酒精和二氧化碳等产物,能够改善产品质地,赋予产品特有的风味特征。酿酒酵母是用于谷物发酵的主要酵母种类,其他种类的酵母也参与了谷物产品的生成,如假丝酵母、克鲁维酵母等。曲霉、根霉等霉菌有时也与乳酸菌或酵母菌共同参与谷物的发酵过程,不同种类的霉菌对谷物蛋白进行碱性发酵,释放出氨,并为后续的酵母或乳酸发酵产生风味前体物质,增加产品的香气。
1.2 谷物发酵产品
受到气候、社会经济条件、烹饪方式、可用基质、自然菌群等多重因素的影响,不同地区的谷物基发酵产品形态各异,种类繁多。表1为世界各地(多集中在非洲和亚洲)常见的谷物发酵产品,主要包括发酵食品和无酒精饮料。
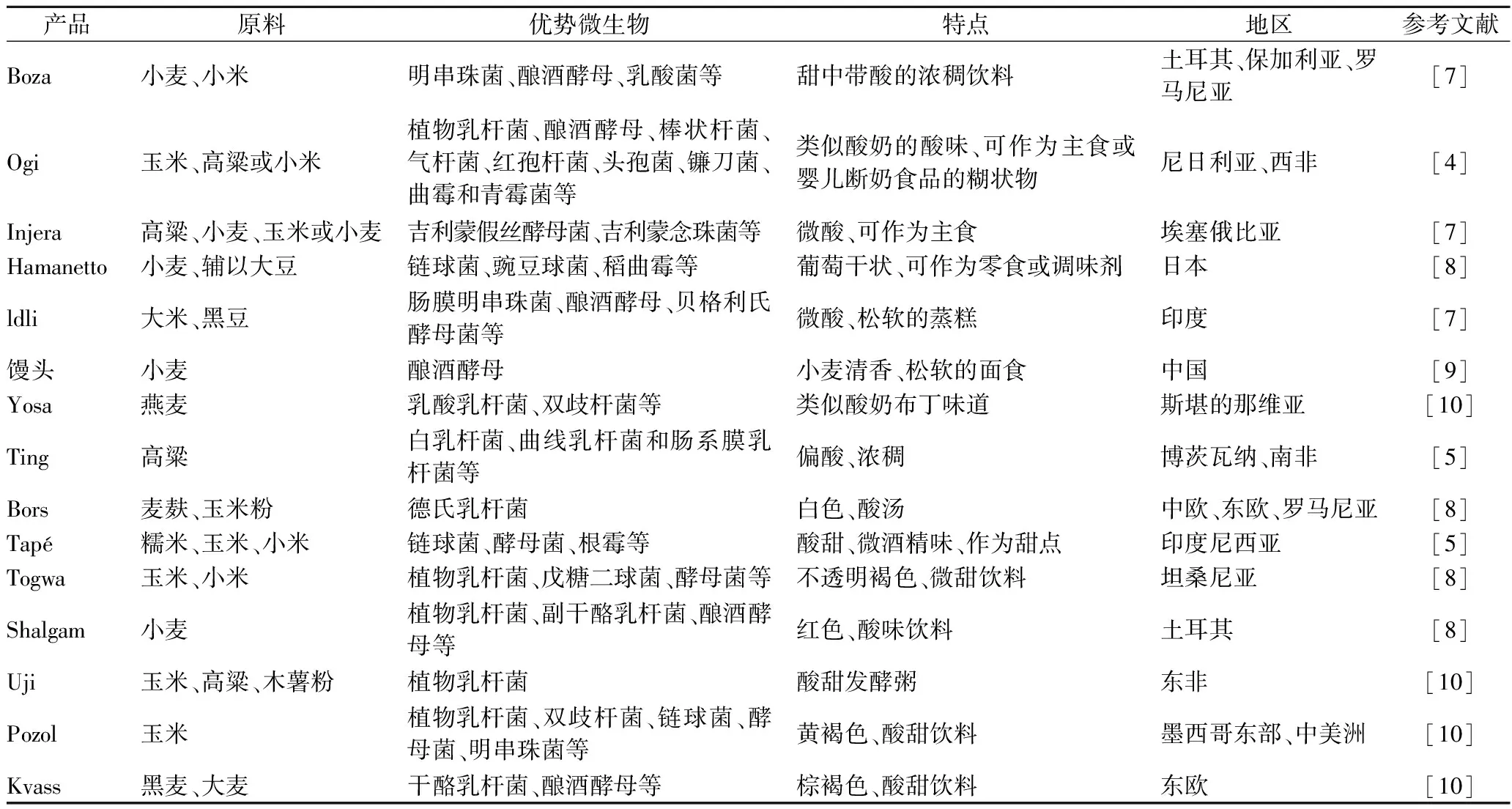
表1 世界各地最常见的谷物发酵产品Table 1 The most common cereal fermentation products in the world
2 谷物发酵食品的营养改良
近年来,利用发酵技术手段在改善谷物的营养缺陷方面取得巨大进展,尤其是在营养价值和功能活性的提升方面(图2)。

图2 发酵对谷物营养价值与生理功能的影响Fig.2 Effect of fermentation on nutritional value and physiological function of grain
2.1 常规营养成分
2.1.1 碳水化合物
碳水化合物是血糖的主要来源,饮食中快速消化的碳水化合物往往会导致血糖水平的迅速上升,继而引发糖尿病等代谢综合征。淀粉是人类食物中最大的碳水化合物来源,因此谷物淀粉的消化对人体健康,尤其是血糖平衡至关重要。
淀粉消化与食物的血糖指数(glycemic index,GI)直接相关,利用乳酸菌发酵小麦和黑麦基质已被证明可降低产品的GI值及胰岛素指数[11]。发酵可降低淀粉消化率的几种可能机制也已被提出,在发酵过程中,有机酸特别是乳酸的形成被认为是主要原因[12],酸作用的生理机制似乎各不相同,乳酸可降低谷物基质pH,减慢淀粉的消化速度,而醋酸和丙酸则延长了胃的排空时间[12]。此外,淀粉凝胶化程度越大,水解速度越快。发酵过程中复杂的化学反应被认为可以降低淀粉的凝胶化程度,这也在一定程度上解释了酸面包消化率较低的原因[13]。发酵也被证明能促进抗性淀粉的形成,而抗性淀粉具有极低的消化速率[14]。在最近的研究中,DIOWKSZ等[15]发现,血糖反应的进行与发酵对α-淀粉酶的抑制有关,与对照面包相比,发酵面包中的α-淀粉酶在体外受到抑制,淀粉降解减少,进而导致可吸收的葡萄糖变少,表现出良好的抗糖尿病潜力。GIL-CARDOSO等[13]对大鼠喂食不同种类的酸面包,结果均显示出较低餐后血糖水平。
除淀粉外,谷物产品中还存在一类吸收较差的短链碳水化合物FODMAPs,FODMAPs是可发酵(fermentable)的单糖(monosaccharides)、寡糖(oligosacchrides)、双糖(disaccharides)和多元醇(polyols)组成的缩写[16]。不同于淀粉,这类具有渗透活性的分子在小肠中吸收较差,到达大肠后会迅速被大肠细菌发酵,从而造成部分人群的腹胀、腹痛、便秘,引发肠易激综合征。研究发现,发酵可以改变谷物中的FODMAPs浓度,乳酸菌、酵母和真菌可以部分甚至完全降解FODMAPs。在面团发酵过程中,除多元醇类外,其他所有的FODMAPs均显著减少,在烘烤加工结束时,蔗糖、果糖和葡萄糖就已被完全降解[17],这也意味着仅自然发酵便可减少30%以上的FODMAPs,从而使产品中可发酵碳水化合物和游离葡萄糖含量降低。同样在STRUYF等[18]的研究中,在克鲁维酵母菌株的作用下,产品中果聚糖水平降低了90%以上,得到较低FODMAPs水平的面包产品,这对肠易激综合征患者来说是十分有益的。MENEZES等[19]也强调乳酸菌和酵母菌混合培养物是降解FODMAPs最有效的工具之一,这两种菌酶的结合,为生产符合特殊饮食需求的高附加值面包提供了可能。
2.1.2 蛋白质
谷物蛋白质资源丰富,脂肪含量低,具有较高的营养价值和功能特性。但谷物蛋白在人体消化率较低,往往低于动物蛋白,这主要受到自身蛋白质结构、抗营养因子(蛋白酶抑制剂、植酸、单宁)的影响。谷物蛋白中脯氨酸的含量较高,其侧链与肽键形成吡咯环限制了肽键的自由旋转,不但使谷蛋白侧链显示出了很强的疏水性,而且也使蛋白质链灵活性降低,对水解酶产生高抗性,蛋白难以消化[20]。酚类可以通过多点氢键、疏水结合等与蛋白质形成复合物,导致蛋白的聚沉[21]。植酸也可通过竞争肽酶活性所需的矿物质辅助因子并与蛋白质直接作用,干扰蛋白质的消化吸收[22]。此外谷物中的膳食纤维也与蛋白质水解受阻有关,但目前膳食纤维的作用被认为是纯粹的物理作用,即通过增加胃肠道内容物的黏度,影响水解酶的扩散,使其难以接触到底物,减少蛋白消化水解[23]。
目前,发酵技术成为提高谷物蛋白消化率,改善谷物蛋白品质的有效手段之一。发酵可通过抑制消化酶的非营养性化合物水平、降低抗营养因子交联及产生微生物蛋白酶改变谷物蛋白质结构及水解参数[24],促进谷物蛋白的降解。在不同谷物的酸面团探究实验中,与普通面包相比,发酵酸面包的消化率提高了16%,总游离氨基酸浓度增加,蛋白质的生物学价值提高了18.7%[25]。OGODO等[26]也证实发酵后的高粱面粉蛋白消化率升高,并且推测水解酶可能已将高度不溶性的贮藏蛋白质转化为更简单和可溶的产物。此外,发酵过程中pH的下降也被认为有利于谷物中内源酶的激活,以及谷蛋白的增溶和解聚[26]。
在发酵过程中,蛋白质的降解更利于活性多肽的产生,如短支链氨基酸和小型多肽,这些化合物已被证明有助于调节胰岛素反应,并为Ⅱ型糖尿病和心血管疾病提供保护[27]。CODA等[25]发现酸面团中的水/盐溶性提取物的自由基清除活性要显著高于化学酸化面团,并且鉴定出的25个多肽中,几乎所有序列都具有抗氧化肽的典型组成特征,所有的纯化组分对小鼠成纤维细胞均显示出体外抗氧化活性。
此外,谷物蛋白也是导致食物过敏最常见的原因之一,小麦蛋白可诱导皮肤、肠道或呼吸道的典型过敏反应[28],造成腹泻、腹痛、红肿等症状,这类人群往往需遵循无谷饮食,而发酵蛋白改性则对降低其致敏性提供了可能。DE ANGELIS等[29]证明了益生菌水解小麦过敏原的能力。他们以该益生菌株为发酵剂发酵小麦,发现小麦蛋白质可被显著降解。此外,大麦和黑麦等的醇溶蛋白易引起小肠黏膜损伤,引发乳糜泻。而发酵过程中醇溶蛋白的大量降解已被证明可以达到乳糜泻患者的耐受程度。同时,一项为期两个月的临床试验表明,发酵小麦生产的饼干和蛋糕对乳糜泻患者没有毒性[30]。
2.1.3 膳食纤维
膳食纤维(dietary fibers,DFs)是一种植物碳水化合物,具有容纳大量水分并增加粪便膨胀,影响胃肠道运转和营养吸收率,改善糖尿病、心血管、胃肠道疾病等多种功能益处。谷物是膳食纤维的良好来源,其DFs主要由非淀粉多糖(non-starch polysaccharides,NSP)、抗性淀粉、低聚糖(主要是果聚糖)和非碳水化合物多酚醚木质素构成,NSP主要包括阿拉伯木聚糖(arabinoxylans,AX)、β-葡聚糖和纤维素,这类聚合物通常与木质素、木酚素、酚酸和少量蛋白质以复合结构共存在于细胞壁中[3],使其难以加工和利用。
发酵是谷物膳食纤维改性和释放的重要手段,由于有机酸(主要是乳酸和乙酸)的产生,可能导致谷物中多种内源性酶或细菌酶被激活,生物聚合物发生降解,质地软化(细胞壁降解),促进纤维的溶解或释放[31]。近年来,探讨发酵对β-葡聚糖和AX的研究尤为广泛。β-葡聚糖是大麦、燕麦中常见的膳食纤维,属于高分子质量聚合物,通常会在胃肠道中增加黏度,从而影响胃排空率、肠道运输时间和营养吸收率。XIAO等[32]发现,通过发酵可以改变β-葡聚糖结构状态,降低β-葡聚糖的分子质量,并且增大了β-(1→3)与β-(1→4)残基的比例,这些结构的改变使得大麦β-葡聚糖的水吸附或分子结合能力增强,生理活性得到强化,增加了对血糖、脂肪的健康调控。谷物产品中的AX因具有增加肠道内食糜的黏度,降低胆固醇和血糖的潜能而被人们关注。发酵可以增大谷物中AX的溶解性,ZHAO等[33]研究了不同发酵程序对麦麸的影响,与未经处理的麦麸相比,所有发酵样品中的AX含量提高了3~4倍。面粉中AX的含量对面团的流变特性和最终产品的质量也具有重要影响,并且由于AX的增溶作用,酸面团发酵可能会改变其发酵方式,并产生益生元低聚糖[34]。
人们对富含膳食纤维的酸面包进行了研究。MIHHALEVSKI等[35]评估了黑麦酵母面包中的膳食纤维,发现在加工过程中,可溶性和不可溶性膳食纤维的比例有所增加。他们将这种差异归因于面包制作过程中的生化和微生物过程。抗性淀粉的形成引起了总膳食纤维含量的增加,而可溶性纤维含量的增加则被认为是不溶性纤维转化的结果。BOSKOV HANSEN等[36]也提出了黑麦面粉中固有酶的活性可能导致了膳食纤维组分的再分配。此外,富含膳食纤维的面包往往存在体积小,硬度大等不良特征,麸皮预发酵被认为是提高高纤维面包质量的潜在途径,使用发酵的麸皮可以影响CO2在面包中的滞留,从而增大面包的体积和面包屑的柔软度[35]。已有报道,含有10%发酵麸皮的面包表现出最佳的感官特性[37]。
2.2 微量营养成分
2.2.1 维生素
维生素是维持人体代谢的重要营养素,但因在人体内合成受限,往往通过食物从外界获取。谷物是优质维生素的天然来源,尤其是核黄素、钴胺素、叶酸等B族维生素和E族维生素含量丰富,对维持人体维生素的平衡具有积极作用。
大多数维生素稳定性差,易受到加工方式的破坏。因此,许多国家实施了补充或强化维生素计划[38],但相较于化学合成的维生素,微生物发酵提供了一种更为天然、安全和经济的强化方法,尤其是对谷物中B族维生素的富集。
叶酸(维生素B9)作为单碳转移的辅酶,参与核苷酸、氨基酸和其他关键细胞成分的合成、相互转换和修饰,对人体代谢至关重要[39]。目前叶酸缺乏症依然是一个全球性的健康问题[38],许多研究表明乳酸菌和酵母菌均可以合成叶酸,但目前在对谷物基发酵食品的研究主要集中于酵母菌对叶酸含量的影响[40]。KARILUOTO等[41]借助SaccharomycescerevisiaeALKO743、CandidamilleriABM4949、Pseudomonassp.ON8等特定菌株对燕麦和大麦进行发酵,结果表明叶酸水平得到大幅度提升。HJORTMO等[42]利用光滑假丝酵母发酵玉米粥,最终叶酸含量是未发酵玉米粥的23倍。虽然有研究证明乳酸菌也可以合成叶酸,例如,黑麦和燕麦酸面包中的叶酸水平在乳酸菌的作用下得以提升[41],但是不同乳酸菌株之间差异性较大[43],甚至许多乳酸杆菌会消耗叶酸。另外,也并非所有产生叶酸的细菌都适用于生物强化,某些乳酸乳杆菌、双歧杆菌和明串珠菌合成的叶酸并不能排出细胞[44],此时叶酸的生物利用度极低,所以对食品营养的改善很小。因此,叶酸生产菌的合理筛选是进行叶酸合成和强化的前提。
维生素B12是一种植物性食物中较为短缺的维生素,因与细胞性贫血、外周动脉疾病和各种神经系统疾病密切相关而受到人们的重视,最近的研究发现,通过麦芽粉、大麦粉和小麦糊粉发酵,可使丙酸杆菌产生充足的活性维生素B12[45]。XIE等[46]利用丙酸杆菌和短乳杆菌的联合发酵,使其维生素B12水平达到干重的(357±8) ng/g。此外,发酵也可以促进其他多种维生素的合成,例如,在乳酸菌、酵母菌等介导下,硫胺素、核黄素等含量得到明显增加[47]。但值得注意的是,目前关于谷物发酵对维生素E浓度影响的研究较少,目前只有VERNI等[48]利用冬虫夏草发酵的陈米,维生素E含量提高了100%。另外利用发酵小麦胚芽和麸皮组成的营养补充剂Lisosan G®喂食家兔,其血液中维生素E含量得到增加[49]。
2.2.2 酚类化合物
酚类化合物由于特殊的化学结构,可促进芳环上的羟基提供氢或电子,并赋予自由基清除活性和金属螯合电位,因此具有高抗氧化、抗癌、抗炎等多种有益特性。谷物中常见的酚类物质有酚酸、类黄酮和单宁。酚酸与单宁可通过捕获自由基起到抗氧化剂的作用,黄酮类化合物则可以清除自由基和螯合金属[50]。它们共同参与增强机体的氧化还原防御,预防和对抗氧化应激,减少自由基相关的细胞损伤。
酚类成分需要以可溶的形式进入人体血液循环系统,并实现其抗氧化特性。谷物中酚类化合物通常与麸皮中的细胞壁基质相连,以酯化形式出现,难以加工利用。CLINOIU等[51]发现燕麦麸皮经酵母发酵后,其蒽酰胺含量提高了48.5%,阿魏酸含量提高了21.2%。同样对黑麦或麸皮进行酵母发酵,尤其是添加细胞壁降解酶后,游离阿魏酸的含量也得到快速增加[52],这可能与谷物致密的细胞壁基质的降解有关,酶的作用使其结构改变,促进了酚类化合物的释放,而这些酶均来源于发酵微生物。此外这些微生物,如乳酸菌和真菌,可能产生不同的酶,如β-葡萄糖苷酶、酯酶和单宁酶,此类酶均可将酚类聚合物代谢成自由形式或降解产物[47]。SALAR等[53]发现,在玉米发酵过程中,原料中总酚含量增加的同时β-葡萄糖苷酶活性增强,该酶可以水解酚苷,释放游离酚。酚类物质的可提取性增加、新活性化合物的合成以及谷物细胞壁结构破坏导致酚类的释放,共同造成了发酵后酚含量的升高。另外来自发酵微生物的蛋白酶、淀粉酶、木聚糖酶与谷物的作用也同样重要,这些酶促进了谷物改性和化学键的扭曲,从而加速了结合酚类物质的进一步降解。
发酵也可增强酚类组分的抗氧化活性。相较于普通小麦面包,黑麦酸面包的抗氧化性显著提升,并且全麦面包的抗氧化活性最大[54]。杜小燕等[55]利用3株泡盛曲霉对小麦麸皮进行固态发酵,发酵后的抗氧化性得到显著增强,并且活性强弱与酚酸的种类及含量有关。此外,DEY等[56]在探究不同谷物发酵时,发现玉米总酚含量降低与微生物菌株作用于酚类物质和谷物组成的特异性有关,因此微生物活性对单个酚类物质水平的影响可能不同,酚类含量可能还会受到微生物特定菌株的影响。
2.2.3 矿物质
植酸即肌醇六磷酸,是谷物中磷的主要储存形式,也是谷物中常见的抗营养因子,不同谷物中植酸含量在0.18%~6.5%变化且分布各不相同[57]。在玉米中,植酸主要存在于胚乳,而在小麦、黑麦和水稻中,则集中于糊粉组织。植酸的负面营养属性来自于它与铁、钙、镁、锌、锰等金属离子形成不可溶解的螯合物,正是这种螯合特性,植酸被认为是食物中最有效的抗营养物质,也是人类营养中矿物质缺乏的原因[22]。
植酸可被植酸酶去磷酸化,形成游离无机磷酸盐和肌醇磷酸酯,以此增加矿物质的溶解度和生物利用度。但由于人体消化道中缺乏足够水平的植酸酶,因此无法代谢植酸,导致大部分通过粪便排出[57]。谷物原料、乳酸菌和酵母菌均存在不同活性的植酸酶,且易受到发酵有机酸的影响[12]。当pH值降低至5.5,便可通过面粉中内源植酸酶降低约70%的植酸[58]。酵母菌和乳酸菌也具有一定的植酸酶活性,相应的,在商业面包中也早已检测出酵母植酸酶活性[59]。FEKRI等[60]也证明,酸面团微生物群中的酵母菌(Kluyveromycesaestuarii、Kluyveromycesmarxianus和Kluyveromyceslactis)和细菌(Leuconostoccitreum、Enterococcusfaecium和Pediococcuspentosaceus)单菌落表现出高植酸酶活性,植酸降解效果显著,并具有抵抗低pH和胆汁的特性。HARALDSSON等[61]研究显示高植酸酶活性的酵母菌株可能是胃肠道中潜在的植酸酶载体。
植酸酶的酶解作用还受到发酵条件(谷物粒度、温度、酸度、时间和水分含量等)的影响[62]。研究发现,酸面团发酵是溶解全麦面粉中矿物质的有效手段,但对完整麸皮效果较差,只在细磨后的麸皮上检测到了钙和铁的增溶作用。乳酸菌则不受麸皮结构的影响,通过对麸皮进行乳酸预发酵,植酸分解率可接近90%,并且镁和磷的溶解度也随之增加[63]。SPAGGIARI等[64]利用鼠李糖乳杆菌菌株对小麦麸皮进行固态发酵,发酵后的植酸下降到36.4%。微生物发酵法也有助于消除其他抗营养因子,提高矿物质的整体生物利用率,OGODO等[65]利用乳酸菌发酵玉米面粉,发酵后的植酸、单宁、多酚及胰蛋白酶抑制剂含量显著下降。此外在小鼠实验中发现,相较于普通面包,用酸面包喂食的老鼠体内铁的生物利用度更高[66]。这可能是因为发酵中酸对Fe3+的还原作用,促进了小肠上皮细胞游离面的二价金属离子转运体1将铁离子转运至细胞中,而未被还原的Fe3+则不能被吸收。
3 谷物发酵食品的益生功能
3.1 益生菌载体
功能性食品成为当前食品加工的新趋势,生产方法之一是加入益生菌和益生元等成分。益生菌为一类活性微生物,当摄入足够水平时,除了固有的基本营养外,还能为宿主带来其他健康益处[67]。乳酸杆菌属和双歧杆菌属为最常见的益生菌。乳制品原料一直是益生菌的良好载体,但乳糖不耐受,乳蛋白过敏,胆固醇、饱和脂肪含量高等问题使益生菌乳制品受到一定的限制,迫使寻求益生菌非乳制品的开发生产。
近年来,越来越多的谷物基质或谷物发酵产品成为益生菌输送载体的首选之一,因为它们可以满足某些益生菌复杂的营养需求[68],并且在恶劣的胃肠道环境下,可以作为乳酸杆菌的高效转运蛋白,刺激益生微生物的生长[69]。此外,发酵过程中植酸盐的降解,释放出铁、钙、锰(乳酸菌重要的生长因子)等矿物质,这些均为益生菌的存活和繁殖提供了有利的条件。在一些谷物发酵产品中已分离出具有益生特性的菌株,并且显示出较强的生长促进能力[4]。然而由于原料差异、生产条件不规范等因素的影响,使得对特定产品的潜在益生菌菌株的识别和研究过程变得复杂,获得有关益生菌特性的信息也较为有限。XU等[70]对我国发酵大米、自制酸面团和曲中分离的乳酸菌菌株,进行了益生菌特性的体外试验,其中3株植物乳杆菌表现出耐低pH、耐胆盐、抗生素耐药性及抗菌活性,显示出潜在的益生菌特性。据了解,传统饮料Boza发酵所需的微生物,可能随着谷物基质及加工条件的改变而变化,现已从Boza中分离出多株不同类型的益生菌株[71]。最近,研究者发现从伊朗传统酸面团里筛选出具有益生菌特性的乳酸菌和酵母菌,可以被用作生产益生菌全麦面包的发酵剂[60]。并且这些益生菌显示出高效的植酸降解能力,有效解决了全麦产品植酸含量高的缺点。
3.2 益生元潜力
益生元主要是一类低消化率的碳水化合物,它通过选择性地支持结肠中特定微生物属的发育和活动来影响宿主[72],因此具有改善健康的潜力。除碳水化合物外,一些酚类物质、脂肪酸、多肽、微量营养素也被认为具有益生元效应[73]。发酵谷物类产品中可能含有可溶性纤维(β-葡聚糖和AX等)、酚类、抗性淀粉、多肽等活性成分,因此是一种很有前景的益生元化合物。虽然目前关于发酵谷物产品益生元效应的报道还不多,但最近的研究表明,谷物益生元成分对微生物群落,尤其是胃肠道菌群的作用显著[74]。
益生元具有调节肠道菌群的潜力。WANG等[75]在AX的结构与功能研究中发现,AXs可以影响肠道中的微生物群落,并且群落组成可能随着阿拉伯糖基取代位置的改变而变化。TAMURA等[76]研究了人类肠道拟杆菌在谷物多糖β(1,3)/β(1,4)-葡聚糖作用下的生长机制,并强调了其对肠道微生物代谢的重要性。此外,多酚对肠道的调节作用也已被证实,据估计,摄入的多酚只有5%~10%被小肠吸收,其余大部分积聚在大肠中[77],因此易受到大肠腔内微生物酶的影响,将其降解为低分子质量的可吸收活性成分,而该成分反过来又增加了肠道微生物群的生物多样性,并有益于宿主的健康。
除了谷物中的纤维、多酚等益生元外,发酵微生物也可以分泌具有益生元特性的代谢物。目前研究主要集中于特定乳酸菌产生的胞外多糖,如葡聚糖、果聚糖、普寡糖和果寡糖[78]。研究表明这类具有益生元特性的化合物可被肠道微生物代谢[79],并生成乙酸、丁酸盐、丙酸盐等功能成分,丁酸盐可以减少细胞增殖和刺激细胞分化来降低结肠癌的发生风险[80],丙酸盐具有降低血液中的胆固醇和甘油三酯的功效,并且可能具有提高胰岛素敏感性的潜力[81]。
4 总结与展望
发酵可影响谷物淀粉和蛋白质消化率、减少抗营养素及增加酚类化合物和维生素等营养成分,同时,谷物发酵产品作为益生菌载体及益生元对肠道菌群调控具有巨大潜力。目前,大量的体外实验表明谷物发酵产品对人体健康具有积极作用,但仍缺少充分的临床验证。因此为揭示谷物发酵产品对人类健康的根本作用机制,需进一步设计临床实验深入评估可能的因果关系,以确立谷物发酵产品在膳食指南中的地位,并为谷物功能食品的研发及生产提供科学依据。
同时,谷物发酵产品往往会受到产品感官特性的限制,如何把握健康和良好风味之间的平衡,满足世界各地消费者的口味成为一大挑战。因此,需要更好地了解发酵菌株的生理与代谢、发酵产物的风味特征以及风味与消费者接受度之间的关系,并制定策略来满足各地消费者的需求。
猜你喜欢
今日农业(2021年17期)2021-11-26 23:38:44
现代畜牧科技(2021年9期)2021-10-13 06:38:44
今日农业(2021年9期)2021-07-28 07:08:26
军事文摘(2020年20期)2020-11-16 00:32:10
畜牧兽医科技信息(2015年5期)2015-12-27 01:52:03
兽医导刊(2015年8期)2015-03-25 10:36:31
中国洗涤用品工业(2015年7期)2015-02-28 19:02:39
中国酿造(2014年9期)2014-03-11 20:21:04
养殖与饲料(2014年10期)2014-02-28 22:14:51
食品科学(2013年23期)2013-03-11 18:30:09
